Caribou Conservation: Restoring Trees on Seismic Lines in Alberta, Canada



Abstract

1. Introduction
2. Materials and Methods
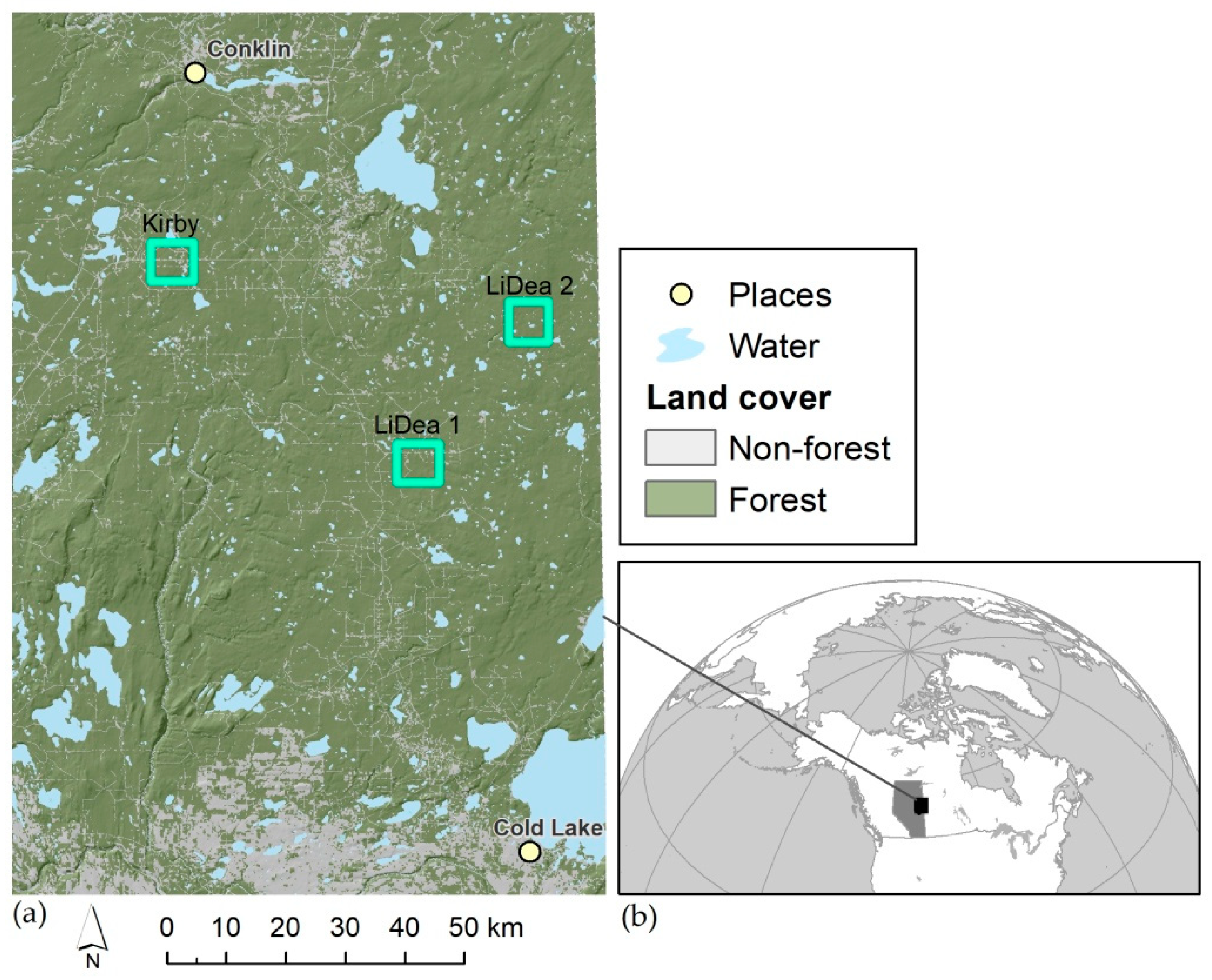
2.1. Study Area
2.2. Site selection and field methods
2.3. Regeneration Density by Restoration Area, Restoration Treatment, Plot Location, and Stand Characteristics
2.4. Regeneration Patterns Based on Seismic Line Characteristics
2.5. Tree Regeneration Height on Seismic Lines and Time Since Disturbance
3. Results
3.1. Overall Characteristics
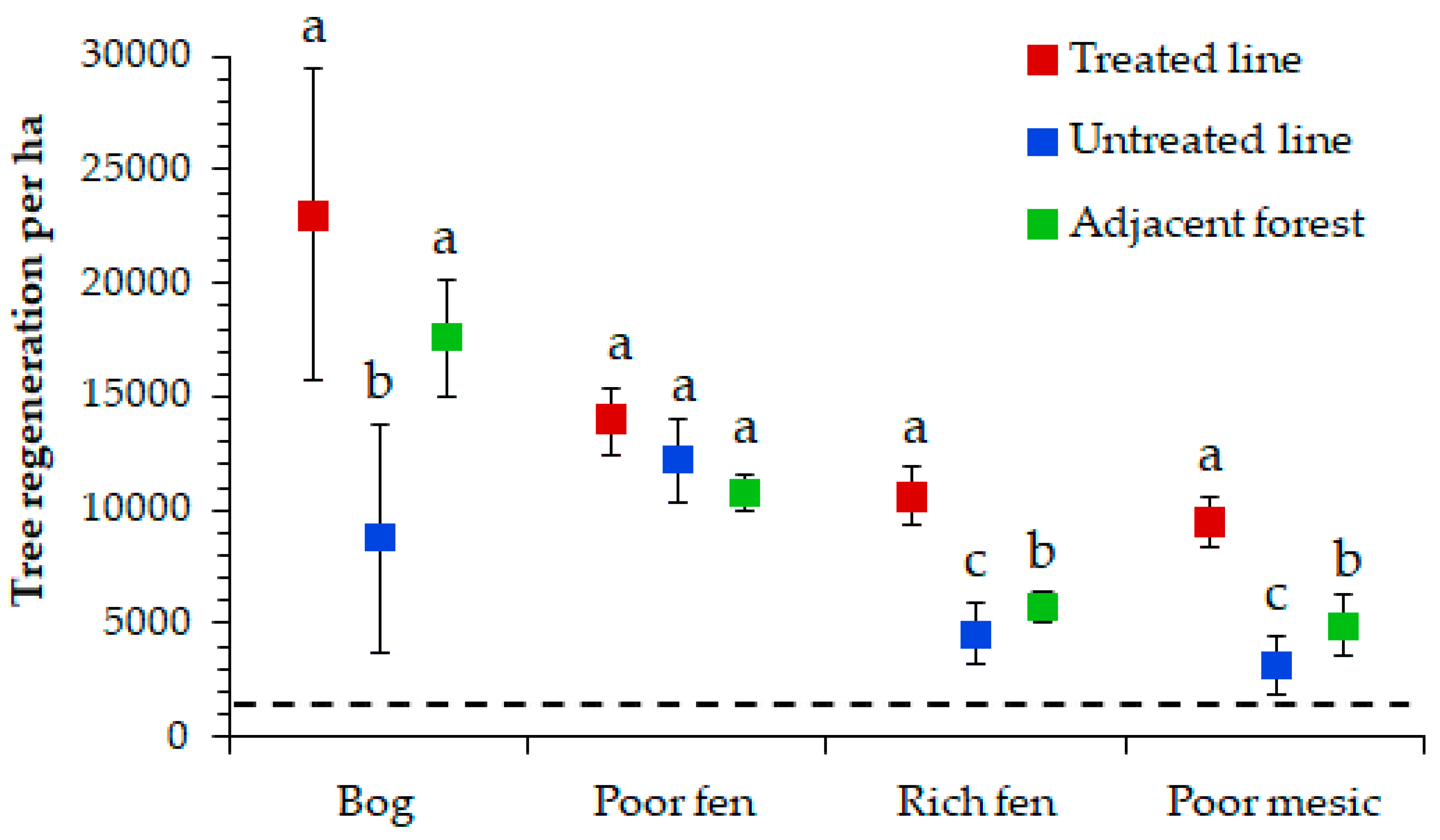
3.2. Restoration Effectiveness on Seismic Lines Versus Adjacent Forest
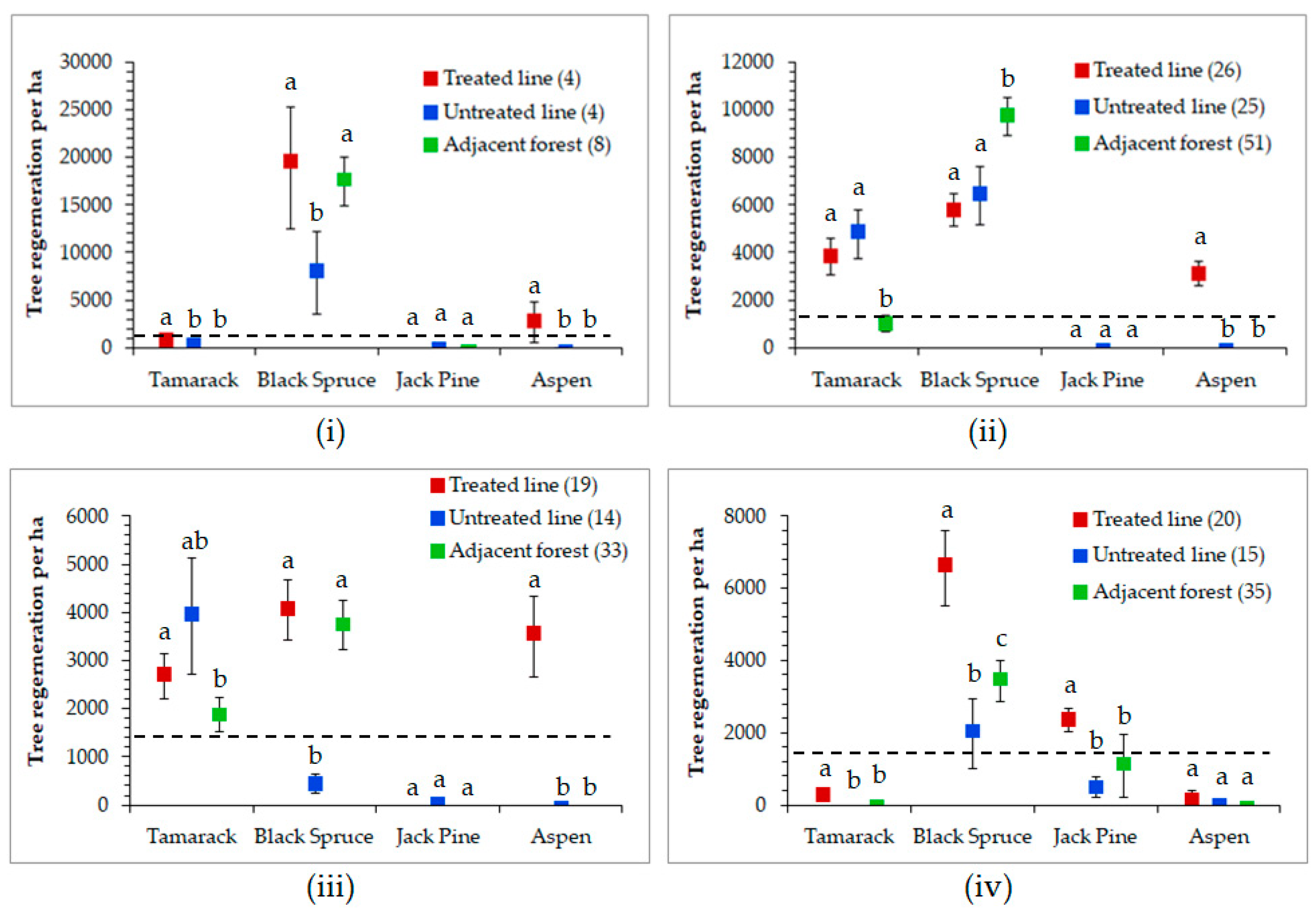
3.3. Seismic Line Regeneration Density
4. Discussion
4.1. Restoration Effectiveness on Seismic Lines Versus Adjacent Forest
4.2. Seismic Line Regeneration Density
4.3. Supplemental Information
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Density (stem/ha) | Bog | Poor Fen | ||||||
| Tamarack | Black Spruce | Jack Pine | Aspen | Tamarack | Black Spruce | Jack Pine | Aspen | |
| β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | |
| Constant (intercept) | 0 (40) | 16404 (29) *** | 0 (29) | 169 (31) *** | 7937 (12) *** | 3 (5) | 4 (11) | |
| Site (restoration area) | ||||||||
| LiDea 1 | 175 (40) ** | |||||||
| Kirby | ||||||||
| LiDea 2 | ||||||||
| Restoration treatment variables | ||||||||
| Mounding & Planting | 185 (96) | 247 (64) * | 836 (67) *** | −9 (48) | 26 (25) | −12 (10) | 1515 (22) *** | |
| Ripping & Planting | N/A | N/A | N/A | N/A | N/A | N/A | N/A | |
| Seismic line location variable | ||||||||
