Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas


Abstract
1. Introduction
2. Materials and Methods
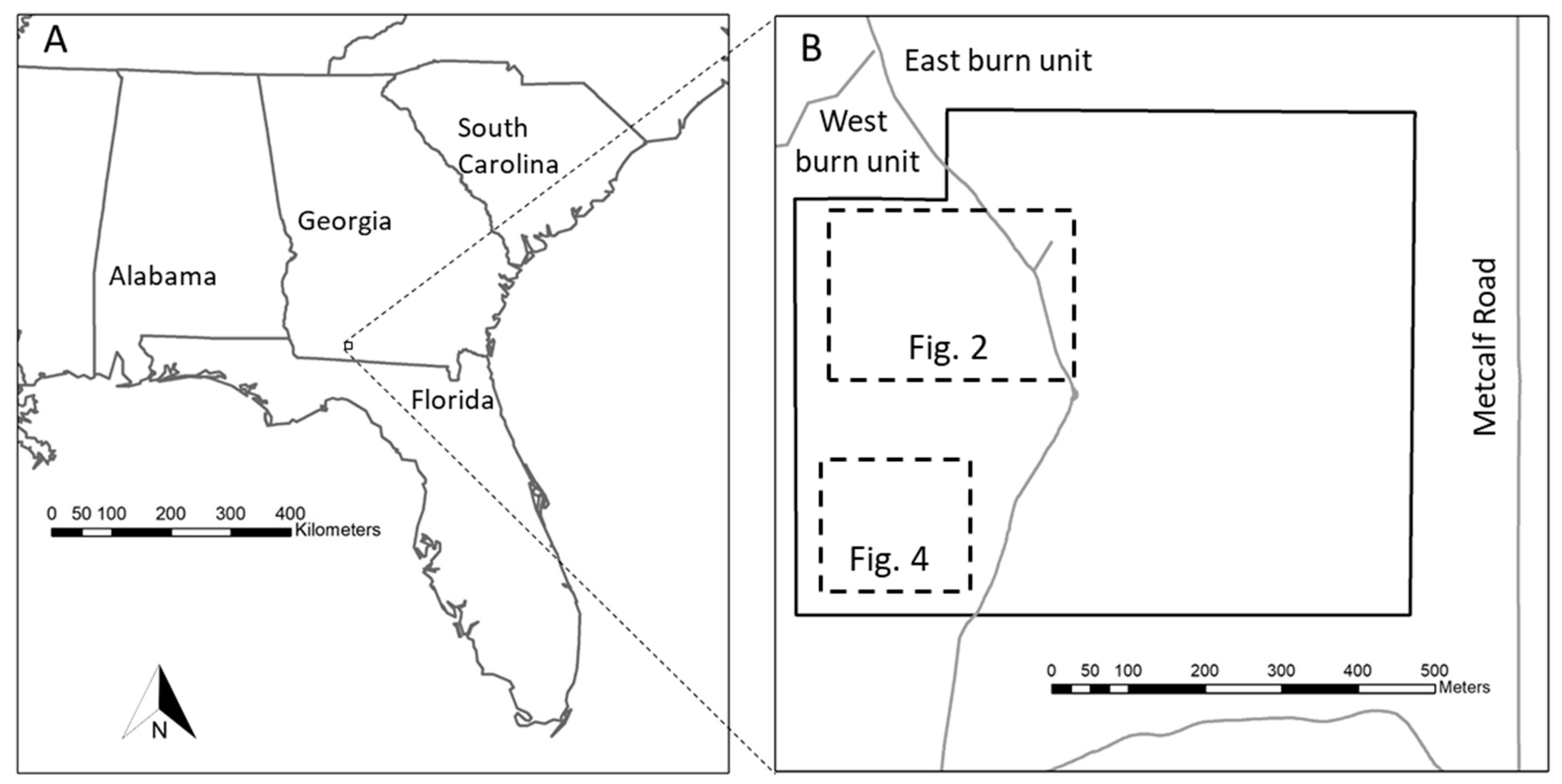
2.1. Study Site
2.2. Data Collection
2.3. Data Analysis
3. Results
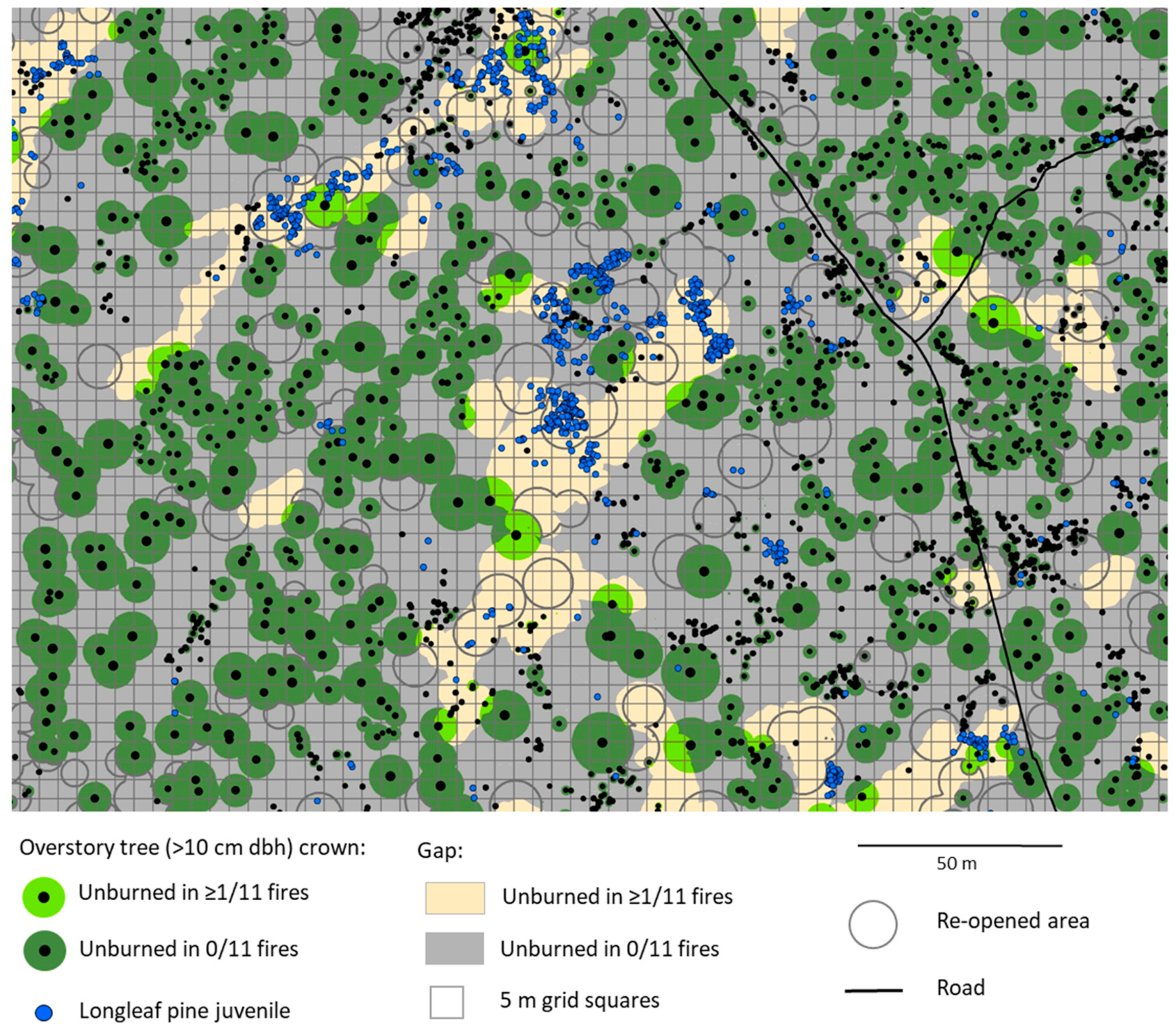
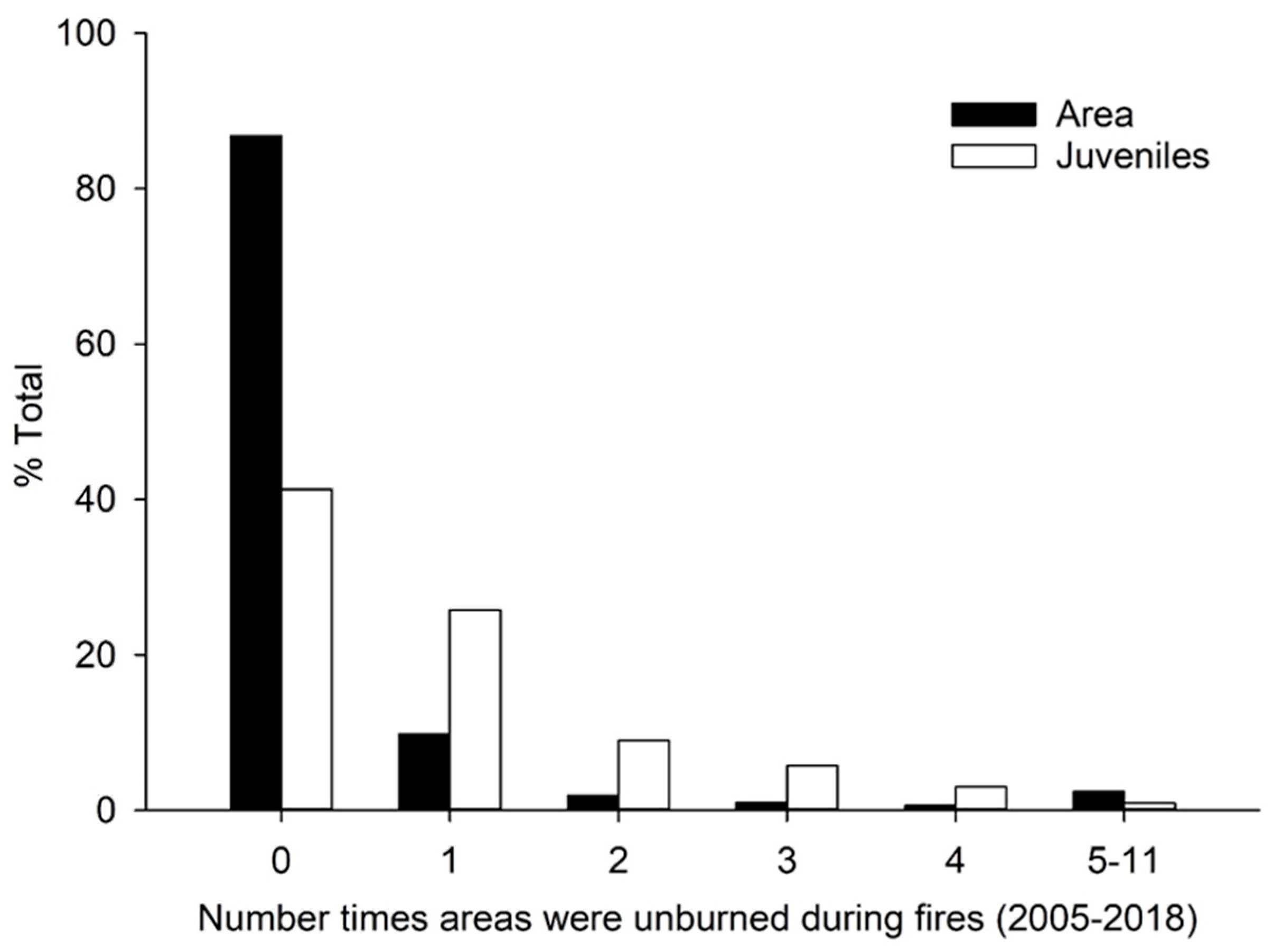
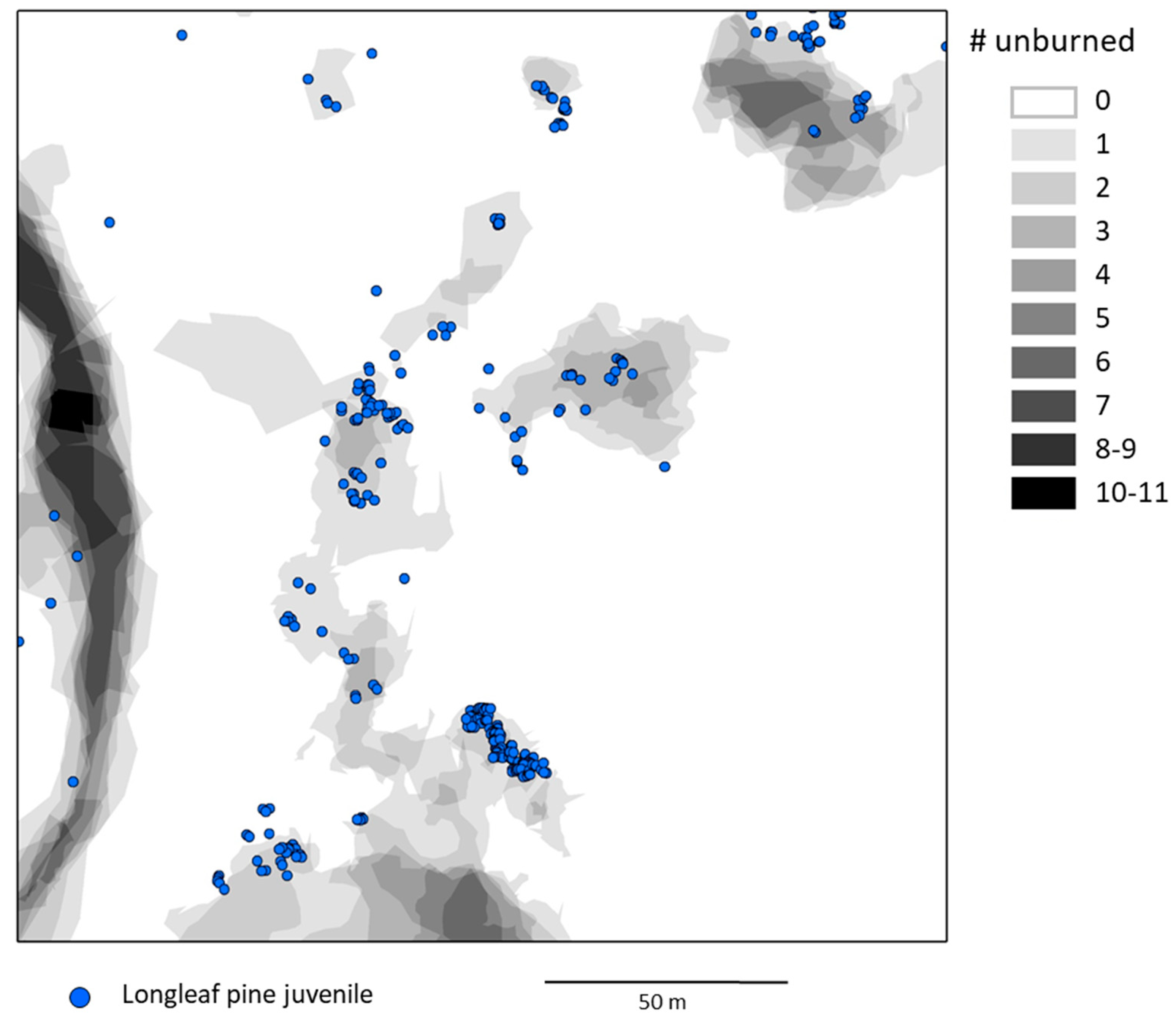
3.1. Spatial Pattern of Unburned Areas
3.2. Longleaf Pine Juveniles
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gap | |||
| Chi-square | 65.75, 1 d.f., p < 0.001 | UB in ≥1 fire % correct | 80.0 |
| Cox & Snell R-sq | 0.064 | UB in 0 fires % correct | 43.7 |
| Nagelkerke’s R-sq | 0.085 | Total % correct | 61.9 |
| ROC AUC | 0.618 | Log likelihood constant model | −692.5 |
| Log likelihood full model | −659.6 | ||
| Inner Gap | |||
| Chi-square | 69.6, 1 d.f., p < 0.001 | UB in ≥1 fire % correct | 90.2 |
| Cox & Snell R-sq | 0.067 | UB in 0 fires % correct | 30.6 |
| Nagelkerke’s R-sq | 0.090 | Total % correct | 60.4 |
| ROC AUC | 0.604 | Log likelihood constant model | −692.5 |
| Log likelihood full model | −657.7 | ||
| Re-Opened Area | |||
| Chi-square | 4.66.7, 1 d.f., p = 0.031 | UB in ≥1 fire % correct | 85.2 |
| Cox & Snell R-sq | 0.005 | UB in 0 fires % correct | 20.0 |
| Nagelkerke’s R-sq | 0.006 | Total % correct | 52.6 |
| ROC AUC | 0.526 | Log likelihood constant model | −692.5 |
| Log likelihood full model | −690.1 | ||
References
- Ratnam, J.; Bond, W.J.; Fensham, R.J.; Hoffmann, W.A.; Archibald, S.; Lehmann, C.E.R.; Anderson, M.T.; Higgins, S.I.; Sankaran, M. Where is a ’forest’ a savanna, and why does it matter. Glob. Ecol. Biogeogr. 2011, 20, 653–660. [Google Scholar] [CrossRef]
- Werner, P.A.; Prior, L.D. Demography and growth of subadult savanna trees: interactions of life history, size, fire season, and grassy understory. Ecol. Monogr. 2013, 83, 67–93. [Google Scholar] [CrossRef]
