1. Introduction
An increasing number of plant diseases have been observed over the past years, most likely initiated by increasing summer temperatures and drought events throughout the past decade [
1]. Besides this accumulating number of biotic pests and diseases, in May 2018, a new phenomenon was observed in
Acer, which was reported by a forest engineer from the forest enterprise Große Hermannsberg, Thuringia, Germany (longitude, 50.7123; latitude 10.6035) at 700 m above sea level, who stated that “
Acer has a blood pressure that is too high. The bark cracks along the trees”. Despite much experience in this area, for the first time in his life, he witnessed an event involving virtually hundreds of young
Acer trees experiencing major bark damage. The bark cracked open in a single long stripe (see
Figure 1A). In some trees, the crack emerged from the base to the crown. To our knowledge, this type of damage has not previously been described [
2,
3]. It is possible that this damage was assumed to be frost-damage in the past, but this damage incident became apparent during warm temperatures of May.
The following sections discuss possible causes of this event. As a main hypothesis, we considered late frost-damage to the expanding xylem cells next to the cambial zone, which initiated growth before the frost event. Desiccation of the active growth layer at the same time that xylem sap flow and bleeding began, but when the phloem was still dormant, could have interacted with the late-frost event. It emerged that the cambium and cortex were more frost- and drought-tolerant than the expanding xylem cells.
2. Materials and Methods
2.1. Weather Conditions in Spring 2018
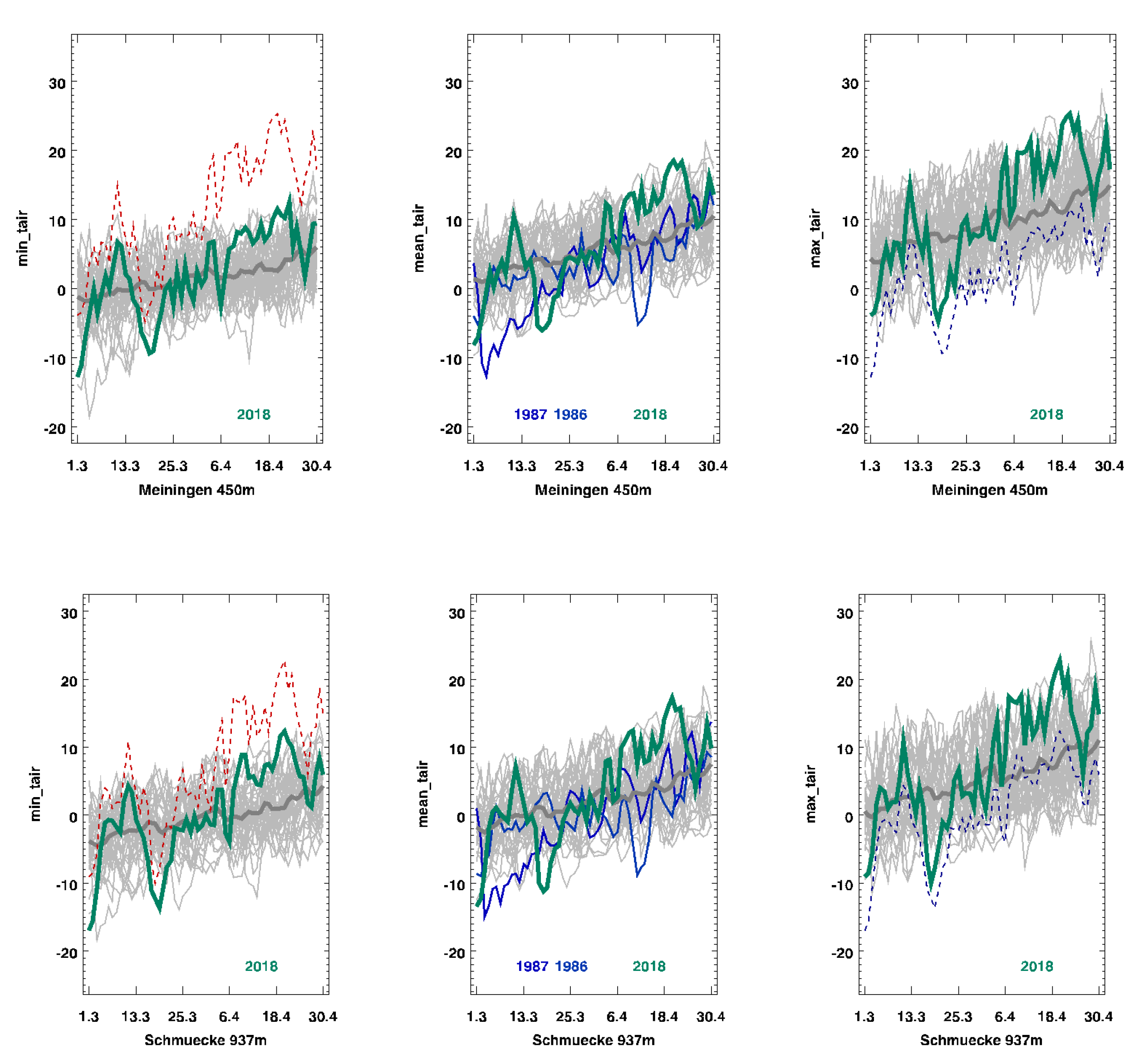
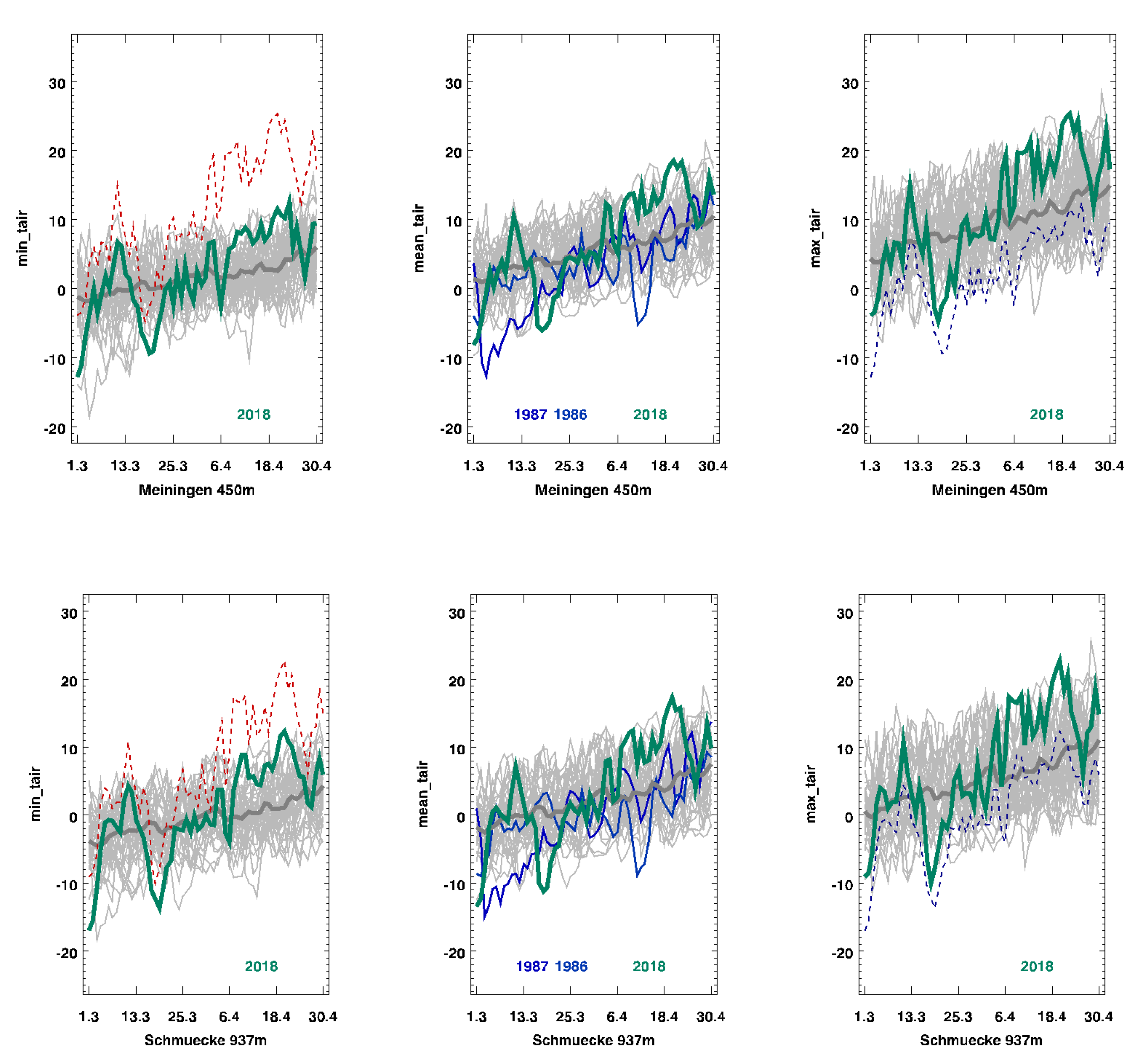
We based our climatic information on the records of two national weather service stations, namely, the station at “Schmuecke”, which was 12 km away from the site but at a higher elevation (937m above sea level), and “Meiningen”, which was 18 km away but at a lower elevation (450 m above sea level).
