Effects of Predominant Tree Species Mixing on Lignin and Cellulose Degradation during Leaf Litter Decomposition in the Three Gorges Reservoir, China

 , , ,
, , ,
Abstract
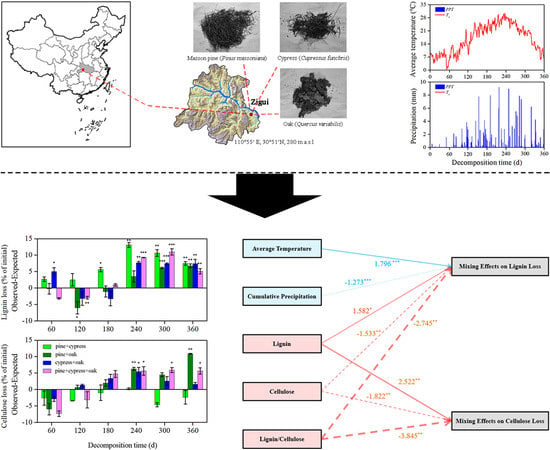
:
1. Introduction
2. Materials and Methods
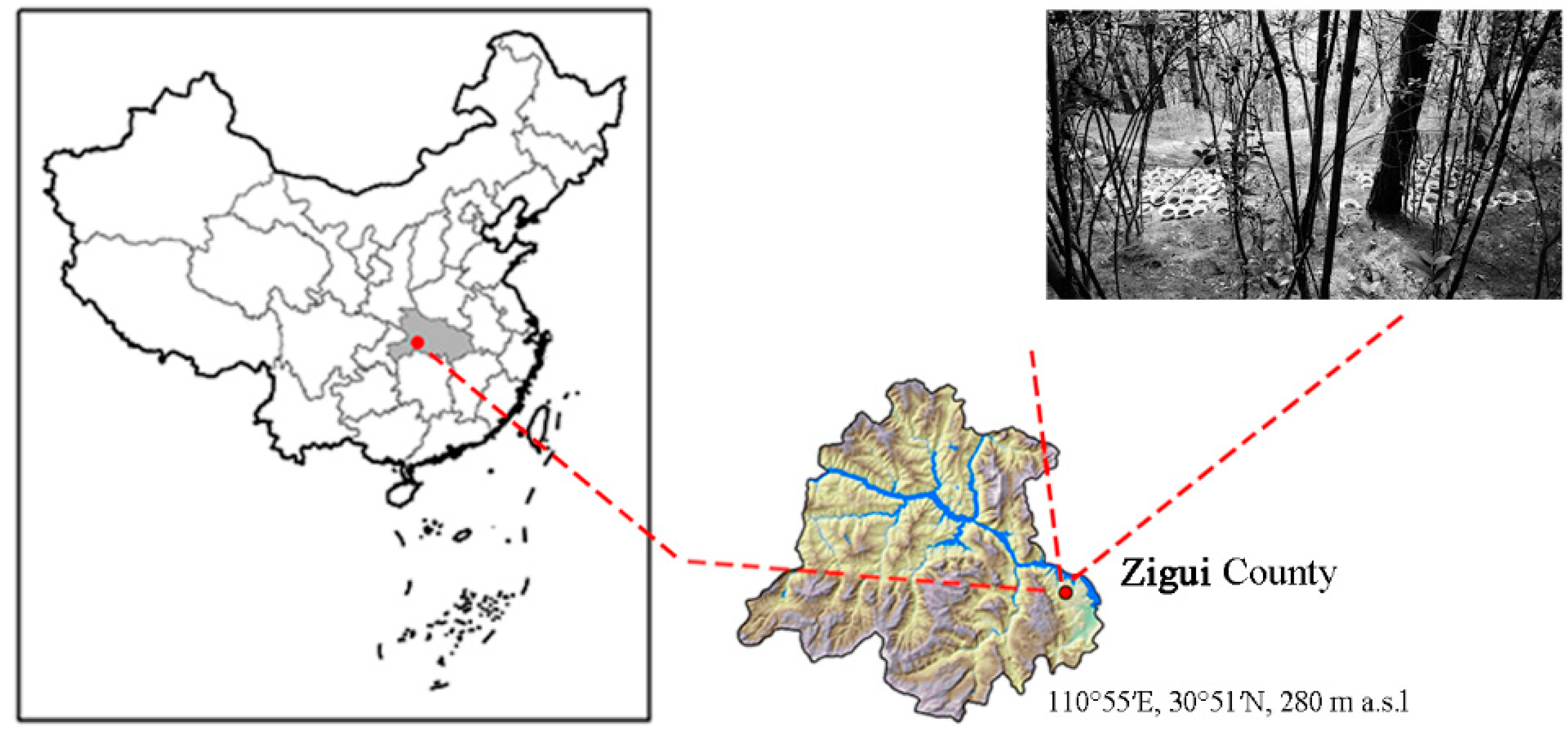
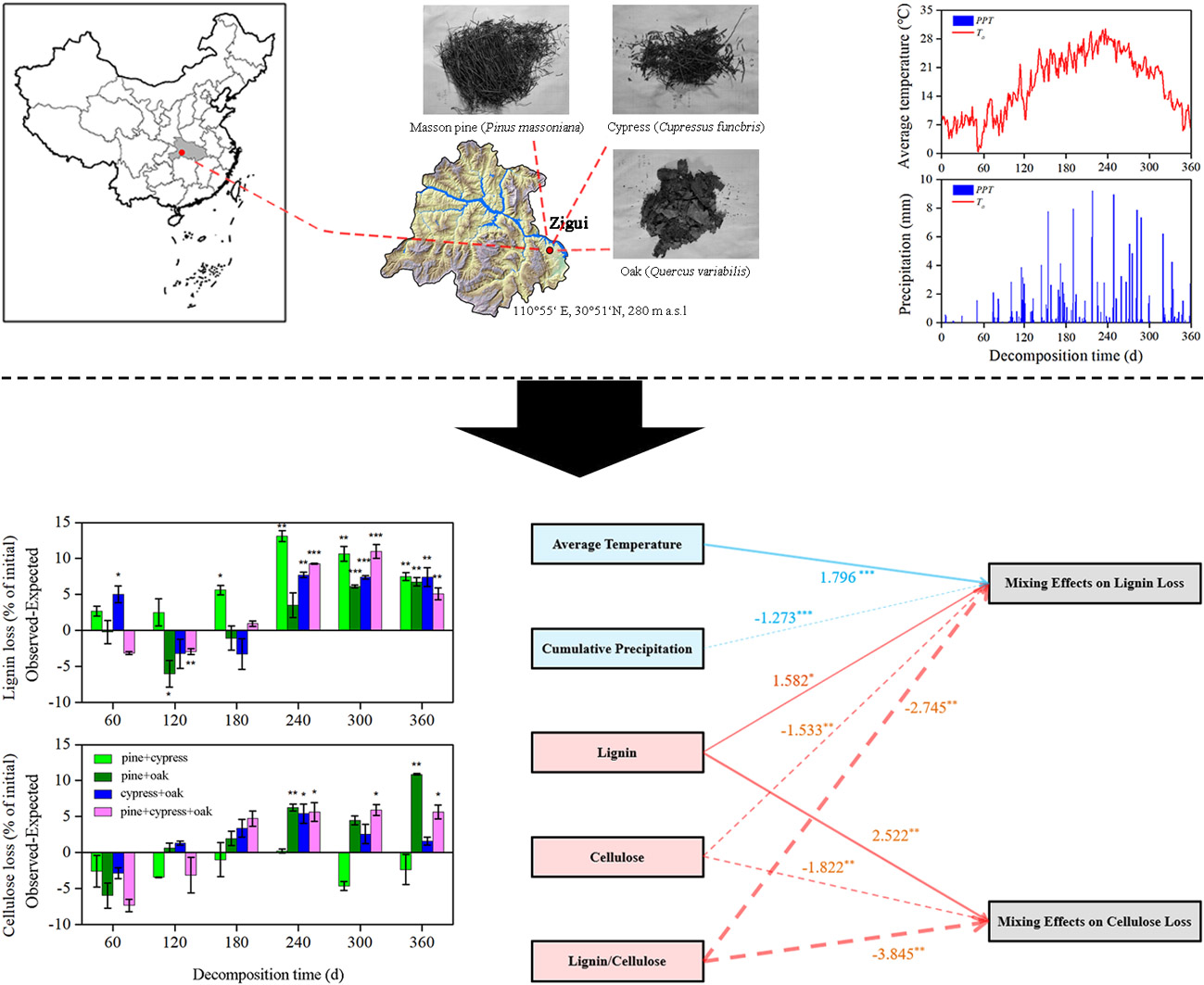
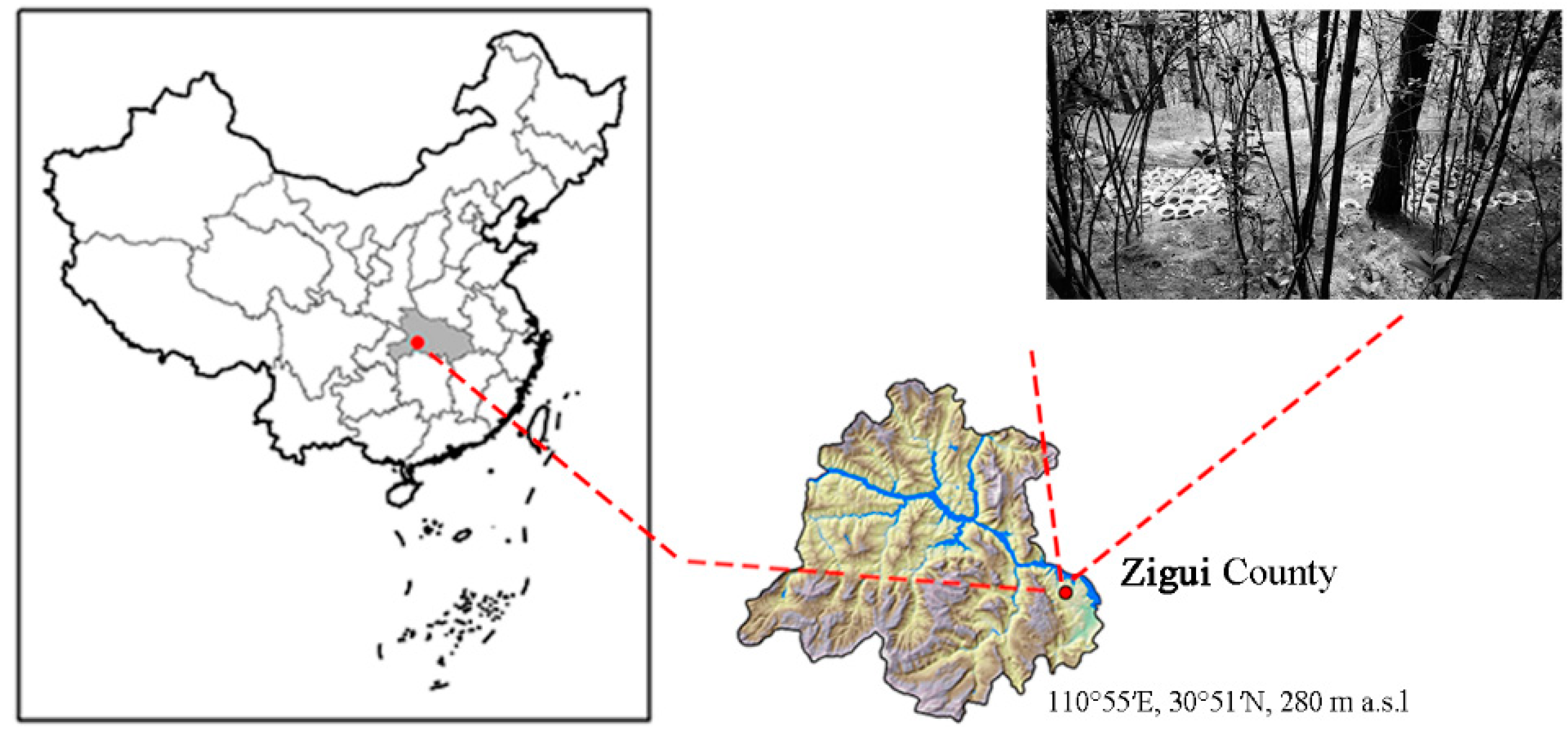
2.1. Study Site and Experimental Design
2.2. Litter Processing
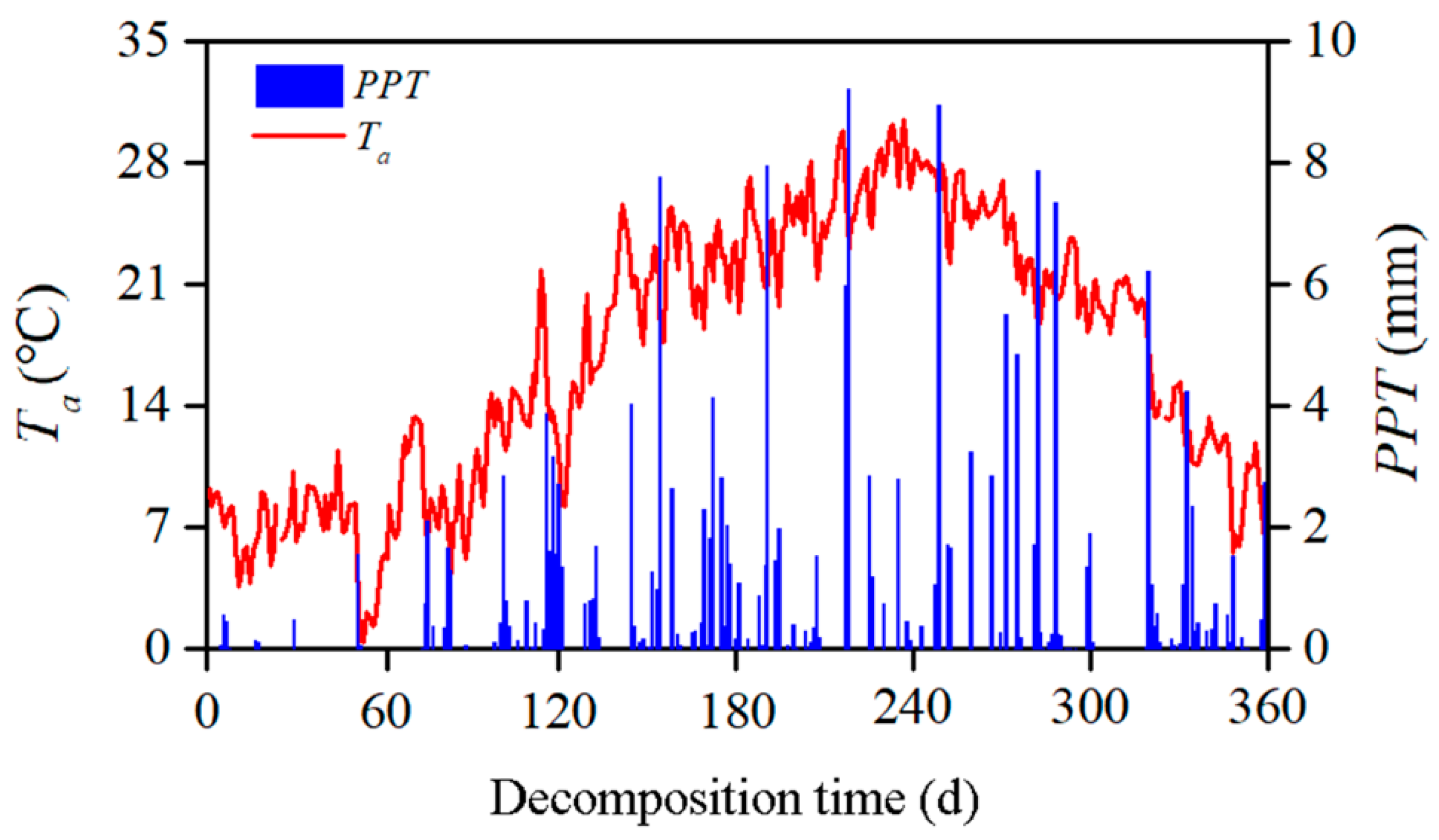
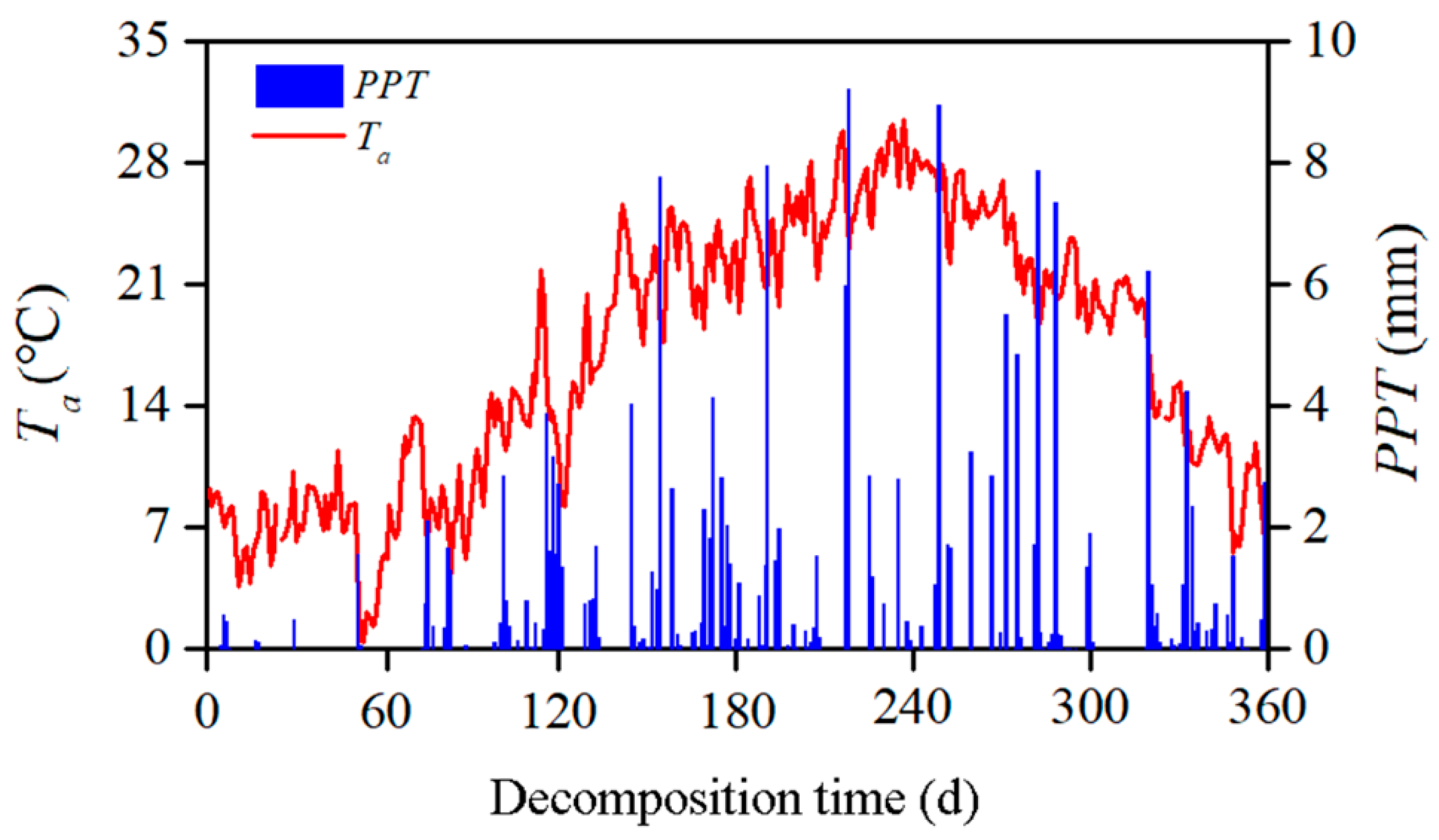
2.3. Sampling and Microenvironmental Measurements
2.4. Analyses and Calculations
2.5. Statistical Analyses
3. Results
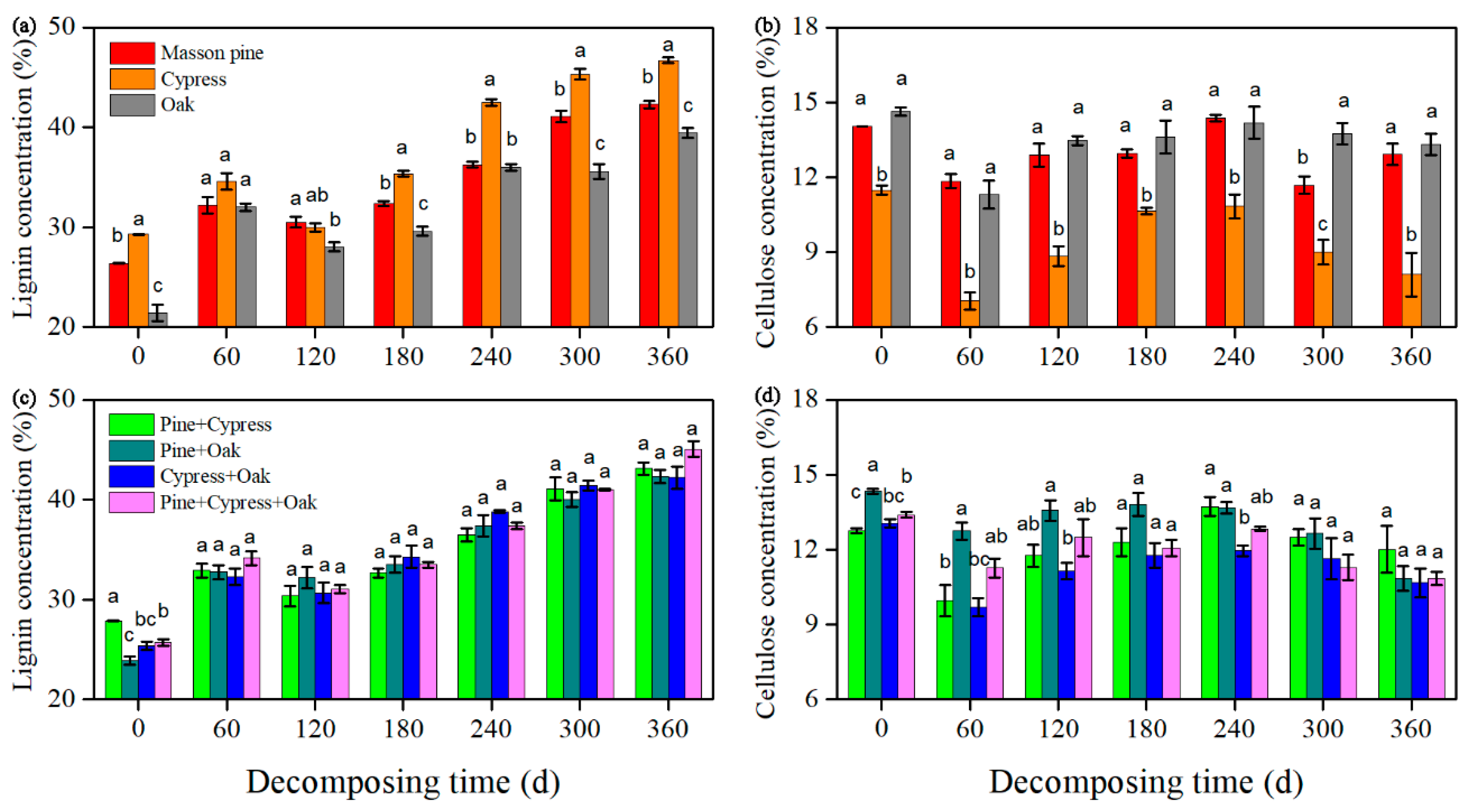
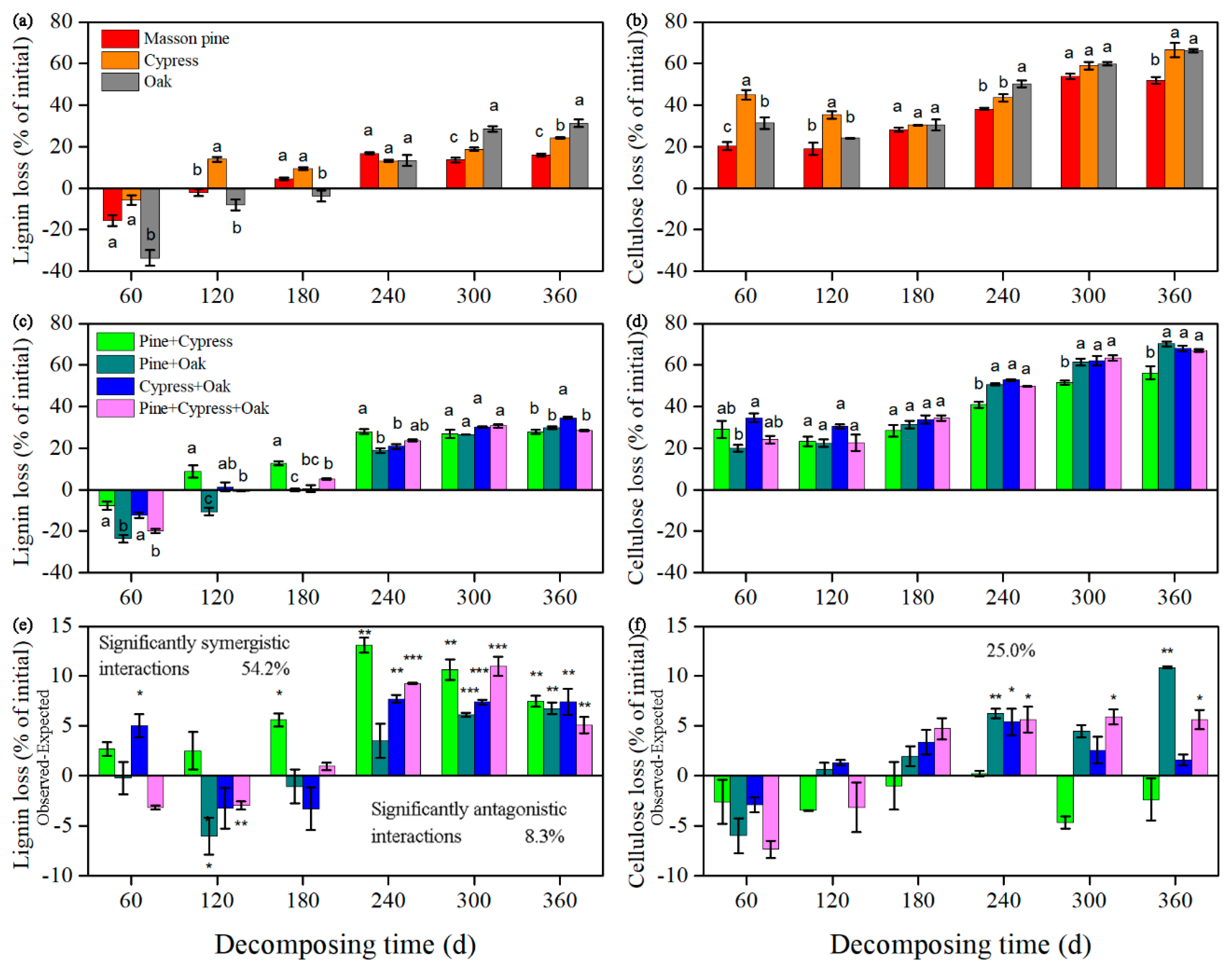
3.1. Effects on the Lignin and Cellulose Concentrations
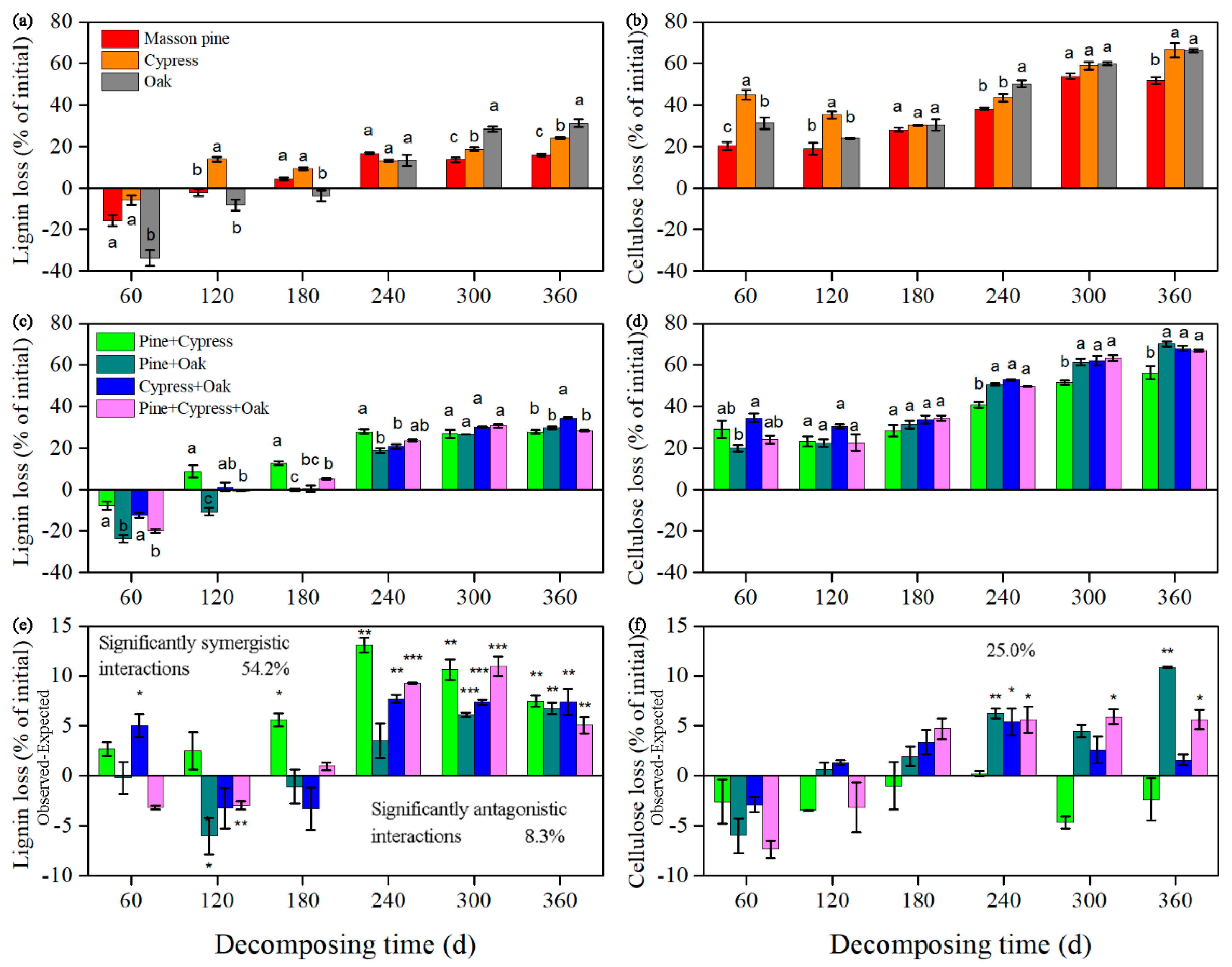
3.2. Effects on Lignin and Cellulose Loss
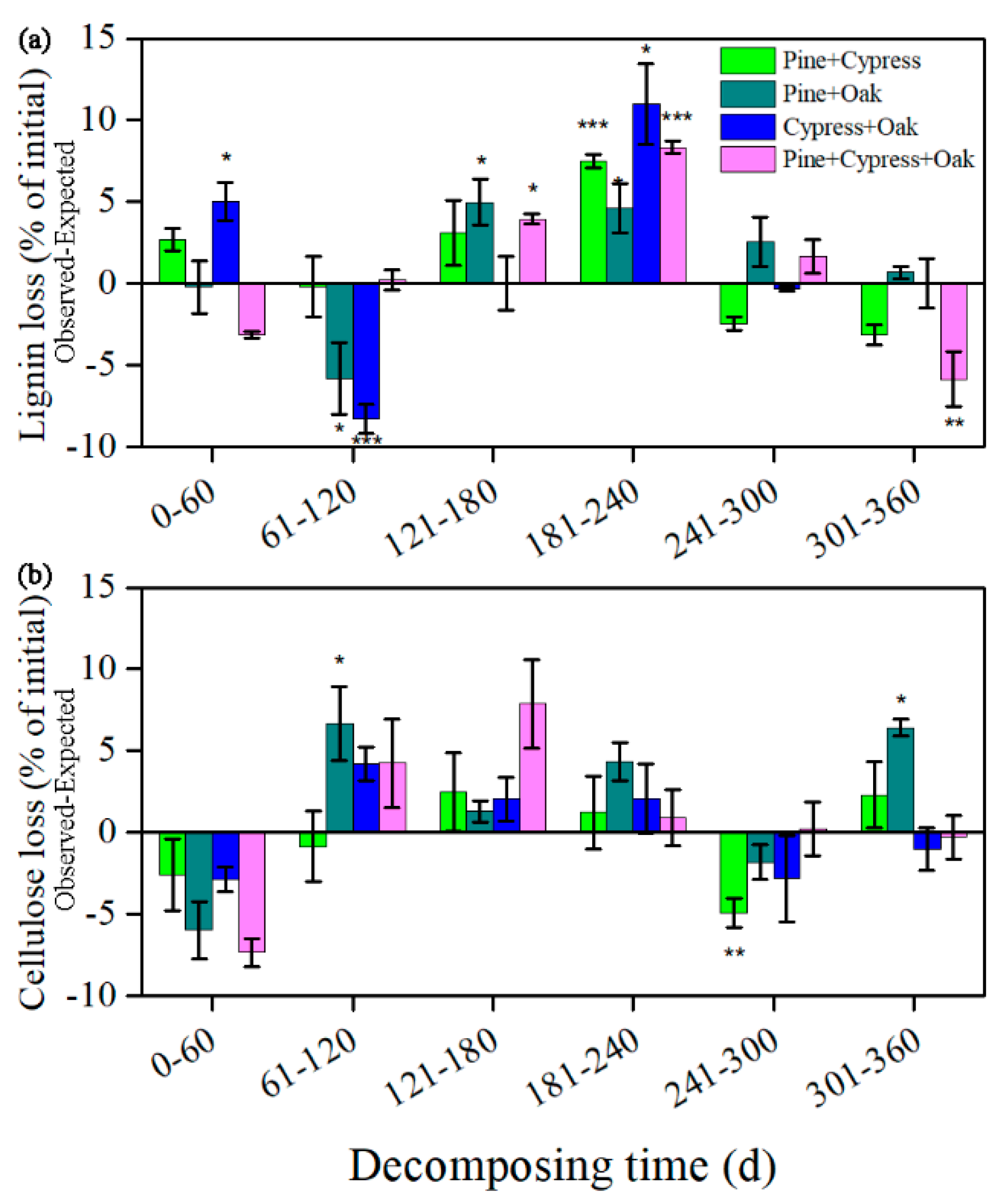
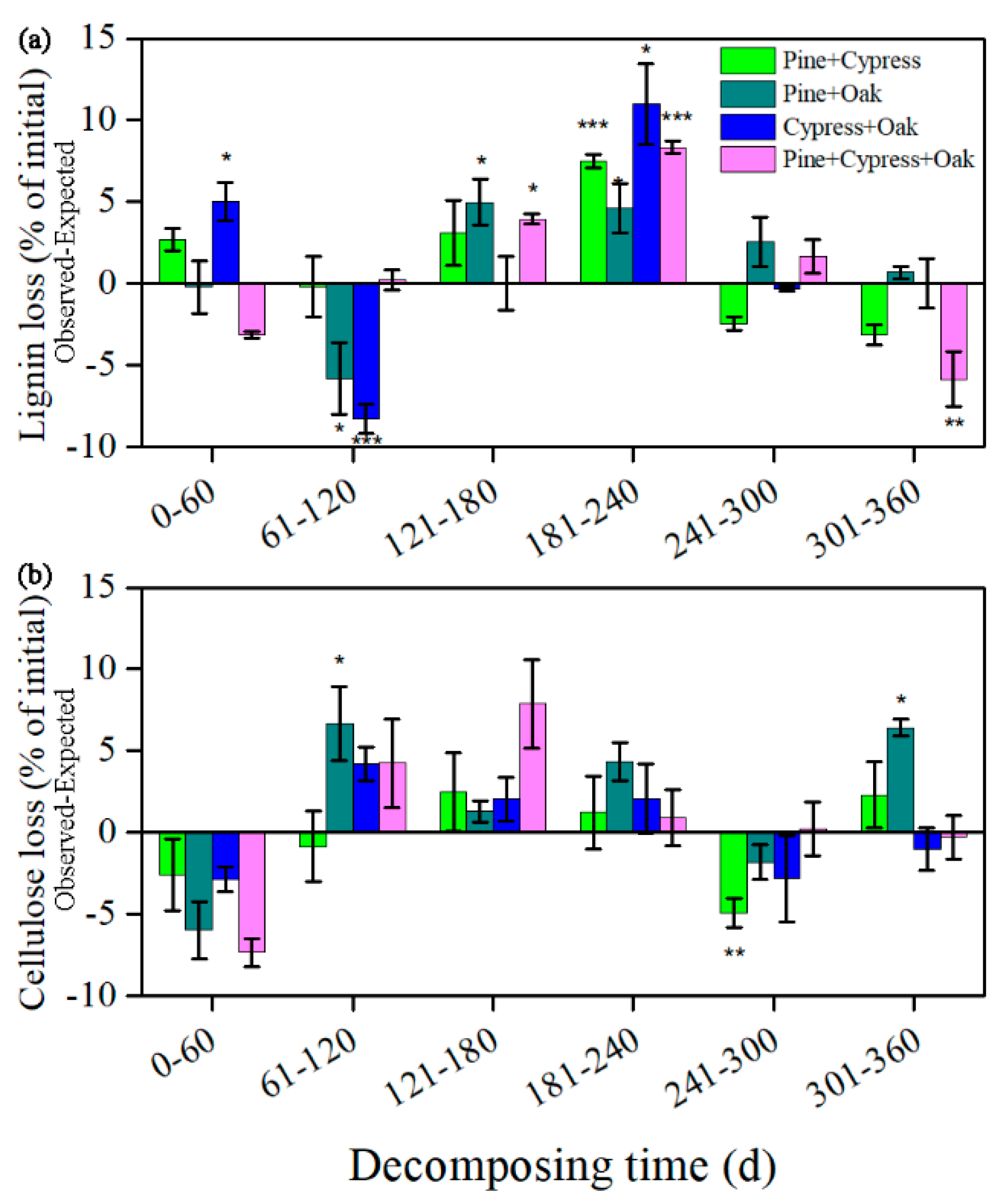
3.3. Synergism and Antagonism in the Litter Mixtures
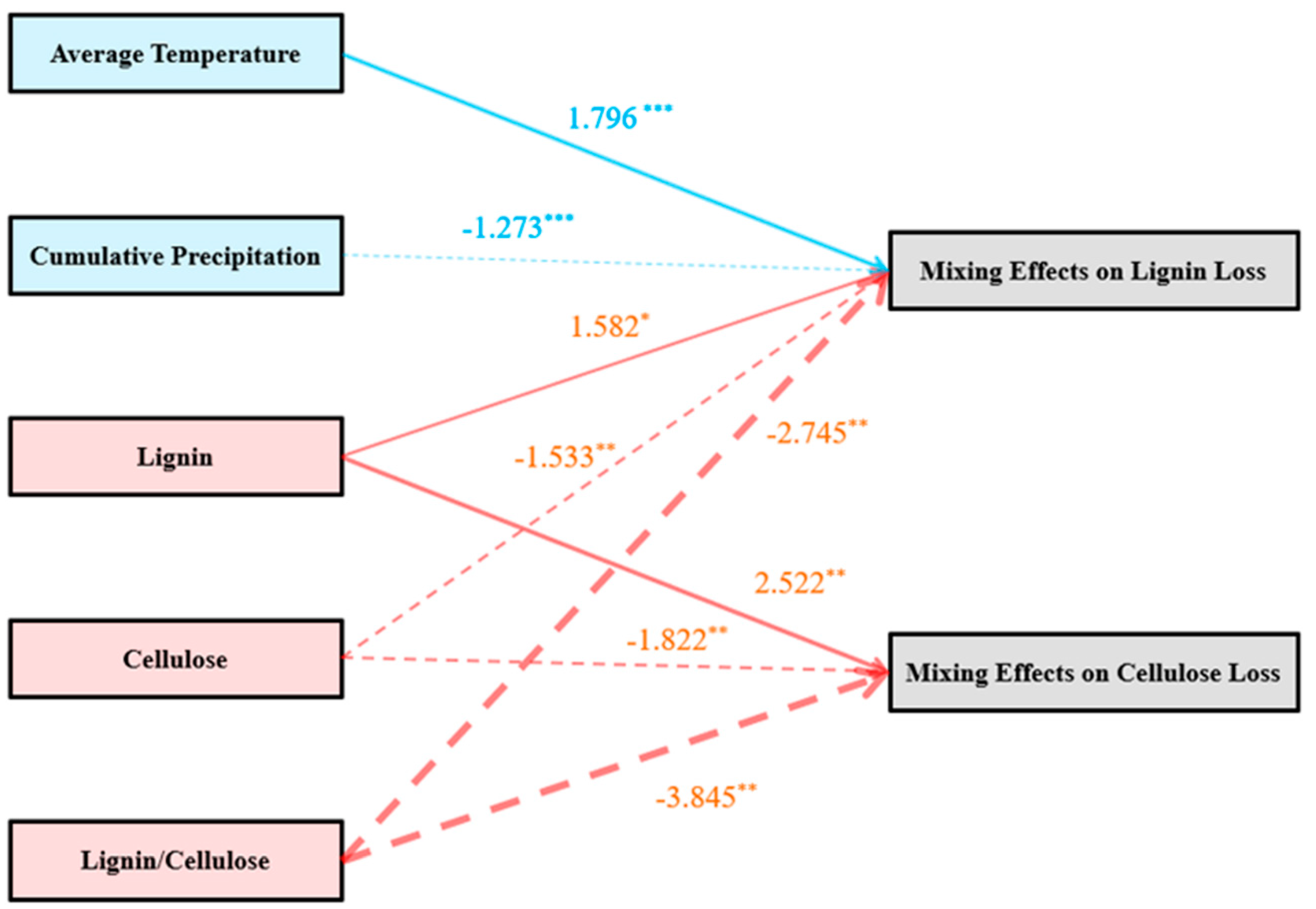
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Zhu, J.X.; Hu, H.F.; Tao, S.L.; Chi, X.L.; Li, P.; Jiang, L.L.; Ji, C.J.; Zhu, J.L.; Tang, Z.Y.; Pan, Y.D.; et al. Carbon stocks and changes of dead organic matter in China’s forests. Nat. Commun. 2017, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- Lemma, B.; Nilsson, I.; Kleja, D.B.; Olsson, M.; Knicker, H. Decomposition and substrate quality of leaf litters and fine roots from three exotic plantations and a native forest in the southwestern highlands of Ethiopia. Soil Biol. Biochem. 2007, 39, 2317–2328. [Google Scholar] [CrossRef]