- Staal, A.; van Nes, E.H.; Hantson, S.; Homgren, M.; Dekker, S.C.; Pueyo, S.; Xu, C.; Scheffer, M. Reslience of tropical tree cover: The roles of climate, fire, and herbivory. Glob. Change Biol. 2018, 24, 5096–5109. [Google Scholar] [CrossRef]
- Wellington, A.B.; Noble, I.R. Seed dynamics and factors limiting recruitment of the mallee Eucalyptus incrassata Labill. in semi-arid, south-eastern Australia. J. Ecol. 1985, 73, 657–666. [Google Scholar] [CrossRef]
- Setterfield, S.A. Seedling establishment in an Australian tropical savanna: effects of seed supply, soil disturbance and fire. J. Appl. Ecol. 2002, 39, 949–959. [Google Scholar] [CrossRef]
- Scholes, R.J.; Archer, S.R. Tree-grass interactions in savannas. Ann. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Van Langevelde, F.; van de Vijver, C.A.D.M.; Kumar, L.; van de Koppel, J.; de Ridder, N.; van Andel, J.; Skidmore, A.K.; Hearne, J.W.; Stroosnijder, L.; Bond, W.J.; et al. Effects of fire and herbivory on the stability of savanna ecosystems. Ecology 2003, 84, 337–350. [Google Scholar] [CrossRef]
- Werner, P.A.; Franklin, D.C. Resprouting and mortality of juvenile eucalypts in an Australian savanna: Impacts of fire season and annual sorghum. Austr. J. Bot. 2010, 58, 619–628. [Google Scholar] [CrossRef]
- Beckage, B.; Gross, L.J.; Platt, W.J. Grass feedbacks on fire stabilize savannas. Ecol. Modell. 2011, 222, 2227–2233. [Google Scholar] [CrossRef]
- Morrison, T.A.; Holdo, R.M.; Rugemalila, D.M.; Nzunda, M.; Anderson, T.M. Grass competition overwhelms effects of herbivores and precipitation on early tree establishment in Serengeti. J. Ecol. 2019, 107, 216–228. [Google Scholar] [CrossRef]
- Higgins, S.I.; Bond, W.J.; Trollope, W.S.W. Fire, resprouting and variability: a recipe for grass-tree coexistence in savanna. J. Ecol. 2000, 88, 213–229. [Google Scholar] [CrossRef]