In 2018, following a relatively mild winter with snow cover, the soil was not frozen. There was a short cold period of 12 ice-days at Schmuecke and 8 ice-days at Meiningen, with maximum temperatures below −5 °C in late February and early March (
Figure 2, green line). The minimum temperatures were between −11 °C (Meiningen) and −14 °C (Schmuecke) above the snow cover. This frost event was followed by a warm period, with maximum daily air temperatures above 10 °C (
Figure 2, green line). No xylem bleeding was observed in February before the frost event. However, the xylem sap flow (bleeding from cut xylem) in
Acer began during the warm period in early March following the frost event, as observed while testing whether it was time for pruning. This warm period was followed by a second cold event in late March, when the minimum temperatures dropped to −6 °C (Meiningen) and −11 °C (Schmuecke) for 3–4 days (
Figure 2, green line). The weather became warm again a week later, when temperatures once again reached more than 10 °C, and
Acer started to resume bleeding (
Table 1).
The five-year average daily temperatures during March to April increased by 1.8 °C between 1979 and 2018. The standard deviation of the average daily temperature in March and April was 4.2 at Meiningen and 4.1 at Schmuecke, as averaged over the 40 years of observation. Compared to this average, 2018 showed the highest standard deviation of daily temperatures during the observation time, specifically, 7.0 at Meiningen and 7.5 at Schmuecke. It seems possible that with ongoing climate change, this pattern of a very warm March followed by cold events in April may become more pronounced in the future.
2.2. Study Site
The observations were carried out at the forest enterprise “Forstbetrieb Hermannsberg”, which consists of an area of about 200 ha covering a carboniferous volcano (867 m above sea level; longitude 50.7123; latitude: 10.6035) that produces acidic bedrock of quartz-porphyry and tuff. The upper elevation (< 700 m elevation) is covered by coniferous forest. The lower elevation (550–700 m) is covered by broad-leaved forest dominated by Acer pseudoplatanus, Sorbus aucuparia, Fraxinus excelsior, Populus tremula, Prunus avium, Betula pendula, Quercus robur, and Fagus sylvatica, with Acer being of greatest abundance and Fagus the least.
Observations were made on the north and northwest sides of the mountain, where old growth Acer stands (>200-year-old trees) exist from natural succession and where meadows were afforested by Acer (2.5 m × 2 m planting distance) about 15 years ago. The damage occurred in stands of natural succession as well as in hedge rows in meadows and afforestations, mainly in stems that were reached by solar radiation. Selected trees were pruned as future trees a year before the damage. In the year of the damage, single branches were pruned in late winter to see if xylem bleeding had started in order to allow for further pruning of twin stems. None of the damaged trees were pruned in 2018.
It should be noted that following this damage, we were informed that bark damage also occurred on calcareous sites (limestone) and sandstone. The damage appeared to be independent of geological conditions and management, but it was related to sun exposure (stands with low levels of cover or at the forest edges).
The observations in this study were made in 2019 at the Hermannsberg site, where the damage was quantified in the field using measurements of the dimensions of the affected trees and their damage. Tissue microscopy of wood samples was carried out at the Swiss Federal Research Institute WSL, Birmensdorf, Switzerland, following instructions by Schweingruber et al. [
4,
5]. Fungi were identified microscopically by the morphology of conidia and conidiophores (Ascomycetes [
6,
7]), by the morphology of basiodiocarps (Basiodiomycetes [
8,
9,
10,
11,
12]), and by DNA sequencing of fungal barcodes (
Cladosporum, Neocucurbitaria). Bark beetle galleries were analysed and identified by experts at WSL, Birmensdorf and remainings of individuals were identified by barcoding. Climate data were retrieved from the National Weather Service database (
https://opendata.dwd.de/climate_environment/CDC/observations_germany/climate/daily/kl/).
