Strong Disturbance Impact of Tropical Cyclone Lionrock (2016) on Korean Pine-Broadleaved Forest in the Middle Sikhote-Alin Mountain Range, Russian Far East



Abstract
1. Introduction
2. Materials and Methods
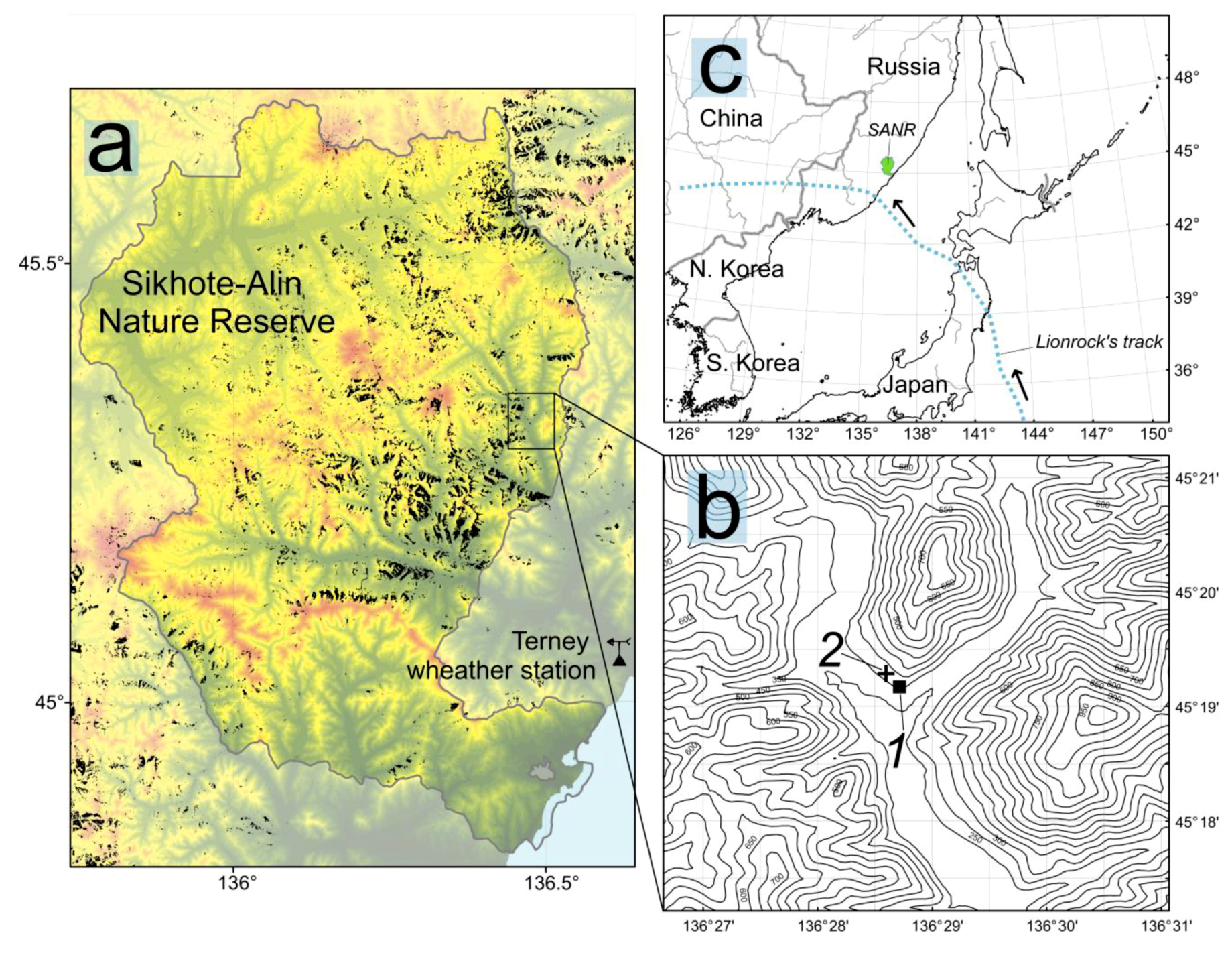
2.1. Study Area
2.2. Field Methods
2.3. Statistical Analyses
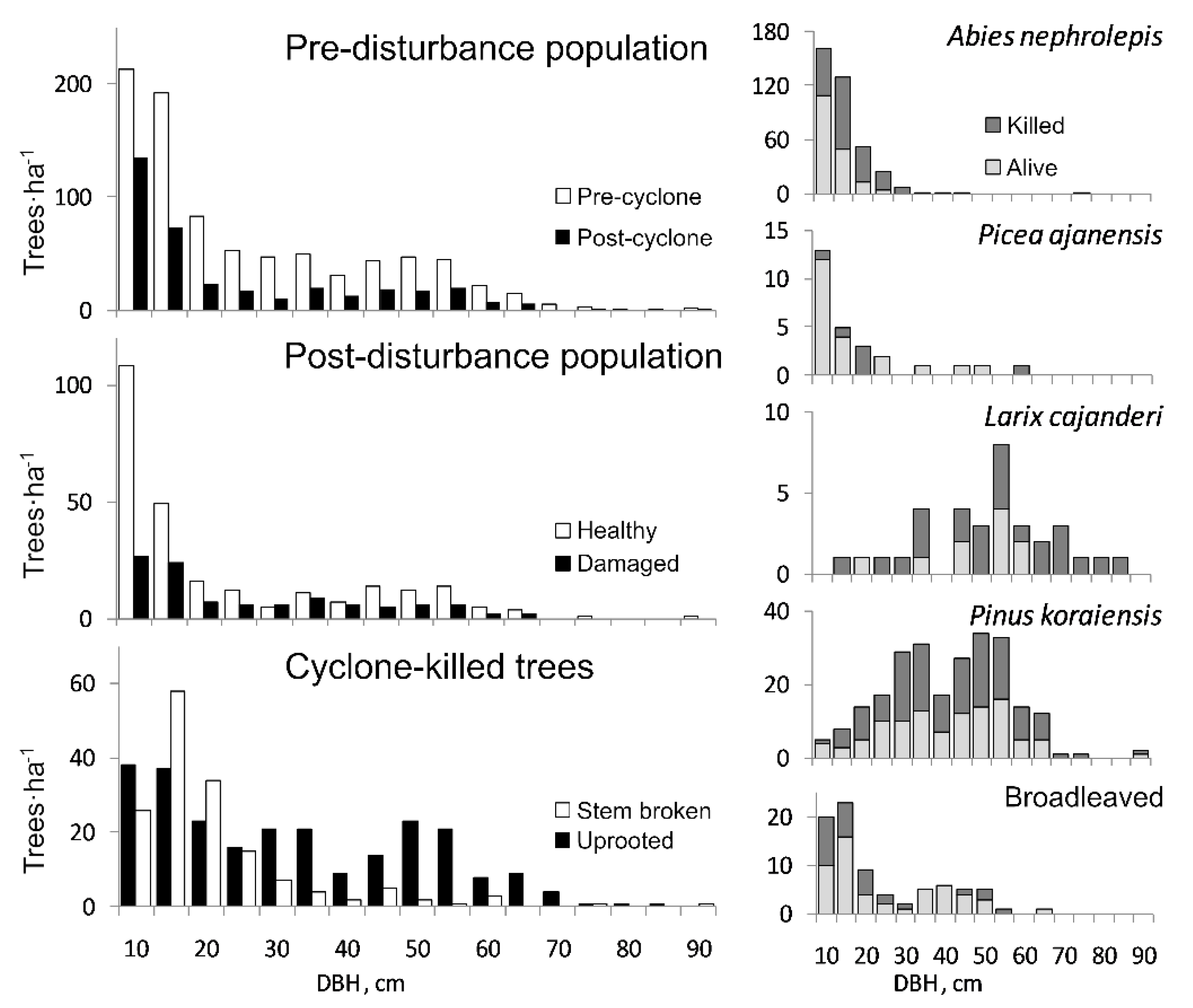
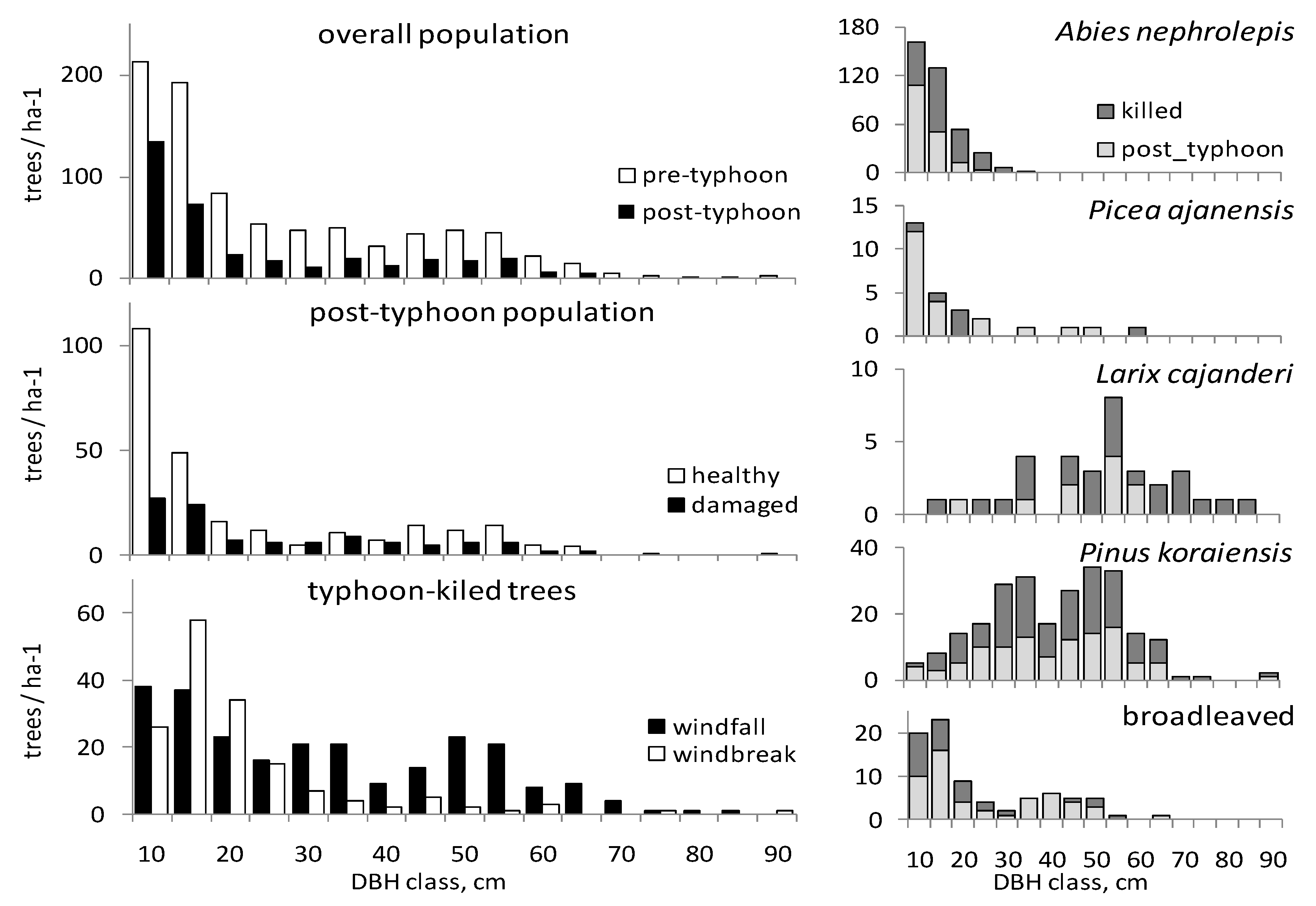
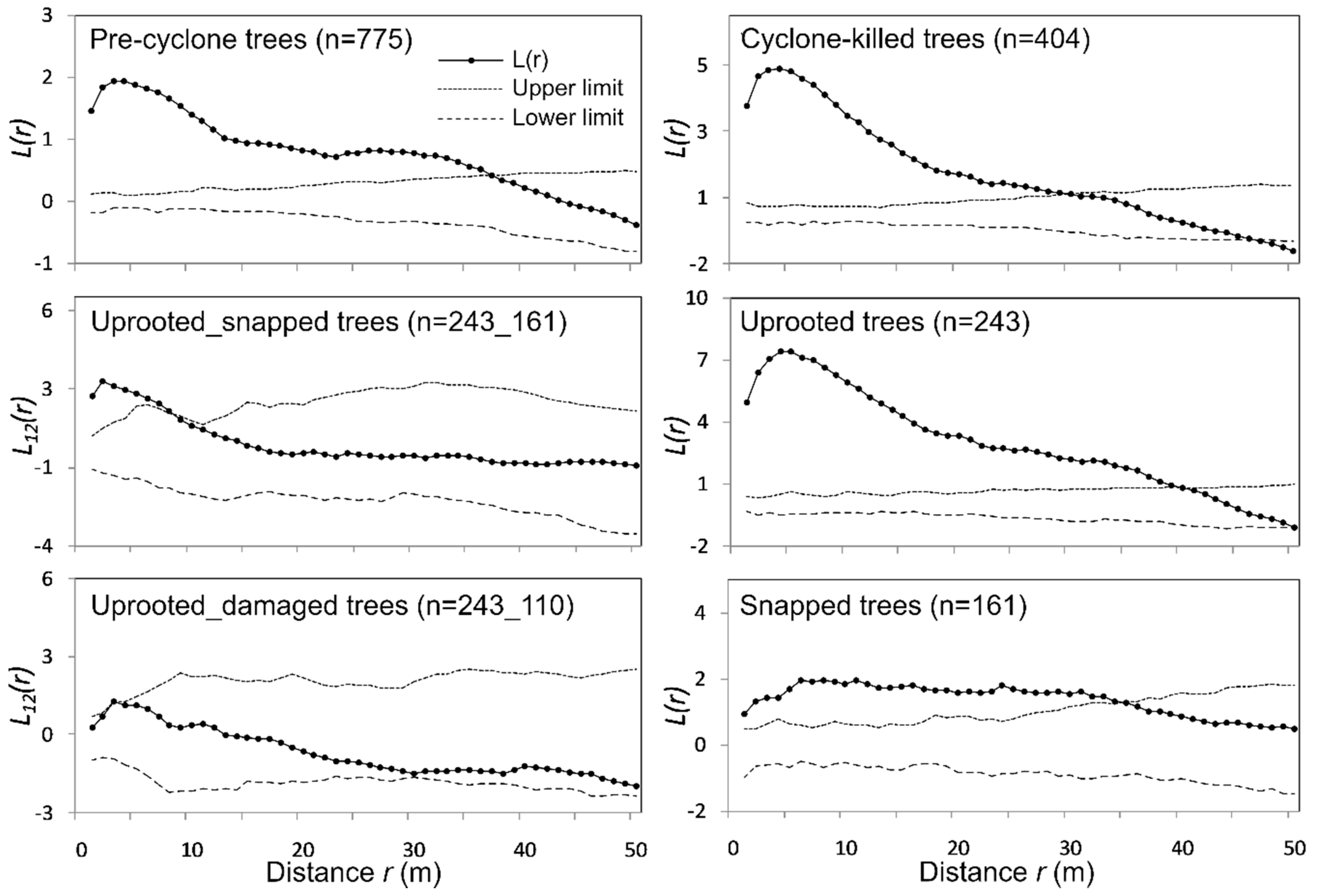
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foster, D.R.; Boose, E.R. Patterns of forest damage resulting from catastrophic wind in central New England, USA. J. Ecol. 1992, 80, 79–98. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kulakowski, D.; Eisenhart, K.S.; Baker, W.L. Subalpine forest damage from a severe windstorm in northern Colorado. Can. J. For. Res. 2001, 31, 2089–2097. [Google Scholar] [CrossRef]
- Canham, C.D.; Loucks, O.L. Catastrophic windthrow in the presettlementforests of Wisconsin. J. Ecol. 1984, 65, 803–809. [Google Scholar] [CrossRef]
- Foster, D.R. Species and stand response to catastrophic wind in central New England, USA. J. Ecol. 1988, 76, 135–151. [Google Scholar] [CrossRef]
- Peterson, C.J.; Pickett, S.T.A. Treefall and resprouting following catastrophic windthrow in an old–growth hemlock–hardwoods forest. For. Ecol. Manag. 1991, 42, 205–217. [Google Scholar] [CrossRef]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 9, 2833–2849. [Google Scholar] [CrossRef]