- He, W.; Wu, F.Z.; Zhang, D.J.; Yang, W.Q.; Bo, T.; Zhao, Y.Y.; Wu, Q.Q. The effects of forest gaps on cellulose degradation in the foliar litter of two shrub species in an alpine fir forest. Plant Soil 2015, 393, 109–122. [Google Scholar] [CrossRef]
- He, W.; Wu, F.Z.; Yang, W.Q.; Tan, B.; Zhao, Y.Y.; Wu, Q.Q.; He, M. Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest. Ecosystems 2016, 19, 115–128. [Google Scholar]
- Cox, P.; Wilkinson, S.P.; Anderson, J.M. Effects of fungal inocula on the decomposition of lignin and structural polysaccharides in Pinus sylvestris litter. Biol. Fert. Soils. 2001, 33, 246–251. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Wang, L.F.; Zhang, J.; He, R.L. Impacts of soil fauna on lignin and cellulose degradation in litter decomposition across an alpine forest-tundra ecotone. Eur. J. Soil. Biol. 2018, 87, 53–60. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant litter. Decomposition, Humus Formation, Carbon Sequestration, 3rd ed.; Springer: Berlin, Germany, 2014. [Google Scholar]
- Yue, K.; Wu, F.Z.; Yang, W.Q.; Zhang, C.; Peng, Y.; Tan, B.; Xu, Z.F.; Huang, C.P. Cellulose dynamics during foliar litter decomposition in an alpine forest meta-ecosystem. Forests 2016, 7, 176. [Google Scholar] [CrossRef]
- Austin, A.T.; Ballare´, C.L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2010, 107, 4618–4622. [Google Scholar] [CrossRef]
- Wu, D.D.; Li, T.T.; Wan, S.Q. Time and litter species composition affect litter-mixing effects on decomposition rates. Plant Soil 2013, 371, 355–366. [Google Scholar] [CrossRef]