| Seismic line plot | 105 (79) | −71 (51) * | 44 (56) | 300 (36) *** | −51 (21) *** | 10 (9) | 19 (18) | |
| Stand variables | ||||||||
| Stand height (m) | −11 (5) * | |||||||
| Basal area of target tree | ||||||||
| Basal area of other tree species | ||||||||
| Model statistics | ||||||||
| n | 16 | 16 | 16 | 102 | 102 | 102 | 102 | |
| R2 (within) | 0.55 | 0.54 | 0.80 | 0.39 | 0.24 | 0.01 | 0.85 | |
| R2 (between) | 0.16 | 0.27 | 0.59 | 0.20 | 0.02 | 0.04 | 0.60 | |
| R2 (overall) | 0.41 | 0.41 | 0.73 | 0.28 | 0.13 | 0.02 | 0.76 | |
| Wald χ2 | 9.18 | 9.98 | 35.42 | 43.30 | 16.16 | 1.97 | 347.80 | |
| p-value | 0.010 | 0.007 | <0.001 | <0.001 | <0.001 | 0.373 | <0.001 | |
| Tree Density (stem/ha) | Rich Fen | Poor Mesic | ||||||
| Tamarack | Black Spruce | Jack Pine | Aspen | Tamarack | Black Spruce | Jack Pine | Aspen | |
| β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | |
| Constant (intercept) | 1974 (33) *** | 1263 (22) *** | 0 (5) | 49 (20) * | 13 (13) | 8588 (82) *** | 185 (30) *** | 13 (14) |
| Site (restoration area) | ||||||||
| LiDea 1 | 198 (41) ** | |||||||
| Kirby | ||||||||
| LiDea 2 | −56 (24) *** | |||||||
| Restoration treatment variables | ||||||||
| Mounding & Planting | 127 (53) | 671 (35) *** | −16 (11) | 1072 (36) *** | N/A | N/A | N/A | N/A |
| Ripping & Planting | N/A | N/A | N/A | N/A | 150 (27) *** | 438 (43) *** | 1837 (35) *** | −2 (29) |
| Seismic line location variable | ||||||||
| Seismic line plot | 17 (45) | −87 (30) *** | 19 (10) | 19 (32) | −16 (23) | −60 (35) ** | −9 (27) | 17 (26) |
| Stand variables | ||||||||
