Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling and Measuring Methods
2.3. Calculations and Statistical Analyses
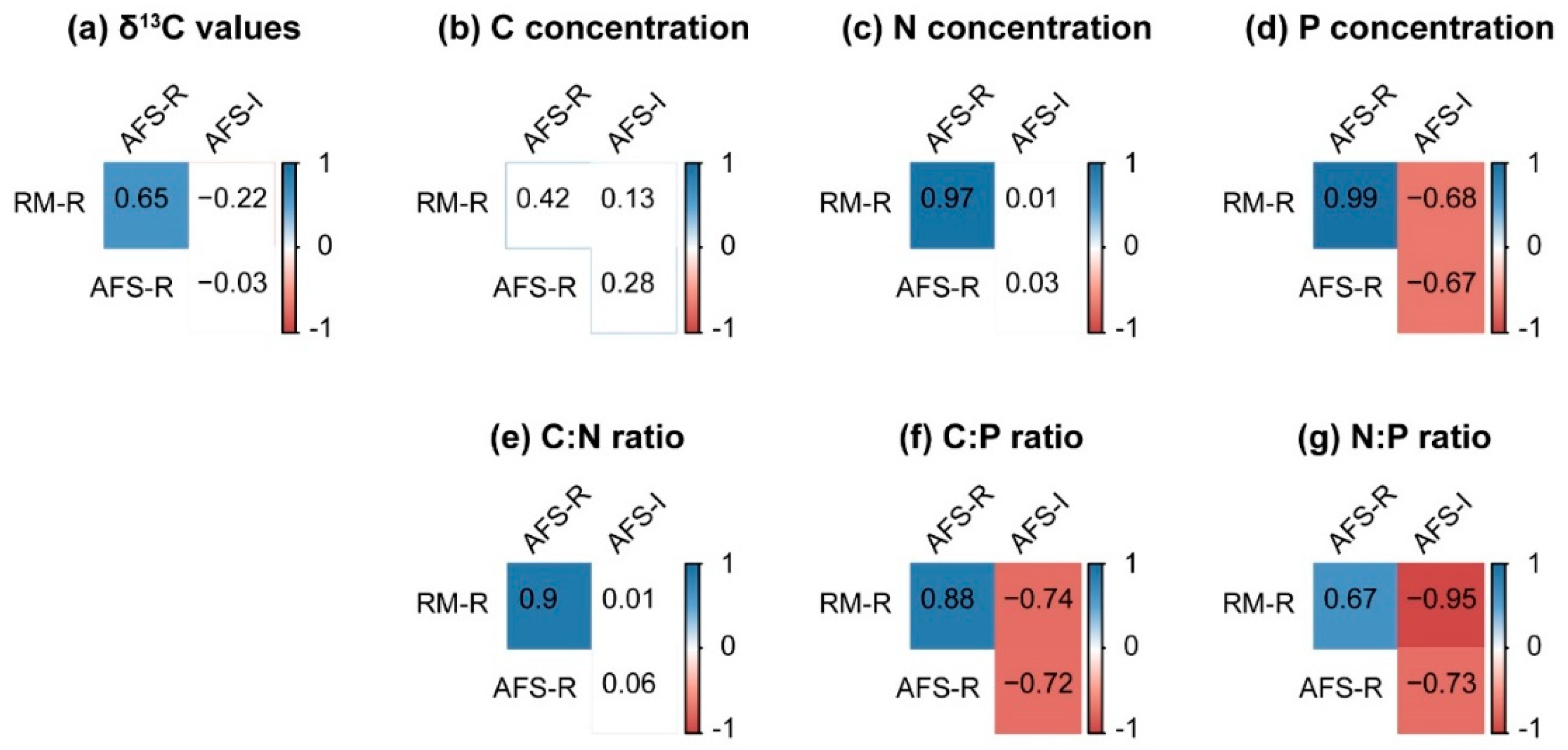
3. Results
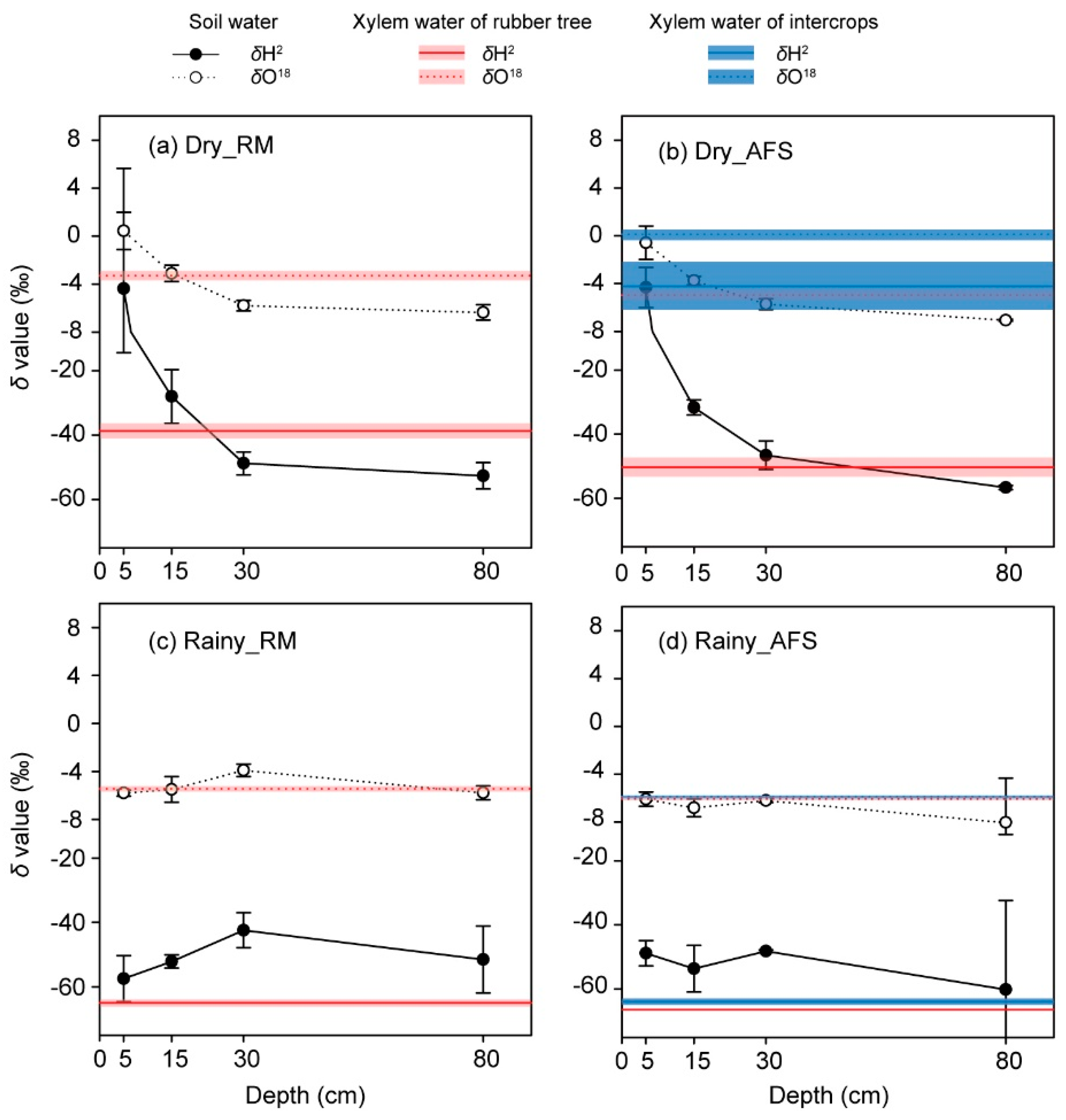