- Swan, C.M.; Healey, B.; Richardson, D.C. The role of native riparian tree species in decomposition of invasive tree of heaven (Ailanthus altissima) leaf litter in an urban stream. Ecoscience 2008, 15, 27–35. [Google Scholar] [CrossRef]
- Finziab, A.C.; Canhamb, C.D. Non-additive effects of litter mixtures on net N mineralization in a southern New England forest. For. Ecol. Manag. 1998, 105, 129–136. [Google Scholar] [CrossRef]
- Liu, C.C.; Liu, Y.G.; Guo, K.; Zhao, H.W.; Qiao, X.G.; Wang, S.J.; Zhang, L.; Cai, X.L. Mixing litter from deciduous and evergreen trees enhances decomposition in a subtropical karst forest in southwestern China. Soil Biol. Biochem. 2016, 101, 44–54. [Google Scholar] [CrossRef]
- Meier, C.L.; Bowman, W.D. Links between plant litter chemistry, species diversity, and below-ground ecosystem function. Proc. Natl. Acad. Sci. USA 2008, 105, 19780–19785. [Google Scholar] [CrossRef] [PubMed]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-species plantations of eucalyptus with nitrogen fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef]
- Forrester, D.I.; Pares, A.; O’Hara, C.; Khanna, P.K.; Bauhus, J. Soil organic carbon is increased in mixed-species plantations of eucalyptus and nitrogen-fixing acacia. Ecosystems 2013, 16, 123–132. [Google Scholar] [CrossRef]
- De Marco, A.; Meola, A.; Maisto, G.; Giordano, M.; De Santo, A.V. Non-additive effects of litter mixtures on decomposition of leaf litters in a Mediterranean maquis. Plant Soil 2011, 344, 305–317. [Google Scholar] [CrossRef]
- Harrison, A.F. The inhibitory effect of oak leaf litter tannins on the growth of fungi, in relation to litter decomposition. Soil Biol. Biochem. 1971, 3, 167–172. [Google Scholar] [CrossRef]