- Wiegand, K.; Saitz, D.; Ward, D. A patch-dynamics approach to savanna dynamics and woody plant encroachment - insights from an arid savanna. Perspect. Plant Ecol. Evol. Syst. 2006, 7, 229–242. [Google Scholar] [CrossRef]
- Menaut, J.C.; Gignoux, J.; Prado, C.; Clobert, J. Tree community dynamics in a humid savanna of the Cote-d’Ivoire: Modelling the effects of fire and competition with grass and neighbours. J. Biogeogr. 1990, 17, 471–481. [Google Scholar] [CrossRef]
- Jeltsch, F.; Milton, S.J.; Dean, W.R.J.; van Rooyen, N. Tree spacing and coexistence in semiarid savannas. J. Ecol. 1996, 84, 583–595. [Google Scholar] [CrossRef]
- Russell-Smith, J.; Whitehead, P.J.; Cook, G.D.; Hoare, J.L. Response of Eucalyptus-dominated savanna to frequent fires: lessons from Munmarlary, 1973–1996. Ecol. Monogr. 2003, 73, 349–375. [Google Scholar] [CrossRef]
- Beckage, B.; Platt, W.J.; Gross, L.J. Vegetation, fire, and feedbacks: a disturbance-mediated model of savannas. Am. Nat. 2009, 174. [Google Scholar]
- Midgley, J.J.; Lawes, M.J.; Chamaille-Jammes, S. Savanna woody plant dynamics: the role of fire and herbivory, separately and synergistically. Austr. J. Bot. 2010, 58, 1–11. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Enright, N.J.; Knox, K.J.E. Resprouting as a key functional trait: how buds, protection and resource drive persistence after fire. New Phytol. 2012, 197, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Murphy, B.P.; Baker, P.J. The existence of a fire-mediated tree-recruitment bottleneck in an Asian savanna. J. Biogeogr. 2019, 46, 745–756. [Google Scholar] [CrossRef]
- Grubb, P.J. The maintenance of species-richness in plant communities: the importance of the regeneration niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Moustakas, A.; Wiegand, K.; Meyer, K.M.; Ward, D.; Sankaran, M. Learning new tricks from old tress: Revisiting the savanna question. Front. Biogeogr. 2010, 2, 47–53. [Google Scholar]