4. Discussion
In this study, we reported a phenomenon of bark-cracking in spring following a very cold event after xylem sap flow had already started during warm temperatures prior to the cold event.
The event of the spring bark-splitting of
Acer indicated that climate change may not only mean a rise in temperatures or a change in precipitation, but also an increase in the variation of springtime temperatures. The temperate zone of Europe will always be in the reach of arctic air masses, especially in spring, when the air masses across the Atlantic tend to be unstable. Thus, late and severe frost may limit growth starting earlier. This may not only be important for
Acer, but also for other deciduous, broad-leaved tree species, in which we observed similar bark damage (mainly oak and ash). Frost-ribs in other species was previously interpreted as damage resulting from winter temperatures and radial cracking of the xylem rather than damage by late frost, which usually affects a narrow cell layer in a tangential direction [
2].
It remains uncertain if the damage was caused by frost per se, or by an interaction of high osmotic concentrations in the bark and the cambium followed by a desiccation of the expanding xylem cells after the frost event. Most likely, the osmotic effect of the bleeding sap in
Acer trees was not sufficient to cause the damage in the cambial cells. These may have an osmotic concentration of about one molar [
16], however, expanding xylem cells could well be affected. Thus, a frost event with temperatures below −5 °C remains the most likely factor to cause this damage. No fungal cells were detected at the end-point of the damage.
It might also be possible that pruning enhanced the damage. At our site, the pruning of selected trees took place a year prior to the damage. In the year of damage, only a few branches were cut to inspect the early timing of xylem sap flow. However, no pruning took place in 2018 due to early bleeding. The damage also took place in unpruned trees. Thus, pruning may have enhanced the damage via microclimatic effects, allowing solar radiation to reach stems that were previously shaded.