| Stand height (m) | −8 (4) * | |||||||
| Basal area of target tree | 9 (2) *** | 44 (16) * | 67 (14) *** | |||||
| Basal area of other tree species | −12 (2) *** | −5 (2) ** | ||||||
| Model statistics | ||||||||
| n | 33 | 33 | 33 | 33 | 35 | 35 | 35 | 35 |
| R2 (within) | 0.23 | 0.55 | 0.07 | 0.70 | 0.35 | 0.23 | 0.86 | 0.02 |
| R2 (between) | 0.51 | 0.76 | 0.04 | 0.68 | 0.22 | 0.35 | 0.24 | 0.33 |
| R2 (overall) | 0.38 | 0.67 | 0.06 | 0.69 | 0.28 | 0.30 | 0.57 | 0.19 |
| Wald χ2 | 36.72 | 126.82 | 3.82 | 140.05 | 26.07 | 26.33 | 198.10 | 15.48 |
| p-value | <0.001 | <0.001 | 0.150 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 |
| Tree Density (stem/ha) on Seismic Line Only | Bog | Poor fen | ||||||
| Tamarack | Black Spruce | Jack Pine | Aspen | Tamarack | Black Spruce | Jack Pine | Aspen | |
| β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | |
| Constant (intercept) | 34 (30) | 4809 (44) *** | 44 (71) | 1342 (100) *** | 5213 (27) *** | −3 (7) | 25 (22) | |
| Site (restoration area) | ||||||||
| LiDea 1 | 540 (45) *** | |||||||
| LiDea 2 | ||||||||
| Restoration treatment variables | ||||||||
| Mounding & Planting | 836 (114) * | 1490 (32) *** | ||||||
| Ripping & Planting | N/A | N/A | N/A | N/A | N/A | N/A | N/A | |
| Line characteristics | ||||||||
| Bearing | 1152 (53) ** | 389 (82) * | ||||||
| Stand variables | ||||||||
| Stand height (m) | −16 (5) ** | |||||||
| Basal area of target tree | 4 (2) * | |||||||
| Basal area of other tree species | ||||||||
| Ground Cover (%) | ||||||||
| Open water | −2 (1) * | |||||||
| Bryophyte | 2 (1) * | |||||||
| Sphagnum fuscum | 2 (1) * | |||||||
