How Long Do Wood Parenchyma Cells Live in the Stem of a Scots Pine (Pinus sylvestris L.)? Studies on Cell Nuclei Status along the Radial and Longitudinal Stem Axes


Abstract
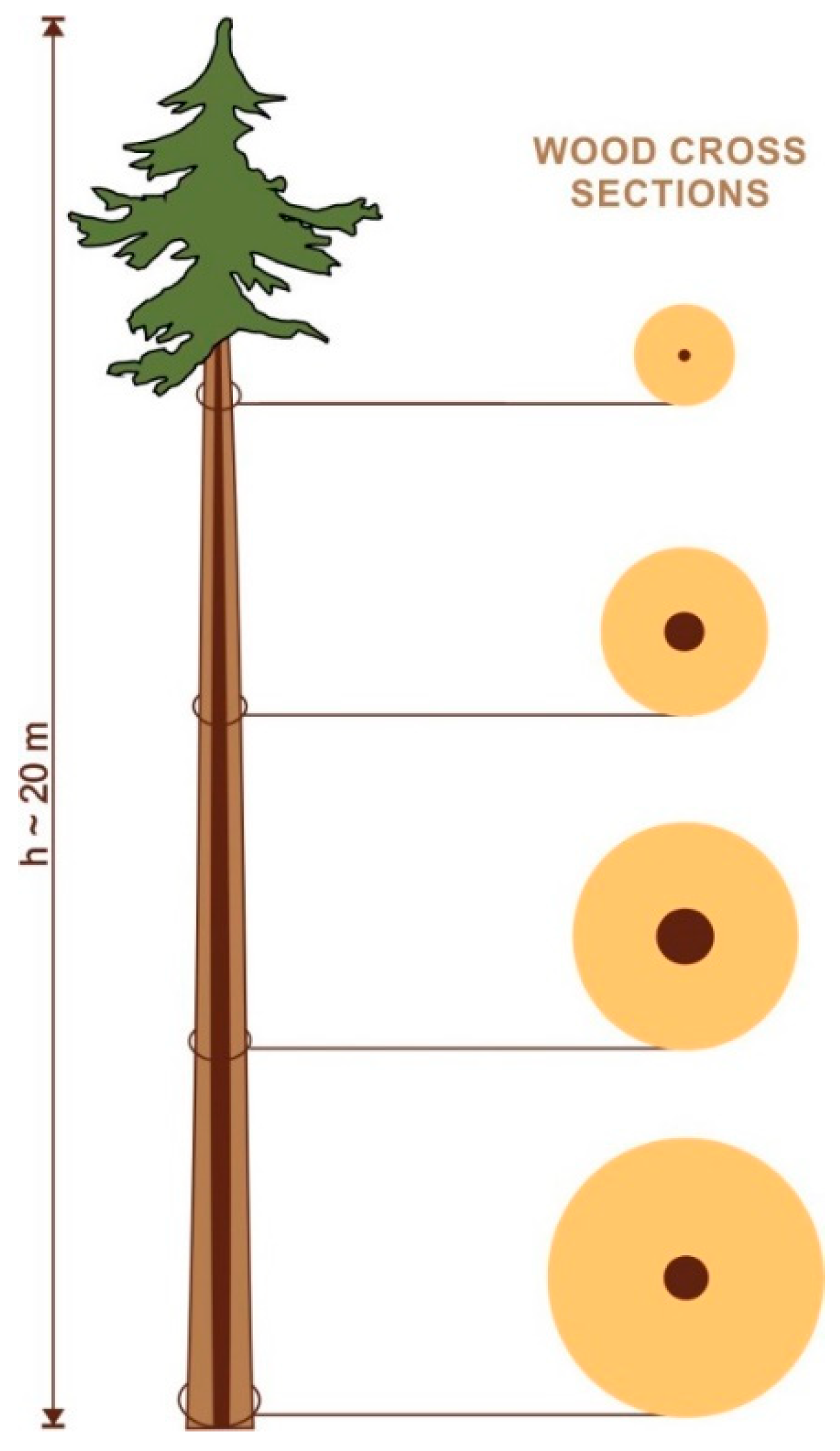
1. Introduction
2. Results and Discussion
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Committee on Nomenclature International Association of Wood Anatomists (IAWA). Multilingual Glossary of Terms Used in Wood Anatomy; Verlagsanstalt Buchdruckerei Konkordia: Winterthur, Switzerland, 1964. [Google Scholar]
- Taylor, A.M.; Gartner, B.; Morrell, J. Heartwood formation and natural durability—A review. Wood Fiber Sci. 2002, 4, 587–611. [Google Scholar]
- Panshin, A.J.; de Zeeuw, C. Textbook of Wood Technology: Structure, Identification, Properties, and Uses of the Commercial Woods of the United States and Canada, 4th ed.; McGraw-Hill Series in Forest Resources: New York, NY, USA, 1980; ISBN 9780070484412. [Google Scholar]
- Wodzicki, T.J. Mechanism of xylem differentiation in Pinus silvestris L. J. Exp. Bot. 1971, 22, 670–687. [Google Scholar] [CrossRef]
- Białobok, S.; Boratyński, A.; Bugała, W. Biology of Scots Pine Sorus; Institute of Dendrology: Poznań-Kórnik, Poland, 1993; ISBN 83-85599-21-5. [Google Scholar]
- Leyton, L.; Juniper, B.E. Cuticle structure and water relations of pine needles. Nature 1963, 198, 770–771. [Google Scholar] [CrossRef]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood formation in trees. Plant Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Sokołowska, K. Symplasmic transport in wood: The importance of living xylem cells. In Symplasmic Transport in Vascular Plants; Sokołowska, K., Sowiński, P., Eds.; Springer Science-Business Media: New York, NY, USA, 2013; pp. 101–132. [Google Scholar]
- Spicer, R. Symplasmic networks in secondary vascular tissues: Parenchyma distribution and activity supporting long-distance transport. J. Exp. Bot. 2014, 65, 1829–1848. [Google Scholar] [CrossRef]
- Wodzicki, T.J.; Brown, C.L. Cellular differentiation of the cambium in the Pinaceae. Bot. Gaz. 1973, 134, 139–146. [Google Scholar] [CrossRef]
- Rathgeber, C.B.K.; Cuny, H.E.; Fonti, P. Biological basis of tree-ring formation: A crash course. Front. Plant Sci. 2016, 7, 734. [Google Scholar] [CrossRef]
- Wodzicki, T.J. Natural factors affecting wood structure. Wood Sci. Technol. 2001, 35, 5–26. [Google Scholar] [CrossRef]
- Zajączkowski, S.; Wodzicki, T.J. Auxin and plant morphogenesis—A model of regulation. Acta Soc. Bot. Pol. 1978, 47, 233–243. [Google Scholar] [CrossRef]