We are aware that our interpretations of this observation are retrospective, but it is difficult to investigate this damage experimentally due to the apparent relationship between the damage and the activation of growth prior to the frost event. Temperatures in March 2018 following a warm winter activated xylem flow, and this period was followed by a cold spell with temperatures below −10 °C. We know from inquiries by several forest managers that the damage occurred regionally on a wide scale, covering North Bavaria and Southern Thuringia (500 km × 500 km), but no systematic investigation regarding the regional extent of the observation was made.
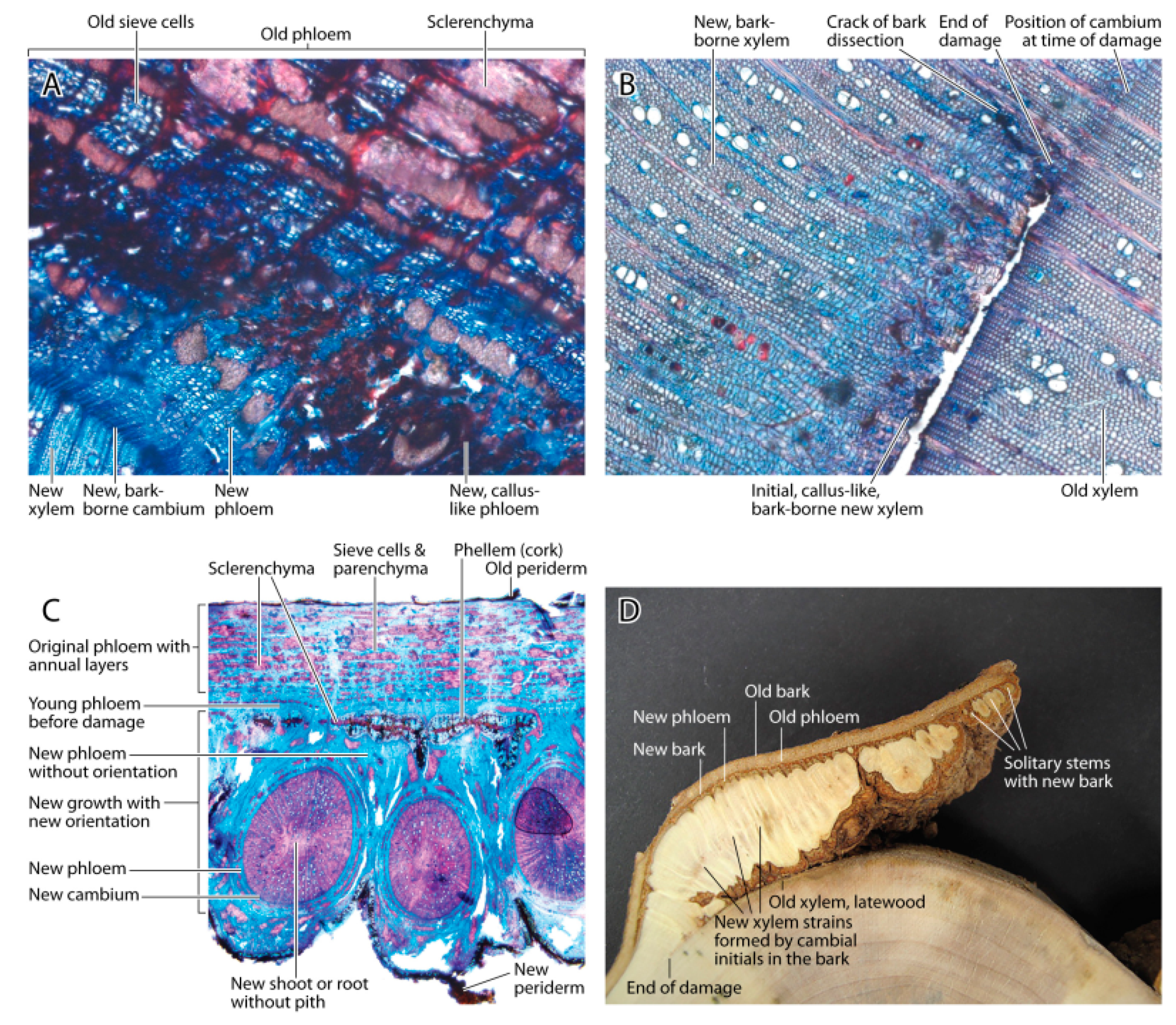
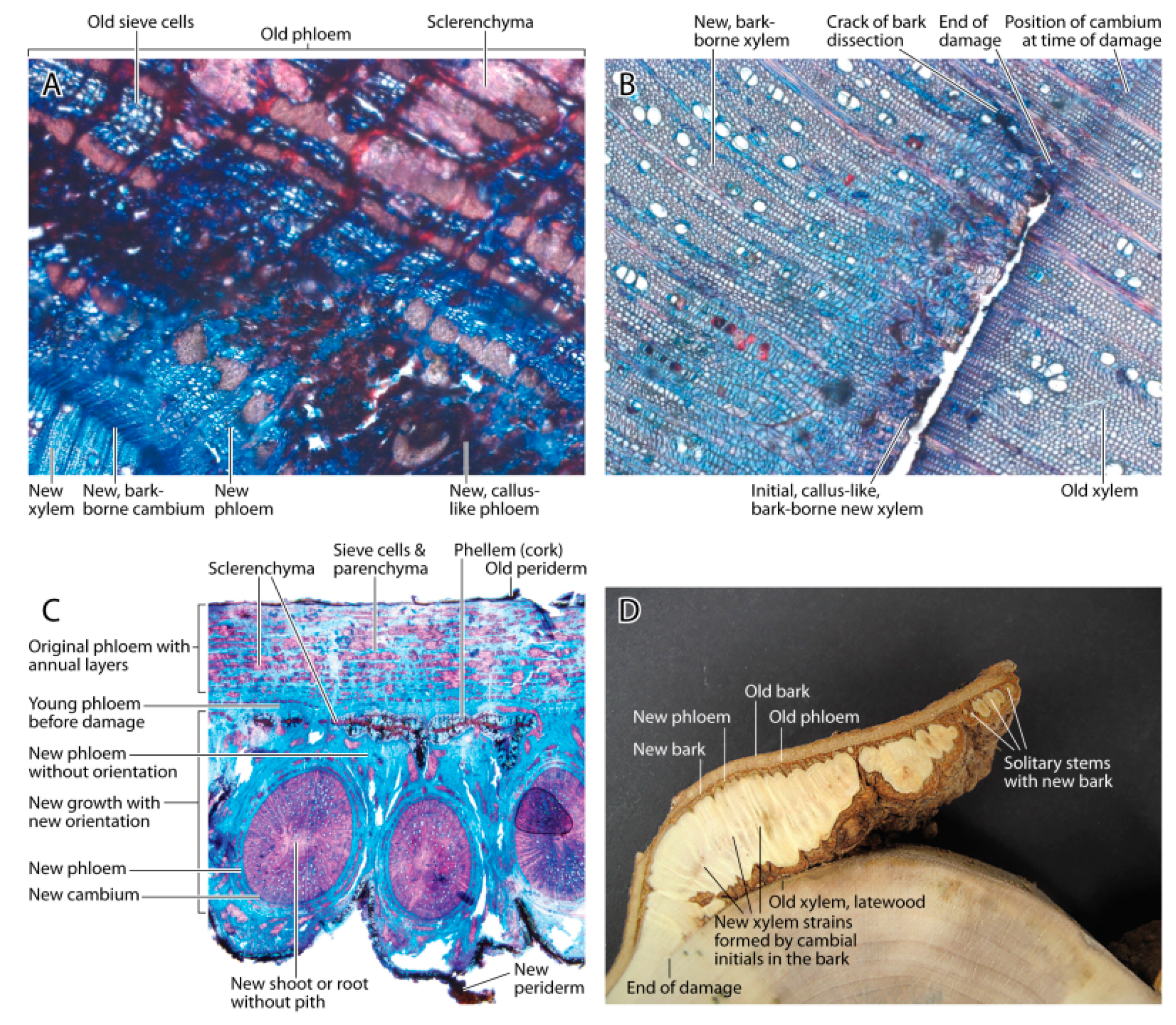
Based on the anatomical evidence, the damage occurred in the expanding xylem cells and not in the primary cambium. The cambium and the cortex appeared to have higher frost tolerances due to higher osmotic cell concentrations of the solutes than the expanding xylem cells. The surviving cambial cells remained attached to the bark rather than the xylem, and these cells were the basis for wound healing and new wood growth. The range of cell types formed after this damage was surprisingly large. Initially, callus-like lignified cells formed after cell division on the phloem and xylem sides of the new secondary cambium. It remains unclear if the cambial cells attached to the bark initially received growth hormones mainly from the shoot, because is it possible that phloem-flow was reversed at the time of growth initiation [
17]. The functional phloem and xylem elements were formed only after water and hormonal transport resumed from the root. Thus, the balance of growth hormones produced by the shoot and roots cause the observed recovery of wood formation [
4,
18].
The damage resembled the well-known observation of bark-cracking and “frost-plates” in fruit trees. In orchards, stems are stained white by liming to reduce solar irradiance [
19]. In orchards and ornamental tree nurseries, this kind of damage was explained by the desiccation of the bark at a time of inactive xylem water-flow. However, the time of the damage observed in orchards coincides with the initiation of growth. Thus, also in these cases, the damage could be due to a combination of desiccation and late frost, which was reduced by liming the stems.