- Vetaas, O.R. Micro-site effects of trees and shrubs in dry savannas. J. Veg. Sci. 1992, 3, 337–344. [Google Scholar] [CrossRef]
- Hoffmann, W.A. The effects of fire and cover on seedling establishment in a neotropical savanna. J. Ecol. 1996, 84, 383–393. [Google Scholar] [CrossRef]
- McGuire, J.P.; Mitchell, R.J.; Moser, E.B.; Pecot, S.D.; Gjerstad, D.H.; Hedman, C.W. Gaps in a gappy forest: Plant resources, longleaf pine regeneration, and understory response to tree removal in longleaf pine savannas. Can. J. For. Res. 2001, 31, 765–778. [Google Scholar] [CrossRef]
- Jeltsch, F.; Weber, G.E.; Grimm, V. Ecological buffering mechanisms in savannas: A unifying theory of long-term tree-grass coexistence. Plant Ecol. 2000, 161, 161–171. [Google Scholar] [CrossRef]
- Mutch, R.W. Wildland fires and ecosystems--A hypothesis. Ecology 1970, 51, 1046–1051. [Google Scholar] [CrossRef]
- Bond, W.E.; Midgley, J.J. Kill thy neighbor: An individualistic argument for the evolution of flammability. Oikos 1995, 73, 79–85. [Google Scholar] [CrossRef]
- Scarff, F.R.; Westoby, M. Leaf litter flammability in some semi-arid Australian woodlands. Funct. Ecol. 2006, 20, 745–752. [Google Scholar] [CrossRef]
- Platt, W.J.; Evans, G.W.; Rathbun, S.L. The population dynamics of a long-lived conifer (Pinus palustris). Am. Nat. 1988, 131, 491–5225. [Google Scholar] [CrossRef]
- Grace, S.L.; Platt, W.J. Effects of adult tree density and fire on the demography of pregrass stage juvenile longleaf pine (Pinus palustris Mill.). J. Ecol. 1995, 83, 75–86. [Google Scholar] [CrossRef]