- Wolpert, L. Positional information and the spatial pattern of cellular differentiation. J. Theor. Biol. 1969, 25, 1–47. [Google Scholar] [CrossRef]
- Zajączkowski, S.; Wodzicki, T.J.; Bruinsma, J. A possibile mechanizm for whole-plant morphogenesis. Physiol. Plant. 1983, 57, 306–310. [Google Scholar] [CrossRef]
- Zajączkowski, S.; Wodzicki, T.J.; Romberger, J.A. Auxin waves and plant morphogenesis. In Encyclopedia of Plant Physiology; Scott, T.K., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; Volume 10, pp. 244–262. [Google Scholar]
- Uggla, C.; Melerowicz, E.J.; Sundberg, B. Indole-3-acetic acid controls cambial growth in Scots pine by positional signaling. Plant Physiol. 1998, 117, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Hillis, W.E. Heartwood and Tree Exudates; Springer: Berlin/Heidelberg, Germany, 1987; ISBN 978-3-642-72536-4. [Google Scholar]
- Kuroda, K.; Yamashita, K.; Fujiwara, T. Cellular level observation of water loss and the refilling of tracheids in the xylem of Cryptomeria japonica during heartwood formation. Trees 2009, 23, 1163–1172. [Google Scholar] [CrossRef]
- Nakada, R.; Fukatsu, E. Seasonal variation of heartwood formation in Larix kaempferi. Tree Physiol. 2012, 32, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- Spicer, R. Senescence in secondary xylem: Heartwood formation as an active developmental program. In Vascular Transport in Plants; Holbrook, N.M., Zwieniecki, M.A., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; pp. 457–475. [Google Scholar]
- Fujii, T.; Harada, H.; Saiki, H. The layered structure of ray parenchyma secondary wall in the wood of 49 Japanese angiosperm species. Mokuzai Gakkaishi 1979, 25, 251–257. [Google Scholar]
- Krahmer, R.L.; Côté, W.A., Jr. Changes in coniferous wood cells associated with heartwood formation. TAPPI 1963, 46, 42–49. [Google Scholar]
- Bamber, R.K. Heartwood, its function and formation. Wood Sci. Technol. 1976, 10, 1–8. [Google Scholar] [CrossRef]
- Nobuchi, T.; Sato, T.; Iwata, R.; Harada, H. Season of heartwood formation and the related cytological structure of ray parenchyma cells in Robinia pseudoacaccia. Mokuzai Gakkaishi 1984, 30, 636–638. [Google Scholar]
- Bito, N.; Nakada, R.; Fukatsu, E.; Matsushita, Y.; Fukushima, K.; Imai, T. Clonal variation in heartwood norlignans of Cryptomeria japonica: Evidence for independent control of agatharesinol and sequirin C biosynthesis. Ann. For. Sci. 2011, 68, 1049–1056. [Google Scholar] [CrossRef]