- Chi, C.H.; McEwan, R.W.; Chang, C.T.; Zheng, C.; Yang, Z.; Chiang, J.M.; Lin, T.C. Typhoon disturbance mediates elevational patterns of forest structure, but not species diversity, in humid monsoon Asia. Ecosystems 2015, 18, 1410–1423. [Google Scholar] [CrossRef]
- Sommerfeld, A.; Senf, C.; Buma, B.; D’Amato, A.W.; Després, T.; Díaz–Hormazábal, I.; Fraver, S.; Frelich, L.E.; Gutiérrez, Á.G.; Hart, S.J.; et al. Patterns and drivers of recent disturbances across the temperate forest biome. Nat. Commun. 2018, 9, 4355. [Google Scholar] [CrossRef]
- Everham, E.M.; Brokaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar] [CrossRef]
- Meigs, G.W.; Keeton, W.S. Intermediate-severity wind disturbance in mature temperate forests: Legacy structure, carbon storage, and stand dynamics. Ecol. Appl. 2018, 28, 798–815. [Google Scholar] [CrossRef]
- Royo, A.A.; Peterson, C.J.; Stanovick, J.S.; Carson, W.P. Evaluating the ecological impacts of salvage logging: Can natural and anthropogenic disturbances promote coexistence? Ecology 2016, 97, 1566–1582. [Google Scholar] [CrossRef]
- Bellingham, P.J. Cyclone effects on Australian rain forests: An overview. Austral. Ecol. 2008, 33, 580–584. [Google Scholar] [CrossRef]