- O’Brien, J.J.; Loudermilk, E.L.; Hiers, J.K.; Pokswinski, S.; Hornsby, B.; Hudak, A.; Strother, D.; Rowell, E.; Bright, B.C. Canopy derived fuels drive patterns of in-fire energy release and understory plant mortality in a longleaf pine (Pinus palustris) sandhill in northwest Florida, USA. Can. J. Remote Sens. 2016, 42, 489–500. [Google Scholar] [CrossRef]
- Platt, W.J.; Ellair, D.P.; Huffman, J.M.; Potts, S.E.; Beckage, B. Pyrogenic fuels produced by savanna trees can engineer humid savannas. Ecol. Monogr. 2016, 86, 352–372. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Jaconis, S.Y.; McKinley, K.L.; Geiger, E.L.; Cotsch, S.G.; Franco, A.C. Fuels or microclimate? Understanding the drivers of fire feedbacks at savanna-forest boundaries. Austral Ecol. 2012, 37, 634–643. [Google Scholar] [CrossRef]
- Mugnani, M.P.; Robertson, K.M.; Miller, D.L.; Platt, W.J. Longleaf pine patch dynamics influence ground-layer vegetation in old-growth pine savanna. Forests 2019, in press. [Google Scholar]
- Gilliam, F.S.; Platt, W.J. Effects of long-term fire exclusion on tree species composition and stand structure in an old-growth Pinus palustris (Longleaf pine) forest. Plant Ecol. 1999, 140, 15–26. [Google Scholar] [CrossRef]
- Gignoux, J.; Lahoreau, G.; Julliard, R.; Barot, S. Establishment and early persistence of tree seedlings in an annually burned savanna. J. Ecology 2009, 97, 484–495. [Google Scholar] [CrossRef]
- Bond, W.J.; Parr, C.L. Beyond the forest edge: Ecology, diversity and conservation of the grassy biomes. Biol. Conserv. 2010. [Google Scholar] [CrossRef]
- Brockway, D.G.; Outcalt, K.W. Gap-phase regeneration in longleaf pine wiregrass ecosystems. For. Ecol. Manag. 1998, 106, 125–139. [Google Scholar] [CrossRef]
- Wahlenberg, W.G. Longleaf Pine: Its Use, Ecology, Regeneration, Protection, and Management; Charles Lathrop Pack Forestry Foundation; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 1946; p. 429.