| Graminoid | −3 (1) * | |||||||
| Model statistics | ||||||||
| n | 8 | 8 | 8 | 51 | 51 | 51 | 51 | |
| R2 | 0.85 | 0.54 | 0.59 | 0.44 | 0.31 | 0.10 | 0.67 | |
| Prob > F | >0.001 | 0.038 | 0.026 | <0.001 | <0.001 | 0.021 | <0.001 | |
| RMSE | 0.25 | 0.36 | 0.47 | 0.54 | 0.33 | 0.19 | 0.43 | |
| Tree Density (stem/ha) on Seismic Line Only | Rich Fen | Poor Mesic | ||||||
| Tamarack | Black Spruce | Jack Pine | Aspen | Tamarack | Black Spruce | Jack Pine | Aspen | |
| β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | β (SE) | |
| Constant (intercept) | 404 (94) * | 163 (30) ** | 8 (8) | 186 (44) ** | 0 (22) | 798 (33) *** | 256 (29) *** | 322 (93) ** |
| Site (restoration area) | ||||||||
| LiDea 1 | ||||||||
| LiDea 2 | −81 (47) *** | |||||||
| Restoration treatment variables | ||||||||
| Mounding & Planting | 1170 (41) *** | 933 (48) *** | N/A | N/A | N/A | N/A | ||
| Ripping & Planting | N/A | N/A | N/A | N/A | 152 (30) ** | 450 (46) *** | 1265 (36) *** | |
| Line characteristics | ||||||||
| Bearing | −48 (33) * | |||||||
| Stand variables | ||||||||
| Stand height (m) | 18 (8) * | |||||||
| Basal area of target tree | ||||||||
| Basal area of other tree species | −6 (2) *** | |||||||
| Ground Cover (%) | ||||||||
| Open water | ||||||||
| Bryophyte | ||||||||
| Sphagnum fuscum | ||||||||
| Graminoid | ||||||||
| Model statistics | ||||||||
| n | 33 | 33 | 33 | 33 | 35 | 35 | 35 | 35 |
| R2 | 0.12 | 0.64 | 0.00 | 0.68 | 0.27 | 0.38 | 0.70 | 0.14 |
| Prob > F | 0.047 | <0.001 | 0.000 | <0.001 | 0.001 | <0.001 | <0.001 | 0.029 |