- Tanner, E.V.J.; Rodriguez–Sanchez, F.; Healey, J.R.; Holdaway, R.J.; Bellingham, P.J. Long–term hurricane damage effects on tropical forest tree growth and mortality. Ecology 2014, 95, 2974–2983. [Google Scholar] [CrossRef]
- Lin, T.C.; Hamburg, S.P.; Lin, K.C.; Wang, L.J.; Chang, C.T.; Hsia, Y.J.; Vadeboncoeur, M.A.; McMullen, C.M.M.; Liu, C.P. Typhoon disturbance and forest dynamics: Lessons from a northwest Pacific subtropical forest. Ecosystems 2011, 14, 127–143. [Google Scholar] [CrossRef]
- Ritchie, E.A.; Elsberry, R.L. Simulations of the extratropical transition of tropical cyclones: Phasing between the upper-level trough and tropical cyclones. Mon. Weather Rev. 2007, 135, 862–876. [Google Scholar] [CrossRef]
- Dvigalo, V.N.; Melekestsev, I.V. The geological and geomorphic impact of catastrophic landslides in the Geyser Valley of Kamchatka: Aerial photogrammetry. J. Volkanol. Seismol. 2009, 3, 314–325. [Google Scholar] [CrossRef]
- Knutson, T.R.; Sirutis, J.J.; Zhao, M.; Tuleya, R.E.; Bender, M.; Vecchi, G.A.; Villarini, G.; Chavas, D. Global projections of intense tropical cyclone activity for the late twenty-first century from dynamical downscaling of CMIP5/RCP4.5 scenarios. J. Clim. 2015, 28, 7203–7224. [Google Scholar] [CrossRef]