- Vivanco, L.; Austin, A.T. Tree species identity alters forest litter decomposition through long-term plant and soil interactions in Patagonia, Argentina. J. Ecol. 2008, 96, 727–736. [Google Scholar] [CrossRef]
- Department of Forest Resources Management, State Forestry Administration. The 7th national forest inventory and status of forest resources. For. Resour. Manag. 2010, 1, 1–8. (In Chinese) [Google Scholar]
- Ge, X.G.; Zeng, L.X.; Xiao, W.F.; Huang, Z.L.; Zhou, B.Z. Dynamic of leaf litter stoichiometric traits dynamic and its relations with decomposition rates under three forest types in Three Gorges Reservoir Area. Acta Ecol. Sinica 2015, 35, 779–787, (In Chinese, English abstract). [Google Scholar]
- Sun, X.; Kang, H.Z.; Du, H.M.; Hu, H.B.; Zhou, J.B.; Hou, J.l.; Zhou, X.; Liu, C.J. Stoichiometric traits of oriental oak (Quercus variabilis) acorns and their variations in relation to environmental variables across temperate to subtropical China. Ecol. Res. 2012, 27, 765–773. [Google Scholar] [CrossRef]
- Zeng, L.X.; He, W.; Teng, M.J.; Luo, X.; Yan, Z.G.; Huang, Z.L.; Zhou, Z.X.; Wang, P.C.; Xiao, W.F. Effects of mixed leaf litter from predominant afforestation tree species on decomposition rates in the Three Gorges Reservoir, China. Sci. Total Environ. 2018, 639, 679–686. [Google Scholar] [CrossRef]
- Ayres, E.; Steltzer, H.; Simmons, B.L.; Simpson, R.T.; Steinweg, J.M.; Wallenstein, M.D.; Mellor, N.; Parton, W.J.; Moore, J.C.; Wall, D.H. Home-field advantage accelerates leaf litter decomposition in forests. Soil Biol. Biochem. 2009, 41, 606–610. [Google Scholar] [CrossRef]