| RMSE | 0.54 | 0.42 | 0.18 | 0.48 | 0.34 | 0.48 | 0.37 | 0.35 |
References
- Arienti, M.C.; Cumming, S.G.; Krawchuk, M.A.; Boutin, S. Road network density correlated with increased lightning fire incidence in the Canadian western boreal forest. Int. J. Wildland Fire 2009, 18, 970–982. [Google Scholar] [CrossRef]
- Schneider, R.R.; Hauer, G.; (Vic) Adamowicz, W.L.; Boutin, S. Triage for conserving populations of threatened species: The case of woodland caribou in Alberta. Biol. Conserv. 2010, 143, 1603–1611. [Google Scholar] [CrossRef]
- Riva, F.; Acorn, J.H.; Nielsen, S.E. Localized disturbances from oil sands developments increase butterfly diversity and abundance in Alberta’s boreal forests. Biol. Conserv. 2018, 217, 173–180. [Google Scholar] [CrossRef]
- Roberts, D.; Ciuti, S.; Barber, Q.E.; Willier, C.; Nielsen, S.E. Accelerated seed dispersal along linear disturbances in the Canadian oil sands region. Sci Rep 2018, 8, 4828. [Google Scholar] [CrossRef] [PubMed]
- Dyer, S.J.; O’Neill, J.P.; Wasel, S.M.; Boutin, S. Avoidance of Industrial Development by Woodland Caribou. J. Wildl. Manag. 2001, 65, 531. [Google Scholar] [CrossRef]
- Dyer, S.J.; O’Neill, J.P.; Wasel, S.M.; Boutin, S. Quantifying barrier effects of roads and seismic lines on movements of female woodland caribou in northeastern Alberta. Can. J. Zool. 2002, 80, 839–845. [Google Scholar] [CrossRef]
- Government of Alberta: Alberta Environment and Parks. Woodland Caribou (Rangifer tarandus caribou); 2017. Available online: http://aep.alberta.ca/fish-wildlife/wild-species/mammals/deer/woodland-caribou.aspx (accessed on 10 December 2018).