- Altman, J.; Ukhvatkina, O.N.; Omelko, A.M.; Macek, M.; Plener, T.; Pejcha, V.; Cerny, T.; Petrik, P.; Srutek, M.; Song, J.-S.; et al. Poleward migration of the destructive effects of tropical cyclones during the 20th century. Proc. Natl. Acad. Sci. USA 2018, 115, 11543–11548. [Google Scholar] [CrossRef]
- Webster, P.J.; Holland, G.J.; Curry, J.A.; Chang, H.R. Changes in tropical cyclone number, duration and intensity in a warming environment. Science 2005, 309, 1844–1846. [Google Scholar] [CrossRef]
- Walsh, K.J.E.; McBride, J.L.; Klotzbach, P.J.; Balachandran, S.; Camargo, S.J.; Holland, G.; Knutson, T.R.; Kossin, J.P.; Lee, T.C.; Sobel, A.; et al. Tropical cyclones and climate change. Nat. Geosci. 2010, 3, 157–163. [Google Scholar] [CrossRef]
- Shuo, L.; Deqin, L.; Han, S.; Li, T.; Ming, Z. The physical mechanism and strong precipitation in Northeast China analysis during Typhoon “Lionrock” merging into extratropical cyclon. Plateau Meteorol. 2019, 38, 804–816. [Google Scholar]
- Nayak, S.; Takemi, T. Dynamical downscaling of Typhoon Lionrock (2016) for assessing the resulting hazards under global warming. J. Meteorol. Soc. Jap. 2019, 97, 69–88. [Google Scholar] [CrossRef]
- Kossin, J.P.; Emanuel, K.A.; Vecchi, G.A. The poleward migration of the location of tropical cyclone maximum intensity. Nature 2014, 509, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Schurman, J.S.; Trotsiuk, V.; Bace, R.; Cada, V.; Fraver, S.; Janda, P.; Kulakowski, D.; Labusova, J.; Mikolas, M.; Nagel, T.A.; et