- Prior, L.D.; Williams, R.J.; Bowman, D.M.J.S. Experimental evidence that fire causes a tree recruitment bottleneck in an Australian tropical savanna. J. Trop. Ecol. 2010, 26, 595–603. [Google Scholar] [CrossRef]
- Bond, W.J. What limits trees in C4 grasslands and savannas? Ann. Rev. Ecol. Syst. 2008, 39, 641–659. [Google Scholar] [CrossRef]
- Archibald, S.; Lehmann, C.E.R.; Belcher, C.M.; Bond, W.J.; Bradstock, R.A.; Daniau, A.L.; Dexter, K.G.; Forresetel, E.J.; Greve, M.; He, T.; et al. Biological and geophysical feedbacks with fire in the Earth system. Environ. Res. Lett. 2018, 13, 033003. [Google Scholar] [CrossRef]
- Glitzenstein, J.S.; Streng, D.R.; Wade, D.D. Fire frequency effects on longleaf pine (Pinus palustris P. Miller) vegetation in South Carolina and Northeast Florida, USA. Research Article. Nat. Areas J. 2003, 23, 22–37. [Google Scholar]
- Cox, J.A.; Jones, C.D. Influence of prescribed fire on winter abundance of Bachman’s sparrow. Wilson J. Ornithol. 2009, 121, 359–365. [Google Scholar] [CrossRef]
- Robertson, K.M.; Hmielowski, T.L. Effects of fire frequency and season on resprouting of woody plants in southeastern US pine-grassland communities. Oecologia 2014, 174, 765–776. [Google Scholar] [CrossRef]
- Huffman, J.M. Historical Fire Regimes In Southeastern Pine Savannas. Ph.D. Thesis, Louisiana State University and Agricultural and Mechanical College, Baton Rouge, LA, USA, 2006. [Google Scholar]
- Bale, A.M. Fire effects and litter accumulation dynamics in a montane longleaf pine system. Master’s Thesis, University of Missouri-Columbia, Columbia, MO, USA, May 2009. [Google Scholar]
- Stambaugh, M.C.; Guyette, R.P.; Marschall, J.M. Longleaf pine (Pinus palustris Mill.) fire scars reveal new details of a frequent fire regime. J. veg. Sci. 2011, 22, 1094–1104. [Google Scholar] [CrossRef]
- Freeman, M.E.; Murphy, B.P.; Richards, A.E.; Vesk, P.A.; Cook, G.D. Facultative and obligate trees in a mesic savanna: Fire effects on savanna structure imply contrasting strategies of eco-taxonomic groups. Front. Plant Sci. 2018, 9, 644. [Google Scholar] [CrossRef] [PubMed]
- Rebertus, A.J.; Williamson, G.B.; Moser, E.B. Longleaf pine pyrogenicity and turkey oak mortality in Florida xeric sandhills. Ecology 1989, 70, 60–70. [Google Scholar] [CrossRef]
- Robertson, K.M.; Ostertag, T.E. Effects of land use on fuel characteristics and fire behavior in pinelands of Southwest Georgia. Tall Timbers Fire Ecol. Conf. Proc. 2007, 23, 181–191. [Google Scholar]
- Williamson, G.B.; Black, E.M. High temperature of forest fires under pines as a selective advantage over oaks. Nature 1981, 293, 643–644. [Google Scholar] [CrossRef]
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of fire regime and habitat on tree dynamics in North Florida longleaf pine savannas. Ecol. Monogr. 1995, 65, 141–476. [Google Scholar] [CrossRef]
- Gagnon, J.L.; Jokela, E.J.; Moser, W.K.; Huber, D.A. Characteristics of gaps and natural regeneration in mature longleaf pine flatwoods ecosystems. For. Ecol. Manag. 2004, 187, 373–380. [Google Scholar] [CrossRef]
- Hammond, D.H.; Varner, J.M.; Fan, Z.; Kush, J.S. Long-term stand dynamics of old-growth mountain longleaf pine (Pinus palustris) woodlands. For. Ecol. Manag. 2016, 364, 154–164. [Google Scholar] [CrossRef]
- Noel, J.M.; Platt, W.J.; Moser, E.B. Structural characteristics of old- and second-growth stands of longleaf pine (Pinus palustris) in the Gulf Coastal Region of the U.S.A. Conserv. Biol. 1998, 12, 533–548. [Google Scholar] [CrossRef]
- Platt, W.J.; Rathbun, S.L. Dynamics of an old-growth longleaf pine population. Tall Timbers Fire Ecol. Conf. Proc. 1993, 18, 275–297. [Google Scholar]
- Brockway, D.G.; Outcalt, K.W.; Boyer, W.D. Longleaf pine regeneration ecology and methods. In Springer Series on Environmental Management; Jose, S., Jokela, J., Miller, D.L., Eds.; Springer: New York, NY, USA, 2006; pp. 95–133. [Google Scholar]
- Chen, X.W.; Brockway, D.G.; Guo, Q.F. Characterizing the dynamics of cone production for longleaf pine forests in the southeastern United States. For. Ecol. Manag. 2018, 429, 1–6. [Google Scholar] [CrossRef]
- Reid, A.M.; Robertson, K.M. Energy content of common fuels in upland pine savannas of the south-eastern US and their application to fire behaviour modelling. Int. J. Wildland Fire 2012, 21, 591–595. [Google Scholar] [CrossRef]
- Chapman, H.H. Is the longleaf type a climax? Ecology 1932, 13, 328–334. [Google Scholar] [CrossRef]
- Pessin, L.J. Stimulating the early height growth of longleaf pine seedlings. J. For. 1944, 42, 95–98. [Google Scholar]
- Smith, L.F. Development of longleaf pine seedlings near large trees. J. For. 1955, 53, 289–290. [Google Scholar]
- Palik, B.J.; Mitchell, R.J.; Houseal, G.; Pederson, N. Effects of canopy structure on resource availability and seedling responses in a longleaf pine ecosystem. Can. J. For. Res. 1997, 27, 1458–1464. [Google Scholar] [CrossRef]
- Grace, S.L.; Platt, W.J. Neighborhood effects on juveniles in an old-growth stand of longleaf pine, Pinus palustris. Oikos 1995, 72, 99–105. [Google Scholar] [CrossRef]
- Ellair, D.P.; Platt, W.J. Fuel composition influences fire characteristics and understorey hardwoods in pine savanna. J. Ecol. 2013, 101, 192–201. [Google Scholar] [CrossRef]
- Lawton, D.E.; Marsalis, W.E.; Moye, F.J.; Murray, J.B.; O’Connor, B.J.; Penley, H.M.; Sandrock, G.S.; Friddell, M.S.; Hetrick, J.H.; Huddlestun, P.F.; et al. Geologic Map of Georgia; Georgia Department of Natural Resources and Georgia Geological Survey: Atlanta, GA, USA, 1976.