- Lee, P.; Boutin, S. Persistence and developmental transition of wide seismic lines in the western Boreal Plains of Canada. J. Environ. Manag. 2006, 78, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Van Rensen, C.K.; Nielsen, S.E.; White, B.; Vinge, T.; Lieffers, V.J. Natural regeneration of forest vegetation on legacy seismic lines in boreal habitats in Alberta’s oil sands region. Biol. Conser. 2015, 184, 127–135. [Google Scholar] [CrossRef]
- Rettie, W.J.; Messier, F.F. Nordic Society Oikos Hierarchical Habitat Selection by Woodland Caribou: Its Relationship to Limiting Factors. Ecography (Cop.) 2000, 23, 466–478. [Google Scholar] [CrossRef]
- Vitt, D.H. AN OVERVIEW OF FACTORS THAT INFLUENCE THE DEVELOPMENT OF CANADIAN PEATLANDS. Mem. Entomol. Soc. Can. 1994, 126, 7–20. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Caners, R.T.; Ge, H. Re-establishment of hummock topography promotes tree regeneration on highly disturbed moderate-rich fens. J. Environ. Manag. 2017, 197, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Latham, A.D.M.; McCutchen, N.A.; Boutin, S. Invading white-tailed deer change wolf-caribou dynamics in northeastern Alberta. J. Wildl. Manag. 2011, 75, 204–212. [Google Scholar] [CrossRef]
- James, A.R.C.; Stuart-Smith, A.K. Distribution of Caribou and Wolves in Relation to Linear Corridors. J. Wildl. Manag. 2000, 64, 154. [Google Scholar] [CrossRef]
- Latham, A.D.M.; Latham, M.C.; Boyce, M.S.; Boutin, S. Movement responses by wolves to industrial linear features and their effect on woodland caribou in northeastern Alberta. Ecol. Appl. 2011, 21, 2854–2865. [Google Scholar] [CrossRef]
- Caners, R.T.; Lieffers, V.J. Divergent Pathways of Successional Recovery for In Situ Oil Sands Exploration Drilling Pads on Wooded Moderate-Rich Fens in Alberta, Canada. Restor. Ecol. 2014, 22, 657–667. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Rothwell, R.L. Rooting of peatland black spruce and tamarack in relation to depth of water table. Can. J. Bot. 1987, 65, 817–821. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Lieffers, V.J. Growth and foliar nutrient status of black spruce and tamarack in relation to depth of water table in some Alberta peatlands. Can. J. For. Res. 1990, 20, 805–809. [Google Scholar]
- Roy, V.; Bernier, P.-Y.; Plamondon, A.P.; Ruel, J.-C. Effect of drainage and microtopography in forested wetlands on the microenvironment and growth of planted black spruce seedlings. Can. J. For. Res. 1999, 29, 563–574. [Google Scholar] [CrossRef]
- Sutton, R.F. Mounding site preparation: A review of European and North American experience. New For. 1993, 7, 151–192. [Google Scholar] [CrossRef]
- Von der Gönna, M.A. Fundamentals of mechanical site preparation. For. Can. Serv. Pub. 1992, 1–28. [Google Scholar]
- Pyper, M.; Nishi, J.; McNeil, L. Linear Feature Restoration in Caribou Habitat: A Summary of Current Practices and a Roadmap for Future Programs; Canada’s Oil Sands Innov. Alliance (COSIA): Calgary, AB, Canada, December 2014. [Google Scholar]
- Alberta Agriculture and Forestry: Forestry Division. Alberta Forest Genetic Resource Management and Conservation Standards – third revision of STIA; Alberta Agriculture and Forestry: Forestry Division: Edmonton, AB, Canada, 2016.