al. Large-scale disturbance legacies and the climate sensitivity of primary Picea abies forests. Glob. Change Biol. 2018, 24, 2169–2181. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Change 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Gromyko, M.N. Climate. In Plants, Fungi and Lichens of the Sikhote-Alin Reserve; Dalnauka: Vladivostok, Russia, 2016; pp. 14–20. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Nakamura, Y.; Krestov, P.V.; Omelko, A.M. Bioclimate and zonal vegetation in Northeast Asia: First approximation to an integrated study. Phytocoenologia 2007, 37, 443–470. [Google Scholar] [CrossRef]
- Oberle, B.; Ogle, K.; Zanne, A.E.; Wooda, C.W. When a tree falls: Controls on wood decay predict standing dead tree fall and new risks in changing forests. PLoS ONE 2018, 13, e0196712. [Google Scholar] [CrossRef]
- Walker, S.H.; Duncan, D.B. Estimation of the probability of an event as a function of several independent variables. Biometrika 1967, 54, 167–179. [Google Scholar] [CrossRef]
- Mehtätalo, L.; de-Miguel, S.; Gregoire, T.G. Modeling height-diameter curves for prediction. Can. J. For. Res. 2015, 45, 826–837. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: http://www.R-project.org/ (accessed on 6 October 2019).