This damage caused major disorder in the stem. In fact, after one year, the damage extended deeper into the wood than was visible from the outside. Apparently, the tree isolated the segment of wood where the ray cells were no longer connected to the phloem. The black border in the wood is generally assumed to be produced in the presence of fungi [
20], but in this case it appeared to be a plant response to separate the active and inactive xylem [
5,
20,
21,
22,
23]. We found phenolic deposits in ray cells in the absence of fungal hyphae.
Since the bark initiated the growth of independent stems, where each stem has the opportunity to form its own bark, it is possible that these stems may never fully merge again. Thus, we expect a frost-rib-like feature to remain visible in older trees until they reach a late age. It seems unlikely that these trees will ever be able to produce stems of veneer-quality.
The more severe damage to the tree was observed in the wedge-like separation of the damaged wood segment from the living wood. The secondary drying cracks of this “dead” part of the wood were entry points for several fungi, which became established near the pith in the tree center. Even if the wounds close in the future, the trees will probably continue to rot in the center. It is most likely that these trees will break in a few years.
In terms of management, it may be necessary to cut these damaged trees and start again with a new sprout, if high-quality timber is the objective of management. Since the damage occurred in young trees only, and since the variation in springtime temperatures appears to have increased, starting with new sprouts may not even be a solution. Since the damage exposed the differentiated xylem, there were no wound-responses of defense against fungal and bark beetle attacks by the wood. Thus, these trees could become “habitat-trees” for saproxylics in managed forests. In the future, we may have to live with frost-scar-like structures in several types of heartwood tree or avoid exposure of the stem to solar radiation by establishing dense stands.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}