- Downing, D.J.; Pettapiece, W.W. Natural Regions and Subregions of Alberta; Technical Report; Government of Alberta: Edmonton, AB, Canada, 2006.
- Christ, A. Mixed Effects Models and Extensions in Ecology with R. J. Stat. Soft. 2009, 32. [Google Scholar] [CrossRef]
- Kuhry, P.; Nicholson, B.J.; Gignac, L.D.; Vitt, D.H.; Bayley, S.E. Development of Sphagnum -dominated peatlands in boreal continental Canada. Can. J. Bot. 1993, 71, 10–22. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Pastor, J.; Harth, C.; Bridgham, S.D.; Updegraff, K.; Chapin, C.T. Response of bog and fen plant communities to warming and ware table manipulations. Ecology 2000, 81, 3464–3478. [Google Scholar] [CrossRef]
- Madec, C.; Walsh, D.; Lord, D.; Tremblay, P.; Boucher, J.-F.; Bouchard, S. Afforestation of Black Spruce Lichen Woodlands by Natural Seeding. North. J. Appl. For. 2012, 29, 191–196. [Google Scholar] [CrossRef]
- Mallik, A.; Kayes, I. Lichen mated seedbeds inhibit while moss dominated seedbeds facilitate black spruce (Picea mariana) seedling regeneration in post-fire boreal forest. For. Ecol. Manag. 2018, 427, 260–274. [Google Scholar] [CrossRef]
- Sutherland, B.; Foreman, F.F. Black spruce and vegetation response to chemical and mechanical site preparation on a boreal mixedwood site. Can. J. For. Res. 2000, 30, 1561–1570. [Google Scholar] [CrossRef]
- Löf, M.; Dey, D.C.; Navarro, R.M.; Jacobs, D.F. Mechanical site preparation for forest restoration. New For. 2012, 43, 825–848. [Google Scholar] [CrossRef]
- LaFleur, B.; Paré, D.; Fenton, N.J.; Bergeron, Y. Growth of planted black spruce seedlings following mechanical site preparation in boreal forested peatlands with variable organic layer thickness: 5-year results. Ann. For. Sci. 2011, 68, 1291–1302. [Google Scholar] [CrossRef]
- Bubier, J.L. Patterns of Picea mariana (Black Spruce) Growth and Raised Bog Development in Victory Basin, Vermont. Bull. Torrey Bot. Club 1991, 118, 399. [Google Scholar] [CrossRef]
- Boucher, D.; Gauthier, S.; De Grandpré, L. Structural changes in coniferous stands along a chronosequence and a productivity gradient in the northeastern boreal forest of Québec. Écoscience 2006, 13, 172–180. [Google Scholar] [CrossRef]
- Sirois, L. Impact of Fire on Picea-Mariana and Pinus-Banksiana Seedlings in Sub-Arctic Lichen Woodlands. J. Veg. Sci. 1993, 4, 795–802. [Google Scholar] [CrossRef]
- Filicetti, A.T.; Nielsen, S.E. Fire and forest recovery on seismic lines in sandy upland jack pine (Pinus banksiana) forests. For. Ecol. Manag. 2018, 421, 32–39. [Google Scholar] [CrossRef]
- Triisberg, T.; Karofeld, E.; Liira, J.; Orru, M.; Ramst, R.; Paal, J. Microtopography and the properties of residual peat are convenient indicators for restoration planning of abandoned extracted peatlands. Restor. Ecol. 2014, 22, 31–39. [Google Scholar] [CrossRef]



| Stand Variable | Minimum | Median | Maximum | Mean (S.E.) |