- Besag, J. Contribution to the discussion of Dr. Ripley’s paper. J. R. Stat. Soc. 1977, B39, 193–195. [Google Scholar]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point Pattern Analysis in Ecology; Chapman and Hall/CRC: Boca Raton, FL, USA, 2014; p. 538. [Google Scholar]
- Gromyko, M.N. The first results of studying the catastrophic effect of typhoon Lyonrok on forest ecosystems of the Sikhote-Alin Nature Reserve. In Proceedings of the XII Far Eastern Conference of Nature Conservation Problems, Birobidzhan, Russian, 10–13 October 2017; pp. 35–37. [Google Scholar]
- Xi, W.; Peet, R.K.; Urban, D.L. Changes in forest structure, species diversity and spatial pattern following hurricane disturbance in a piedmont North Carolina forest, USA. J. Plant Ecol. 2008, 1, 43–57. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef]
- Boose, E.R.; Serrano, M.I.; Foster, D.R. Landscape and regional impacts of hurricanes in Puerto Rico. Ecol. Monogr. 2004, 74, 335–352. [Google Scholar] [CrossRef]
- Lin, S.Y.; Shaner, P.J.L.; Lin, T.C. Characteristics of old–growth and secondary forests in relation to age and typhoon disturbance. Ecosystems 2018, 21, 1521–1532. [Google Scholar] [CrossRef]
- Peterson, C.J. Within-stand variation in windthrow in southern boreal forests of Minnesota: Is it predictable? Can. J. For. Res. 2004, 34, 365–375. [Google Scholar] [CrossRef]
- Tan, F.; Lim, H.S.; Abdullah, K. The effects of orography in Indochina on wind, cloud, and rainfall patterns during Typhoon Ketsana (2009). Asia Pac. J. Atmos. Sci. 2012, 48, 295–314. [Google Scholar] [CrossRef]
- Sheffield, R.M.; Thompson, M.T. Hurricane Hugo: Effects on South Carolina’s Forest Resource; USDA Forest Service: Asheville, NC, USA, 1992; p. 51.
- Szwagrzyk, J.; Gazda, A.; Dobrowolska, D.; Chećko, E.; Zaremba, J.; Tomski, A. Tree mortality after wind disturbance differs among tree species more than among habitat types in a lowland forest in northeastern Poland. For. Ecol. Manag. 2017, 398, 174–184. [Google Scholar] [CrossRef]
- Walker, L.R. Tree damage and recovery from Hurricane Hugo in Luquillo Experimental Forest, Puerto Rico. Biotropica 1991, 23, 379–385. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Kapos, V.; Varty, N.; Healey, J.R.; Tanner, E.V.J.; Kelly, D.L.; Dalling, J.W.; Burns, L.S.; Lee, D.; Sidrak, G. Hurricanes need not cause high mortality: The effects of Hurricane Gilbert on forests in Jamaica. J. Trop. Ecol. 1992, 8, 217–223. [Google Scholar] [CrossRef]
- Bellingham, P.J. Landforms influence patterns of hurricane damage: Evidence from Jamaican montane forests. Biotropica 1991, 23, 427–433. [Google Scholar] [CrossRef]
- Whigham, D.F.; Dickinson, M.B.; Brokaw, N.V. Background canopy gap and catastrophic wind disturbances in tropical forests. In Ecosystems of Disturbed Ground; Elsevier Science: Amsterdam, The Netherlands, 1999; pp. 223–252. [Google Scholar]
- Woods, K.D. Intermediate disturbance in a late-successional hemlock northern hardwood forest. J. Ecol. 2004, 92, 464–476. [Google Scholar] [CrossRef]
- Jactel, H.; Bauhus, J.; Boberg, J.; Bonal, D.; Castagneyrol, B.; Gardiner, B.; Gonzalez-Olabarria, J.R.; Koricheva, J.; Meurisse, N.; Brockerhoff, E.G. Tree diversity drives forest stand resistance to natural disturbances. Curr. For. Rep. 2017, 3, 223–243. [Google Scholar] [CrossRef]