- Chomel, M.; Guittonny-Larchevêque, M.; Desrochers, A.; Baldy, V. Home field advantage of litter decomposition in pure and mixed plantations under boreal climate. Ecosystems 2015, 18, 1014–1028. [Google Scholar] [CrossRef]
- Vanderbilt, K.L.; White, C.S.; Hopkins, O.; Craig, J.A. Aboveground decomposition in arid environments: Results of a longterm study in central New Mexico. J. Arid. Environ. 2008, 72, 696–709. [Google Scholar] [CrossRef]
- Gartner, T.B.; Cardon, Z.G. Decomposition dynamics in mixed-species leaf litter. Oikos 2004, 104, 230–246. [Google Scholar] [CrossRef]
- Mao, B.; Zeng, D.H. Non-additive effects vary with the number of component residues and their mixing proportions during residue mixture decomposition: A microcosm study. Geoderma 2012, 170, 112–117. [Google Scholar] [CrossRef]
- Mu, J.P.; Yang, Y.L.; Luo, Y.L. Pollinator preference and pollen viability mediated by flower color synergistically determine seed set in an Alpine annual herb. Ecol. Evol. 2017, 7, 2947–2955. [Google Scholar] [CrossRef]
- Taylor, B.R.; Parkinson, D.; Parsons, W.F. Nitrogen and lignin content as predictors of litter decay rates: A microcosm test. Ecology 1989, 70, 97–104. [Google Scholar] [CrossRef]
- McClaugherty, C.; Berg, B. Cellulose, lignin and nitrogen concentrations as rate regulating factors in late stages of forest litter decomposition. Pedobiologia 1987, 30, 101–112. [Google Scholar]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The microbial efficiency-matrix stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Aerts, R.; De Caluwe, H. Nutritional and plant-mediated controls on leaf litter decomposition of Carex species. Ecology 1997, 78, 244–260. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A.; Berg, B.; Maynard, D.S.; Wieder, W.R.; Wood, S.A. Understanding the dominant controls on litter decomposition. J. Ecol. 2016, 104, 229–238. [Google Scholar] [CrossRef]
- Wang, Y.J.; Müller-Schärer, H.; van Kleunen, M.; Cai, A.M.; Zhang, P.; Yan, R.; Dong, B.C.; Yu, F.H. Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.Z.; Peng, C.H.; Yang, W.Q.; Zhang, J.; Han, Y.; Mao, T. Admixture of alder (Alnus formosana) litter can improve the decomposition of eucalyptus (Eucalyptus grandis) litter. Soil Biol. Biochem. 2014, 73, 115–121. [Google Scholar] [CrossRef]
- Lecerf, A.; Risnoveanu, G.; Popescu, C.; Gessner, M.O.; Chauvet, E. Decomposition of diverse litter mixtures in streams. Ecology 2007, 88, 219–227. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Tiunov, A.V.; Scheu, S. Biodiversity and litter decomposition in terrestrial ecosystems. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Keith, A.M.; Van Der Wal, R.; Brooker, R.W.; Osler, G.H.R.; Chapman, S.J.; Burslem, D.F.R.P.; Elston, D.A. Increasing litter species richness reduces variability in a terrestrial decomposer system. Ecology 2008, 89, 2657–2664. [Google Scholar] [CrossRef]
- Gessner, M.O.; Swan, C.M.; Dang, C.K.; McKie, B.G.; Bardgett, R.D.; Wall, D.H.; Hättenschwiler, S. Diversity meets decomposition. Trends Ecol. Evol. 2010, 25, 372–380. [Google Scholar] [CrossRef]
- Norby, R.J.; Kozlowski, T.T. Allelopathic potential of ground cover species on Pinus resinosa seedlings. Plant Soil 1980, 57, 363–374. [Google Scholar] [CrossRef]