- Bush, D.; McCarthy, K.; Meder, R. Genetic variation of natural durability traits in Eucalyptus cladocalyx (sugar gum). Ann. For. Sci. 2011, 68, 1057–1066. [Google Scholar] [CrossRef]
- Reyes-Garcia, C.; Andrade, J.J. Sapwood to heartwood ratio affects whole-tree water use in dry forest legume and non-legume trees. Trees 2012, 26, 1317–1330. [Google Scholar] [CrossRef]
- Stokes, A.; Berthier, S. Irregular heartwood formation in Pinus pinaster Ait. is related to eccentric, radial, stem growth. For. Ecol. Manag. 2000, 135, 115–121. [Google Scholar] [CrossRef]
- Climent, J.M.; Gil, L.; Pardos, J.A. Xylem anatomical traits related to resinous heartwood formation in Pinus canariensis Sm. Trees 1998, 12, 139–145. [Google Scholar] [CrossRef]
- Gartner, B.L. Patterns of xylem variation within a tree and their hydraulic and mechanical consequences. In Tree Stems; Gartner, B.L., Ed.; Academic Press: London, UK, 1995; pp. 125–149. [Google Scholar]
- Huang, Z.; Meilan, R.; Woeste, K.A. Knat3-like homeobox gene from Juglans nigra L., JnKNAT3-like, highly expressed during heartwood formation. Plant Cell Rep. 2009, 28, 1717–1724. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.J.; Paasela, T.; Harju, A.; Venäläinen, M.; Paulin, L.; Auvinen, P.; Kärkkäinen, K.; Teeri, T.H. Developmental changes in Scots pine transcriptome during heartwood formation. Plant Physiol. 2016, 172, 1403–1417. [Google Scholar] [CrossRef] [PubMed]
- Nakaba, S.; Kubo, T.; Funada, R. Differences in patterns of cell death between ray parenchyma cells and ray tracheids in the conifers Pinus densiflora and Pinus rigida. Trees 2008, 22, 623–630. [Google Scholar] [CrossRef]
- Uggla, C.; Magel, E.; Moritz, T.; Sundberg, B. Function and dynamics of auxin and carbohydrates during earlywood/latewood transition in Scots pine. Plant Physiol. 2001, 125, 2029–2039. [Google Scholar] [CrossRef]
- Bhalerao, R.; Bennett, M.J. The case for morphogens in plants. Nat. Cell Biol. 2003, 5, 939–943. [Google Scholar]
- Shain, L.; Mackay, J.F.G. Seasonal fluctuations in respiration of aging xylem in relation to heartwood formation in Pinus radiata. Can. J. Bot. 1973, 51, 737–741. [Google Scholar] [CrossRef]
- Nakaba, S.; Sano, Y.; Kubo, T.; Funada, R. The positional distribution of cell death of ray parenchyma in a conifer, Abies sachalinensis. Plant Cell Rep. 2006, 25, 1143–1148. [Google Scholar] [CrossRef]
- Nakaba, S.; Yamagishi, Y.; Sano, Y.; Funada, R. Temporally and spatially controlled death of parenchyma cells is involved in heartwood formation in pith regions of branches of Robinia pseudoaccaia var. inermis. J. Wood Sci. 2012, 58, 69–76. [Google Scholar] [CrossRef]