- Sanders, T.E. Soil survey of Leon County, Florida; U.S. Department of Agriculture Soil Conservation Service; University of Florida: Tallahassee, FL, USA, 1981; p. 151.
- Staff, S.S. Web Soil Survey. Available online: https://websoilsurvey.nrcs.usda.gov/ (accessed on 15 January 2019).
- Gonzalez-Benecke, C.A.; Gezan, S.A.; Samuelson, L.J.; Cropper, W.P.J.; Leduc, D.J.; Martin, T.A. Estimating Pinus palustris tree diameter and stem volume from tree height, crown area and stand-level parameters. J. For. Res. 2014, 25, 43–52. [Google Scholar] [CrossRef]
- Gagnon, J.L.; Jokela, E.J.; Moser, W.K.; Huber, D.A. Dynamics of artificial regeneration in gaps within a longleaf pine flatwoods ecosystem. For. Ecol. Manag. 2003, 172, 133–144. [Google Scholar] [CrossRef]
- O’Brien, J.J.; Hiers, J.K.; Callaham, M.A.J.; Mitchell, R.J.; Jack, S.B. Interactions among overstory structure, seedling life-history traits, and fire in frequently burned neotropical pine forests. Ambio 2008, 37, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Koenig, W.D. Spatial autocorrelation of ecological phenomena. Trends Ecol. Evol. 1999, 14, 22–26. [Google Scholar] [CrossRef]
- Grace, S.L.; Hamrick, J.L.; Platt, W.J. Estimation of seed dispersal in an old-growth population of longleaf pine (Pinus Palustris) using maternity exclusion analysis. Castanea 2004, 69, 207–215. [Google Scholar] [CrossRef]
- Pessin, L.J. Density of stocking and character of ground cover as factors in longleaf pine reproduction. J. For. 1939, 37, 255–258. [Google Scholar]
- Boyer, W.D. Development of Longleaf Pine Seedlings Under Parent Trees. U.S. Forest Research Paper SO-4; Southern Forest Experiment Station, Forest Service, U.S. Department of Agriculture: Asheville, NC, USA, 1963.
- Harrington, T.B.; Dagley, C.M.; Edwards, M.B. Above- and belowground competition from longleaf pine plantations limits performance of reintroduced herbaceous species. For. Sci. 2003, 49, 681–695. [Google Scholar]
- Pessin, L.J.; Chapman, R.A. The effect of living grass on the growth of longleaf pine seedlings in pots. Ecology 1944, 25, 85–90. [Google Scholar] [CrossRef]
- Loudermilk, E.L.; O’Brien, J.J.; Mitchell, R.J.; Cropper, W.P.; Hiers, J.K.; Grunwald, S.; Grego, J.; Fernandez-Diaz, J.C. Linking complex forest fuel structure and fire behaviour at fine scales. Int. J. Wildland Fire 2012, 21, 882–893. [Google Scholar] [CrossRef]
- Dell, J.E.; Richards, L.A.; O’Brien, J.J.; Loudermilk, E.L.; Hudak, A.T.; Pokswinski, S.M.; Bright, B.C.; Hiers, J.K.; Williams, B.W.; Dyer, L.A. Overstory-derived fuels mediate plant species diversity in frequently burned longleaf forests. Ecosphere 2017, 8, e01964. [Google Scholar] [CrossRef]
- Hermann, S.M. Small-scale disturbances in longleaf pine forests. Tall Timbers Fire Ecol. Conf. Proc. 1993, 18, 265–274. [Google Scholar]
- Beckage, B.; Gross, L.J.; Platt, W.J. Modelling responses of pine savannas to climate change and large-scale disturbance. Appl. Veg. Sci. 2006, 9, 75–82. [Google Scholar] [CrossRef]
- Moss, P.T.; Greenwood, D.R.; Archibald, S.B. Regional and local vegetation community dynamics of the Eocene Okanagan Highlands (British Columbia – Washington State) from palynology. Can. J. Earth Sci. 2005, 42, 187–204. [Google Scholar] [CrossRef]
- Boyer, W.D. Impact of Prescribed Fires on Mortality of Released and Unreleased Longleaf Pine Seedlings Under Parent Trees; U.S. Forest Service Southern Forest Experiment Station Research Note SO-182; U.S. Department of Agriculture: New Orleans, LA, USA, 1974; p. 24.