- Kil, B.S.; Yim, Y.J. Allelopathic effects of Pinus densiflora on undergrowth of red pine forest. Plant Soil 1983, 9, 1135–1151. [Google Scholar]
- Barantal, S.; Roy, J.; Fromin, N.; Schimann, H.; Hättenschwiler, S. Long-termpresence of tree species but not chemical diversity affect litter mixture effects on decomposition in a neotropical rainforest. Oecologia 2011, 167, 241–252. [Google Scholar] [CrossRef]
- Lecerf, A.; Marie, G.; Kominoski, J.S.; LeRoy, C.J.; Bernadet, C.; Swan, C.M. Incubation time, functional litter diversity, and habitat characteristics predict litter-mixing effects on decomposition. Ecology 2011, 92, 160–169. [Google Scholar] [CrossRef]
- Coûteaux, M.M.; Bottner, P.; Berg, B. Litter decomposition, climate and litter quality. Trends Ecol. Evol. 1995, 10, 63–66. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
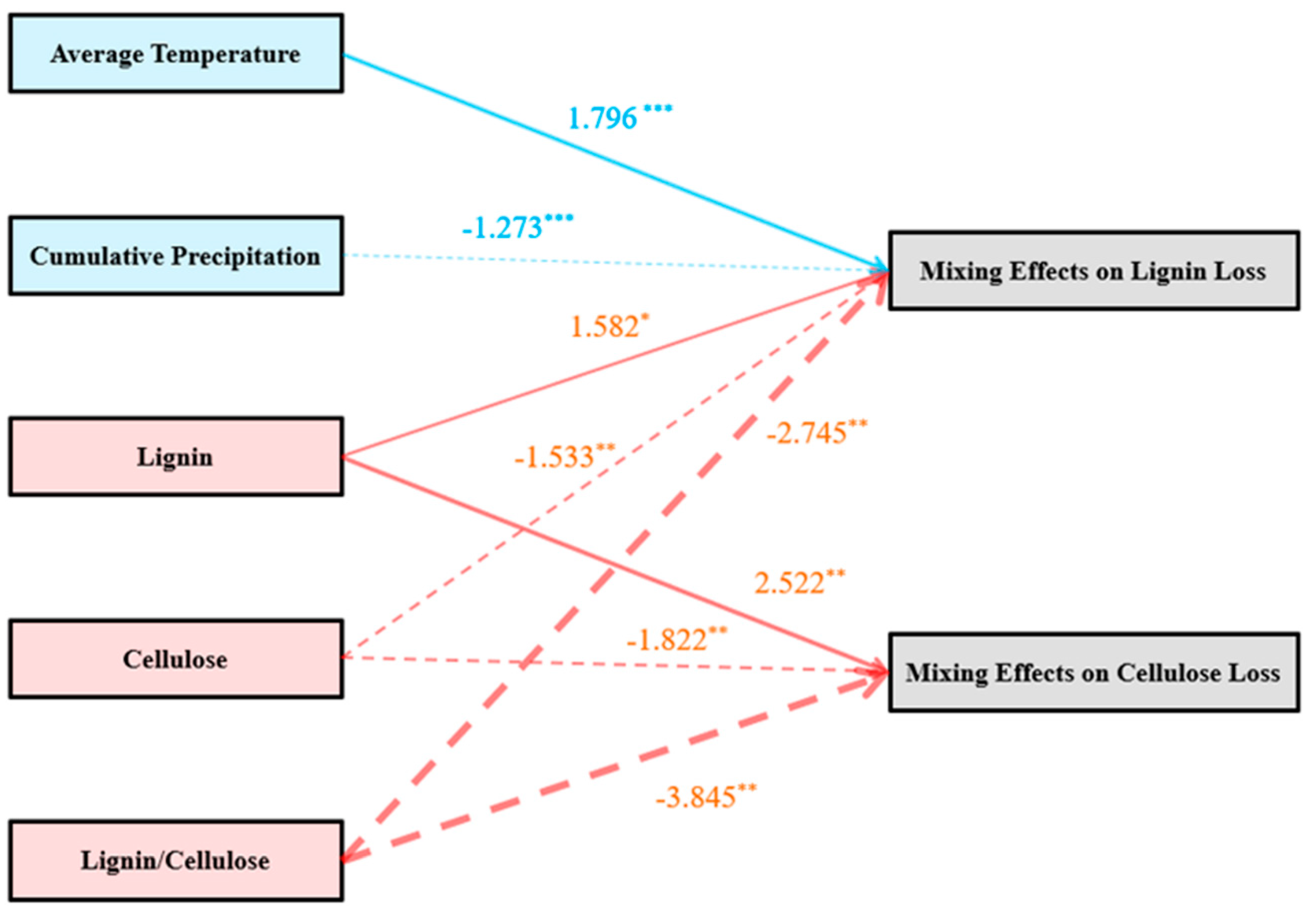{kind=link}
| Index | Lignin | Cellulose | Lignin/Cellulose | Average Temperature | Cumulative Precipitation | |
|---|---|---|---|---|---|---|
| (A) | Lignin loss | (+) 11.733 ** (0.184) | (−) 6.570 * (0.112) | (+) 9.773 ** (0.158) | (+) 8.106 ** (0.135) | (+) 12.794 ** (0.197) |
| Cellulose loss | (−) 6.984 * (0.118) | (+) 13.978 *** (0.212) | (−) 14.850 *** (0.222) | 0.931 (0.018) | (−) 4.944 * (0.087) | |
| (B) | Lignin loss | (+) 16.690 *** (0.193) | (−) 15.983 *** (0.186) | (+) 26.476 *** (0.274) | (+) 31.202 *** (0. 083) | (+) 37.041 *** (0.346) |
| Cellulose loss | (−) 21.477 *** (0.235) | (+) 45.747 *** (0.395) | (−) 66.224 *** (0.486) | 0.520 (0.007) | (−) 5.325 * (0.071) |