- Spicer, R.; Holbrook, N.M. Perenchyma cell respiration and survival in secondary xylem: Does metabolic activity decline with cell age? Plant Cell Environ. 2007, 30, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Climent, J.; Gil, L.; Pardos, J. Heartwood and sapwood development and its relationship to growth and environment in Pinus canariensis Chr.Sm. ex DC. For. Ecol. Manag. 1993, 59, 165–174. [Google Scholar] [CrossRef]
- Song, K.; Liu, B.; Jiang, X.; Yin, Y. Cellular changes of tracheids and ray parenchyma cells from cambium to heartwood in Cunninghamia lanceolata. J. Trop. For. Sci. 2011, 23, 478–487. [Google Scholar]
- Climent, J.; Chambel, M.R.; Gil, L.; Pardos, J.A. Vertical heartwood variation patterns and prediction of heardwood volume in Pinus canariensis Sm. For. Ecol. Manag. 2003, 174, 203–211. [Google Scholar] [CrossRef]
- Berthier, S.; Kokutse, D.A.; Stokes, A.; Fourcaud, T. Irregular heartwood formation in Maritime pine (Pinus pinaster Ait.): Consequences for biomechanical and hydraulic tree functioning. Ann. Bot. 2001, 87, 19–25. [Google Scholar] [CrossRef]
- Mencuccini, M.; Grace, J.; Fioravanti, M. Biomechanical and hydraulic determinants of tree structure in Scots pine: Anatomical characteristics. Tree Physiol. 1997, 17, 105–113. [Google Scholar] [CrossRef]
- Nakaba, S.; Begum, S.; Yamagishi, Y.; Jin, H.O.; Kubo, T.; Funada, R. Differences in the timing of cell death, differentiation and function among three different types of ray parenchyma cells in the hardwood Populus sieboldii×P. grandidentata. Trees 2012, 26, 743–750. [Google Scholar] [CrossRef]
- Murakami, Y.; Funada, R.; Sano, Y.; Ohtani, J. The differentiation of contact cells and isolation cells in the xylem ray parenchyma of Populus maximowiczii. Ann. Bot. 1999, 84, 429–435. [Google Scholar] [CrossRef][Green Version]
- Hillis, W.E. Wood quality and utilization. In Eucalypts for Wood Production; Hillis, W.E., Brown, A.G., Eds.; CSIRO: East Melbourne, Australia, 1978; pp. 259–289. [Google Scholar]
- Kraft, G. Zur Lehre von den Durch Forstungen; Schlagstellungen und Lichtungshieben: Hanover, Germany, 1884. [Google Scholar]
- Broda, B. Metody Histochemii Roślinnej; Państwowy Zakład Wydawnictw Lekarskich: Warszawa, Poland, 1971. [Google Scholar]
- Albinsson, B.; Li, S.; Lundquist, K.; Stomberg, R. The origin of lignin fluorescence. J. Mol. Struct. 1999, 508, 19–27. [Google Scholar] [CrossRef]