- Ford, C.R.; Minor, E.S.; Fox, G.A. Long-term effects of fire and fire-return interval on population structure and growth of longleaf pine (Pinus palustris). Can. J. For. 2010, 40, 1410–1420. [Google Scholar] [CrossRef]
- Touboul, J.D.; Staver, A.C.; Levin, S.A. On the complex dynamics of savanna landscapes. Proc. Natl. Acad. Sci. USA 2018, 115, E1336–E1345. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Jones, C. Bachman’s Sparrow the Order of the Pheonix. Birding 2008, May/June, 38–45. [Google Scholar]
- Brudvig, L.A.; Grman, E.; Habeck, C.W.; Orrock, J.L.; Ledvina, J.A. Strong legacy of agricultural land use on soils and undestory plant communities in longleaf pine woodlands. For. Ecol. Manag. 2013, 310, 944–955. [Google Scholar] [CrossRef]
- Martin, R.E.; Sapsis, D.B. Fires as agents of biodiversity: Pyrodiversity promotes biodiversity. In Proceedings on the Symposium of Biodiversity in Northwestern California, 1991 pgs 150–151; Kerner, H.M., Ed.; Wildland Resources Center, University of California, Berkeley: Berkeley, CA, USA, 1992; pp. 150–157. [Google Scholar]
- Brockett, B.C.; Biggs, H.C.; van Wilgen, V.W. A patch mosaic burning system for conservation areas in southern African savannas. Int. J. Wildland Fire 2001, 10, 169–183. [Google Scholar] [CrossRef]
- Parr, C.L.; Anderson, A.N. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv. Biol. 2006, 20, 1610–1619. [Google Scholar] [CrossRef] [PubMed]




| Parameter | Estimate | S.E. | Z Score | p | Odds Ratio |
|---|---|---|---|---|---|
| Constant | −0.779 | 0.121 | −6.453 | <0.001 | |
| Gap | 1.132 | 0.144 | 7.881 | <0.001 | 3.103 |
| Constant | −0.260 | 0.071 | −3.638 | <0.001 | |
| Inner gap | 1.399 | 0.179 | 7.812 | <0.001 | 4.049 |
| Constant | −0.061 | 0.070 | −0.870 | 0.384 | |
| Re-opened area | 0.362 | 0.168 | 2.148 | 0.032 | 1.436 |
| UB ≥ 1 Fire | Gap | Inner Gap | Re-Opened | |
|---|---|---|---|---|
| UB ≥ 1 fire | 100 | 81.6 | 28.9 | 17.0 |
| Gap | 26.9 | 100 | 19.7 | 17.5 |
| Inner gap | 48.1 | 100 | 100 | 22.5 |
| Re-opened | 32.2 | 100 | 20.1 | 100 |
| Parameter | Estimate | S.E. | Z Score | p | Odds Ratio | |
|---|---|---|---|---|---|---|
| Constant | −1.562 | 0.319 | −4.892 | <0.001 | ||
| UB area | 0.640 | 0.292 | 2.192 | 0.028 | 1.897 | |
| Gap | 1.270 | 0.332 | 3.825 | <0.001 | 3.561 | |
| Inner gap | 0.718 | 0.357 | 2.012 | 0.044 | 2.051 | |
| Re-opened area | 0.936 | 0.329 | 2.846 | 0.004 | 2.550 | |
| Overall metrics | ||||||
| Chi-square | 46.7, 4 d.f., p <0.0001 | Juveniles present % correct | 63.6 | |||
| Cox & Snell R-sq | 0.162 | Juveniles absent % correct | 70.5 | |||
| Nagelkerke’s R-sq | 0.216 | Total % correct | 67.0 | |||
| Hosmer-Lemeshow | 3.535, p = 0.739, 6 d.f. | Log likelihood constant model | −182.9 | |||
| ROC Area Under Curve | 0.730 | Log likelihood full model | −159.6 | |||
| Cover Category | # UB | ha | % Area | Juveniles | % Total | Density | % in Patches |
|---|---|---|---|---|---|---|---|
| All | 0–11 | 49.4 | 100.0 | 2565 | 100.0 | 51.9 | 80.7 |
| ≥1 | 9.6 | 19.4 | 1553 | 58.7 | 161.8 | 92.5 | |
| 0 | 39.8 | 80.6 | 1012 | 41.3 | 25.4 | 63.1 | |
| Canopy | ≥1 | 2.3 | 4.7 | 143 | 5.6 | 62.2 | 79.7 |
| 0 | 17.3 | 35.1 | 190 | 7.4 | 11.0 | 51.1 | |
| Gap | ≥1 | 7.3 | 14.8 | 1410 | 55.0 | 193.2 | 90.6 |
| 0 | 22.5 | 45.4 | 822 | 32.0 | 36.5 | 69.5 | |
| Inner gap (IG) | ≥1 | 2.5 | 5.1 | 638 | 25.7 | 255.2 | 92.9 |
| 0 | 3.1 | 6.4 | 138 | 5.4 | 44.5 | 71.7 | |
| Re-opened (RO) | ≥1 | 2.1 | 4.1 | 660 | 25.7 | 314.3 | 93.8 |
| 0 | 5.6 | 11.3 | 311 | 12.1 | 55.5 | 82.6 | |
| IG/RO | ≥1 | 0.6 | 1.2 | 268 | 10.4 | 446.7 | 96.6 |
| 0 | 0.8 | 1.6 | 57 | 2.2 | 71.3 | 84.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robertson, K.M.; Platt, W.J.; Faires, C.E. Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas. Forests 2019, 10, 367. https://doi.org/10.3390/f10050367
Robertson KM, Platt WJ, Faires CE. Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas. Forests. 2019; 10(5):367. https://doi.org/10.3390/f10050367
Chicago/Turabian StyleRobertson, Kevin M., William J. Platt, and Charles E. Faires. 2019. "Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas" Forests 10, no. 5: 367. https://doi.org/10.3390/f10050367
APA StyleRobertson, K. M., Platt, W. J., & Faires, C. E. (2019). Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas. Forests, 10(5), 367. https://doi.org/10.3390/f10050367