| Best Model | AIC |
|---|---|
| Nonadditive lignin loss = Lignin − Cellulose * − Lignin/Cellulose * + Average Temperature *** + Cumulative Precipitation *** | 186.77 |
| Nonadditive cellulose loss = Lignin − Cellulose * − Lignin/Cellulose * | 272.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, W.; Ma, Z.; Pei, J.; Teng, M.; Zeng, L.; Yan, Z.; Huang, Z.; Zhou, Z.; Wang, P.; Luo, X.; et al. Effects of Predominant Tree Species Mixing on Lignin and Cellulose Degradation during Leaf Litter Decomposition in the Three Gorges Reservoir, China. Forests 2019, 10, 360. https://doi.org/10.3390/f10040360
He W, Ma Z, Pei J, Teng M, Zeng L, Yan Z, Huang Z, Zhou Z, Wang P, Luo X, et al. Effects of Predominant Tree Species Mixing on Lignin and Cellulose Degradation during Leaf Litter Decomposition in the Three Gorges Reservoir, China. Forests. 2019; 10(4):360. https://doi.org/10.3390/f10040360
Chicago/Turabian StyleHe, Wei, Zhiyuan Ma, Jing Pei, Mingjun Teng, Lixiong Zeng, Zhaogui Yan, Zhilin Huang, Zhixiang Zhou, Pengcheng Wang, Xin Luo, and et al. 2019. "Effects of Predominant Tree Species Mixing on Lignin and Cellulose Degradation during Leaf Litter Decomposition in the Three Gorges Reservoir, China" Forests 10, no. 4: 360. https://doi.org/10.3390/f10040360
APA StyleHe, W., Ma, Z., Pei, J., Teng, M., Zeng, L., Yan, Z., Huang, Z., Zhou, Z., Wang, P., Luo, X., & Xiao, W. (2019). Effects of Predominant Tree Species Mixing on Lignin and Cellulose Degradation during Leaf Litter Decomposition in the Three Gorges Reservoir, China. Forests, 10(4), 360. https://doi.org/10.3390/f10040360