|---|---|---|---|---|
| Age | 5 | 52 | 165 | 56.6 (1.8) |
| Height | 2 | 9.5 | 25.2 | 10.3 (0.3) |
| Basal area (m2/ha) | 0 | 12 | 44 | 13.7 (0.5) |
| Tree stems per ha (DBH < 1 cm) | ||||
| Seismic line | 0 | 9000 | 39,333 | 10,186 (719) |
| Adjacent stand | 0 | 6000 | 48,333 | 8236 (631) |
| Tree stems per ha (DBH ≥ 1 cm) | ||||
| Seismic line | 0 | 0 | 2500 | 68 (29) |
| Adjacent stand | 1333 | 14,333 | 45,000 | 14,587 (743) |
| Tree Density (stems/ha) | Bog | Poor Fen | Rich Fen | Poor Mesic |
|---|---|---|---|---|
| β (SE) | β (SE) | β (SE) | β (SE) | |
| Constant (intercept) | 16,450 (30) *** | 7743 (14) *** | 6454 (20) *** | 2710 (20) *** |
| Site (restoration area) | ||||
| LiDea 1 | 41 (17) * | |||
| Kirby | ||||
| LiDea 2 | −44 (24) ** | |||
| Restoration treatment | ||||
| Mounding & Planting | 300 (67) ** | 34 (24) | 271 (37) *** | N/A |
| Ripping & Planting | N/A | N/A | N/A | 731 (38) *** |
| Seismic line location | ||||
| Seismic line plot | −70 (52) ** | −16 (20) | −55 (32) ** | −63 (30) *** |
| Model statistics | ||||
| n | 16 | 102 | 66 | 70 |
| R2 (within) | 0.52 | 0.04 | 0.17 | 0.51 |
| R2 (between) | 0.29 | 0.12 | 0.44 | 0.21 |
| R2 (overall) | 0.41 | 0.08 | 0.31 | 0.32 |
| Wald χ2 | 9.82 | 8.30 | 27.53 | 43.72 |
| p-value | 0.007 | 0.040 | <0.001 | <0.001 |
| Tree Density (stems/ha) on Seismic Line Only | Bog | Poor Fen | Rich Fen | Poor Mesic |
|---|---|---|---|---|
| β (SE) | β (SE) | β (SE) | β (SE) | |
| Constant (intercept) | 5024 (45) *** | 13,012 (22) *** | 534 (80) *** | 1224 (32) *** |
| Restoration treatment | ||||
| Mounding & Planting | 70 (26) * | 206 (49) ** | N/A | |
| Ripping & Planting | N/A | N/A | N/A | 540 (44) *** |
| Line characteristics | ||||
| Bearing | 477 (84) * | |||
| Stand variables | ||||
| Stand height (m) | 19 (8) * | |||
| Ground Cover (%) | ||||
| Lichen | −5 (3) * | |||
| Graminoid | −5 (1) *** | |||
| Model statistics | ||||
| n | 8 | 51 | 33 | 35 |
| R2 | 0.58 | 0.30 | 0.39 | 0.44 |
| Prob > F | 0.028 | <0.001 | <0.001 | <0.001 |
| RMSE | 0.36 | 0.35 | 0.48 | 0.46 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filicetti, A.T.; Cody, M.; Nielsen, S.E. Caribou Conservation: Restoring Trees on Seismic Lines in Alberta, Canada. Forests 2019, 10, 185. https://doi.org/10.3390/f10020185
Filicetti AT, Cody M, Nielsen SE. Caribou Conservation: Restoring Trees on Seismic Lines in Alberta, Canada. Forests. 2019; 10(2):185. https://doi.org/10.3390/f10020185
Chicago/Turabian StyleFilicetti, Angelo T., Michael Cody, and Scott E. Nielsen. 2019. "Caribou Conservation: Restoring Trees on Seismic Lines in Alberta, Canada" Forests 10, no. 2: 185. https://doi.org/10.3390/f10020185
APA StyleFilicetti, A. T., Cody, M., & Nielsen, S. E. (2019). Caribou Conservation: Restoring Trees on Seismic Lines in Alberta, Canada. Forests, 10(2), 185. https://doi.org/10.3390/f10020185