- Canham, C.D.; Papaik, M.J.; Latty, E.F. Interspecific variation in susceptibility to windthrow as a function of tree size and storm severity for northern temperate tree species. Can. J. For. Res. 2001, 31, 1–10. [Google Scholar] [CrossRef]
- Imbert, D.; Labbe, P.; Rousteau, A. Hurricane damage and forest structure in Guadeloupe, French West Indies. J. Trop. Ecol. 1996, 12, 663–680. [Google Scholar] [CrossRef]
- Greenberg, C.H.; McNab, W.H. Forest disturbance in hurricane-related downbursts in the Appalachian Mountains of North Carolina. For. Ecol. Manag. 1998, 104, 179–191. [Google Scholar] [CrossRef]
- Basnet, K.; Likens, G.E.; Scatera, F.N.; Lugo, A.E. Hurricane Hugo: Damage to a tropical rain forest in Puerto Rico. J. Trop. Ecol. 1992, 8, 47–55. [Google Scholar] [CrossRef]
- Tanner, E.V.J.; Kapos, V.; Healey, J.R. Hurricane effects on forest ecosystems in the Caribbean. Biotropica 1991, 23, 513–521. [Google Scholar] [CrossRef]
- Nagel, T.A.; Diaci, J. Intermediate wind disturbance in an old-growth beech–fir forest in southeastern Slovenia. Can. J. For. Res. 2006, 36, 629–638. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Density, Stem·(ha−1) | Density, % of Overall | Basal Area, m2·ha−1 | Basal Area, % of Overall | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Overall | Damaged | Cylone-killed | Damaged | Cylone-killed | Overall | Damaged | Cylone-killed | Damaged | Cyclone-killed | |
| Betula costata | 16 | 4 | 11 | 0.5 | 1.4 | 0.23 | 0.03 | 0.15 | 0.1 | 0.3 |
| Pice aajanensis | 28 | 6 | 7 | 0.8 | 0.9 | 0.91 | 0.15 | 0.34 | 0.3 | 0.7 |
| Pinus koraiensis | 246 | 33 | 140 | 4.3 | 18.2 | 33.84 | 4.54 | 19.63 | 8.8 | 37.9 |
| Quercus mongolica | 15 | 7 | 4 | 0.9 | 0.5 | 2.06 | 1.04 | 0.61 | 2.0 | 1.2 |
| Tilia amurensis | 26 | 8 | 7 | 1.0 | 0.9 | 1.43 | 0.47 | 0.26 | 0.9 | 0.5 |
| Larix cajanderi | 34 | 0 | 24 | 0.0 | 3.1 | 7.15 | 0.00 | 5.38 | 0.0 | 10.4 |
| Abies nephrolepis | 381 | 38 | 204 | 4.9 | 26.6 | 5.92 | 0.41 | 3.92 | 0.8 | 7.6 |
| Acer mono | 6 | 1 | 4 | 0.1 | 0.5 | 0.02 | 0.00 | 0.01 | 0.0 | 0.0 |
| Acer ukurunduense | 9 | 4 | 1 | 0.5 | 0.1 | 0.06 | 0.04 | 0.00 | 0.1 | 0.0 |
| Betula platyphylla | 1 | 1 | 0 | 0.1 | 0.0 | 0.10 | 0.10 | 0.00 | 0.2 | 0.0 |
| Prunus maximowiczii | 5 | 3 | 2 | 0.4 | 0.3 | 0.07 | 0.05 | 0.02 | 0.1 | 0.0 |
| Sorbus amurensis | 1 | 1 | 0 | 0.1 | 0.0 | 0.01 | 0.01 | 0.00 | 0.0 | 0.0 |
| Total | 768 | 106 | 404 | 13.8 | 52.6 | 51.80 | 6.85 | 30.32 | 13.2 | 58.5 |
| Size Class (cm) | Snapped Trees | Uprooted Trees | Total Killed Trees | ||||||
|---|---|---|---|---|---|---|---|---|---|
| No | DBH | P ± Δ | No | DBH | P ± Δ | % | DBH | P ± Δ | |
| 10–30 | 133 | 13.9 ± 4.9 | 0.17 ± 0.03 | 114 | 13.3 ± 5.5 | 0.15 ± 0.02 | 32.0 | 13.6 ± 5.2 | 0.32 ± 0.03 |
| 30–60 | 21 | 35.8 ± 7.8 | 0.03 ± 0.01 | 109 | 40.7 ± 9.2 | 0.14 ± 0.02 | 16.9 | 39.9 ± 9.1 | 0.17 ± 0.03 |
| >60 | 5 | 66.9 ± 13.5 | 0.006 ± 0.005 | 24 | 63.3 ± 6.8 | 0.03 ± 0.01 | 3.8 | 63.9 ± 8.2 | 0.38 ± 0.03 |
| Total | 159 | 18.3 ± 12.7 | 0.21 ± 0.03 | 247 | 30.2 ± 18.6 | 0.32± 0.03 | 52.7 | 25.5 ± 17.5 | 0.53 ± 0.03 |
| Year | Density | Basal Area | ||