- Zheng, P.; Aoki, D.; Yoshida, M.; Matsushita, Y.; Imai, T.; Fukushima, K. Lignification of ray parenchyma cells in the xylem of Pinus densiflora. Part I: Microscopic investigation by POM, UV microscopy, and TOF-SIMS. Holzforschung 2014, 68, 897–906. [Google Scholar] [CrossRef]
- Mishra, G.; Collings, D.A.; Altaner, M.A. Cell organelles and fluorescence of parenchyma cells in Eucalyptus bosistoana sapwood and heartwood investigated by microscopy. N. Z. J. For. Sci. 2018, 48, 13. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Number of SW and HW Rings at Successive Stem Heights | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Heights (m) | 0 | 1.3 | 2.3 | 3.3 | 4.3 | 5.3 | 6.3 | 7.3 | 8.3 | 9.3 | 10.3 | 11.3 | 12.3 | 13.3 | 14.3 |
| SW1 | 42 | 36 | 33 | 33 | 30 | 30 | 28 | 27 | 25 | 23 | 21 | 21 | 21 | 18 | 16 |
| SW2 | 40 | 37 | 34 | 33 | 33 | 31 | 30 | 27 | 27 | 26 | 25 | 24 | 19 | 18 | nd |
| Mean | 41.0 | 36.5 | 33.5 | 33.0 | 31.5 | 30.3 | 29.0 | 27.0 | 26.0 | 24.5 | 23.0 | 22.5 | 20.0 | 18.0 | 16.0 |
| HW1 | 8 | 11 | 11 | 11 | 11 | 9 | 9 | 8 | 8 | 7 | 7 | 4 | 4 | 3 | 2 |
| HW2 | 10 | 12 | 12 | 12 | 8 | 8 | 7 | 6 | 5 | 5 | 3 | 2 | 2 | 0 | nd |
| Mean | 9.0 | 11.5 | 11.5 | 11.5 | 9.5 | 8.5 | 8.0 | 7.0 | 6.5 | 6.0 | 5.0 | 3.0 | 3.0 | 1.5 | 2.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tulik, M.; Jura-Morawiec, J.; Bieniasz, A.; Marciszewska, K. How Long Do Wood Parenchyma Cells Live in the Stem of a Scots Pine (Pinus sylvestris L.)? Studies on Cell Nuclei Status along the Radial and Longitudinal Stem Axes. Forests 2019, 10, 977. https://doi.org/10.3390/f10110977
Tulik M, Jura-Morawiec J, Bieniasz A, Marciszewska K. How Long Do Wood Parenchyma Cells Live in the Stem of a Scots Pine (Pinus sylvestris L.)? Studies on Cell Nuclei Status along the Radial and Longitudinal Stem Axes. Forests. 2019; 10(11):977. https://doi.org/10.3390/f10110977
Chicago/Turabian StyleTulik, Mirela, Joanna Jura-Morawiec, Anna Bieniasz, and Katarzyna Marciszewska. 2019. "How Long Do Wood Parenchyma Cells Live in the Stem of a Scots Pine (Pinus sylvestris L.)? Studies on Cell Nuclei Status along the Radial and Longitudinal Stem Axes" Forests 10, no. 11: 977. https://doi.org/10.3390/f10110977
APA StyleTulik, M., Jura-Morawiec, J., Bieniasz, A., & Marciszewska, K. (2019). How Long Do Wood Parenchyma Cells Live in the Stem of a Scots Pine (Pinus sylvestris L.)? Studies on Cell Nuclei Status along the Radial and Longitudinal Stem Axes. Forests, 10(11), 977. https://doi.org/10.3390/f10110977