|---|---|---|---|---|
| Stem·(ha−1) | Percent·10 year−1 | m2·ha−1 | Percent·10 year−1 | |
| 1967–1976 | 46 | 3.6 | 1.2 | 2.4 |
| 1977–1986 | 88 | 6.9 | 2.4 | 5.0 |
| 1987–1996 | 88 | 6.9 | 2.4 | 5.0 |
| 1997–2006 | 156 | 13.7 | 3.4 | 7.2 |
| 2007–2016 | 128 | 11.2 | 3.2 | 6.9 |
| 2017dead trees | 88 | 7.7 | 3.0 | 6.4 |
| 2017_damaged trees | 20 | 1.8 | 1.4 | 2.8 |
| Mean (1967–2016) | 101 ± 42 | 8.5 ± 4.0 | 2.5 ± 0.9 | 5.3 ± 2.0 |
| Total (1967–2016) | 506 | 42.4 | 12.4 | 26.5 |
| Tree Number | Proportion (%) | |||
|---|---|---|---|---|
| Healthy | Hollow | Healthy | Hollow | |
| Larixcajanderi | ||||
| alive | 5 | 3 | 62.5 | 37.5 |
| snapped | 1 | 0 | 100.0 | 0.0 |
| uprooted | 6 | 1 | 85.7 | 14.3 |
| Pinuskoraiensis | ||||
| alive | 40 | 21 | 65.6 | 34.4 |
| snapped | 4 | 1 | 80.0 | 20.0 |
| uprooted | 46 | 19 | 70.8 | 29.2 |
| Total | ||||
| alive | 45 | 24 | 65.2 | 34.8 |
| snapped | 5 | 1 | 83.3 | 16.7 |
| uprooted | 52 | 20 | 72.2 | 27.8 |
| Logistic Regression Coefficients | Estimate | ||||||
|---|---|---|---|---|---|---|---|
| Species | Intercept | DBH | P | H | W | LOOCV | Killed Trees (%) |
| All trees case | |||||||
| Pinus koraiensis | 6.29*** | 0.17*** | −599.0* | −0.51*** | - | 0.85 | 0.7 |
| Abies nephrolepis | 0.37 | 0.21*** | −998.3*** | −0.12 | - | 0.66 | 0.63 |
| Larix cajanderi | 0.56 | 0.006 | 1595* | −0.064 | - | 0.54 | 0.71 |
| Broadleaved species | 4.24*** | 0.12*** | −1676* | −0.36 | - | 0.77 | 0.71 |
| Total | 0.29*** | 0.12*** | −610.0*** | −0.26*** | - | 0.71 | 0.66 |
| Hollow presence trees case | |||||||
| Pinus koraiensis | 6.82*** | 0.22*** | −581 | −0.61*** | −0.27 | 0.86 | 0.66 |
| Larix cajanderi | −8 | −0.037 | 533 | 0.33 | −1.9 | 0.31 | 0.5 |
| Total | 5.28*** | 0.19*** | −648 | −0.49*** | −0.13 | 0.8 | 0.64 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vozmishcheva, A.S.; Bondarchuk, S.N.; Gromyko, M.N.; Kislov, D.E.; Pimenova, E.A.; Salo, M.A.; Korznikov, K.A. Strong Disturbance Impact of Tropical Cyclone Lionrock (2016) on Korean Pine-Broadleaved Forest in the Middle Sikhote-Alin Mountain Range, Russian Far East. Forests 2019, 10, 1017. https://doi.org/10.3390/f10111017
Vozmishcheva AS, Bondarchuk SN, Gromyko MN, Kislov DE, Pimenova EA, Salo MA, Korznikov KA. Strong Disturbance Impact of Tropical Cyclone Lionrock (2016) on Korean Pine-Broadleaved Forest in the Middle Sikhote-Alin Mountain Range, Russian Far East. Forests. 2019; 10(11):1017. https://doi.org/10.3390/f10111017
Chicago/Turabian StyleVozmishcheva, Anna S., Svetlana N. Bondarchuk, Mikhail N. Gromyko, Dmitriy E. Kislov, Elena A. Pimenova, Michail A. Salo, and Kirill A. Korznikov. 2019. "Strong Disturbance Impact of Tropical Cyclone Lionrock (2016) on Korean Pine-Broadleaved Forest in the Middle Sikhote-Alin Mountain Range, Russian Far East" Forests 10, no. 11: 1017. https://doi.org/10.3390/f10111017
APA StyleVozmishcheva, A. S., Bondarchuk, S. N., Gromyko, M. N., Kislov, D. E., Pimenova, E. A., Salo, M. A., & Korznikov, K. A. (2019). Strong Disturbance Impact of Tropical Cyclone Lionrock (2016) on Korean Pine-Broadleaved Forest in the Middle Sikhote-Alin Mountain Range, Russian Far East. Forests, 10(11), 1017. https://doi.org/10.3390/f10111017