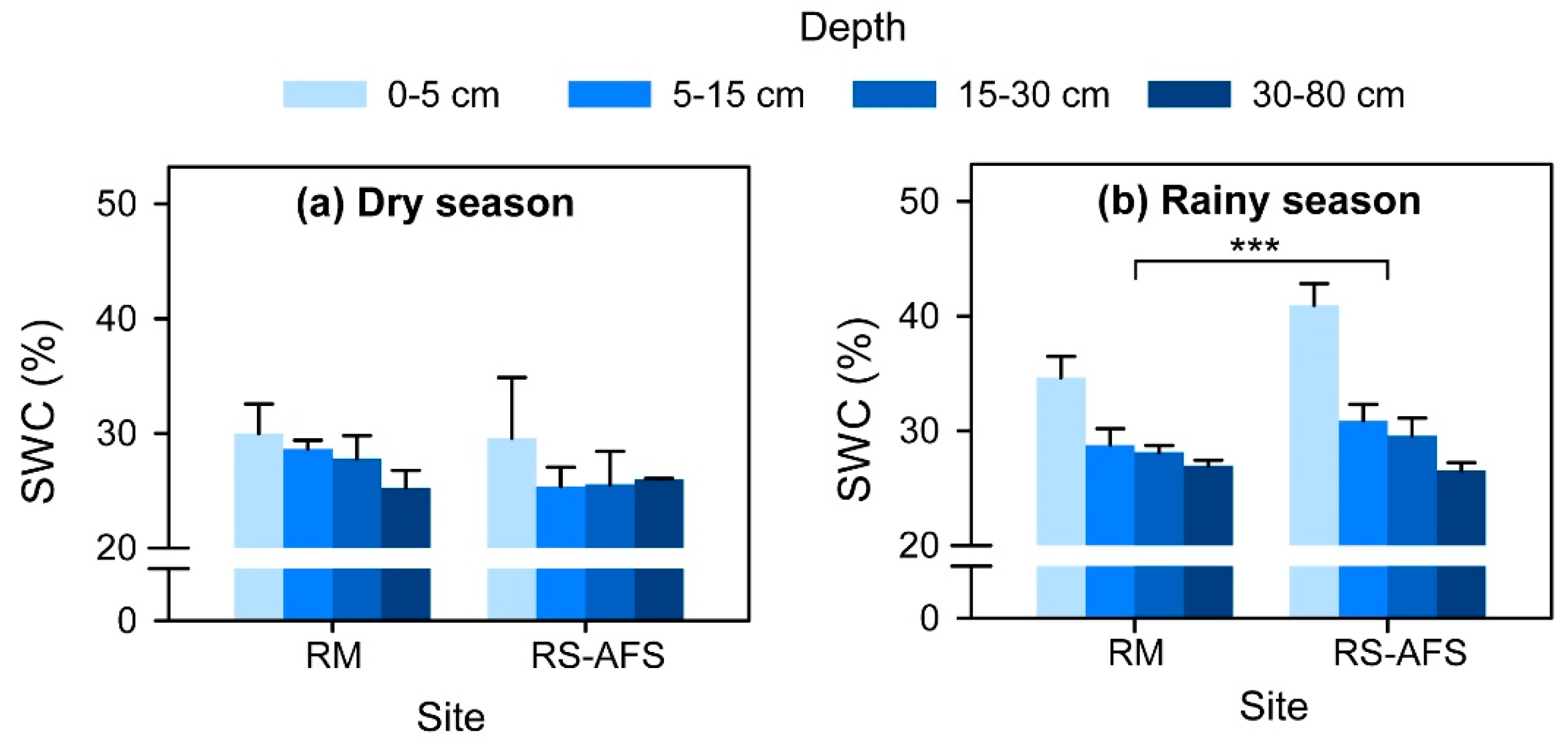
3.1. Plant Xylem Water and Soil Water
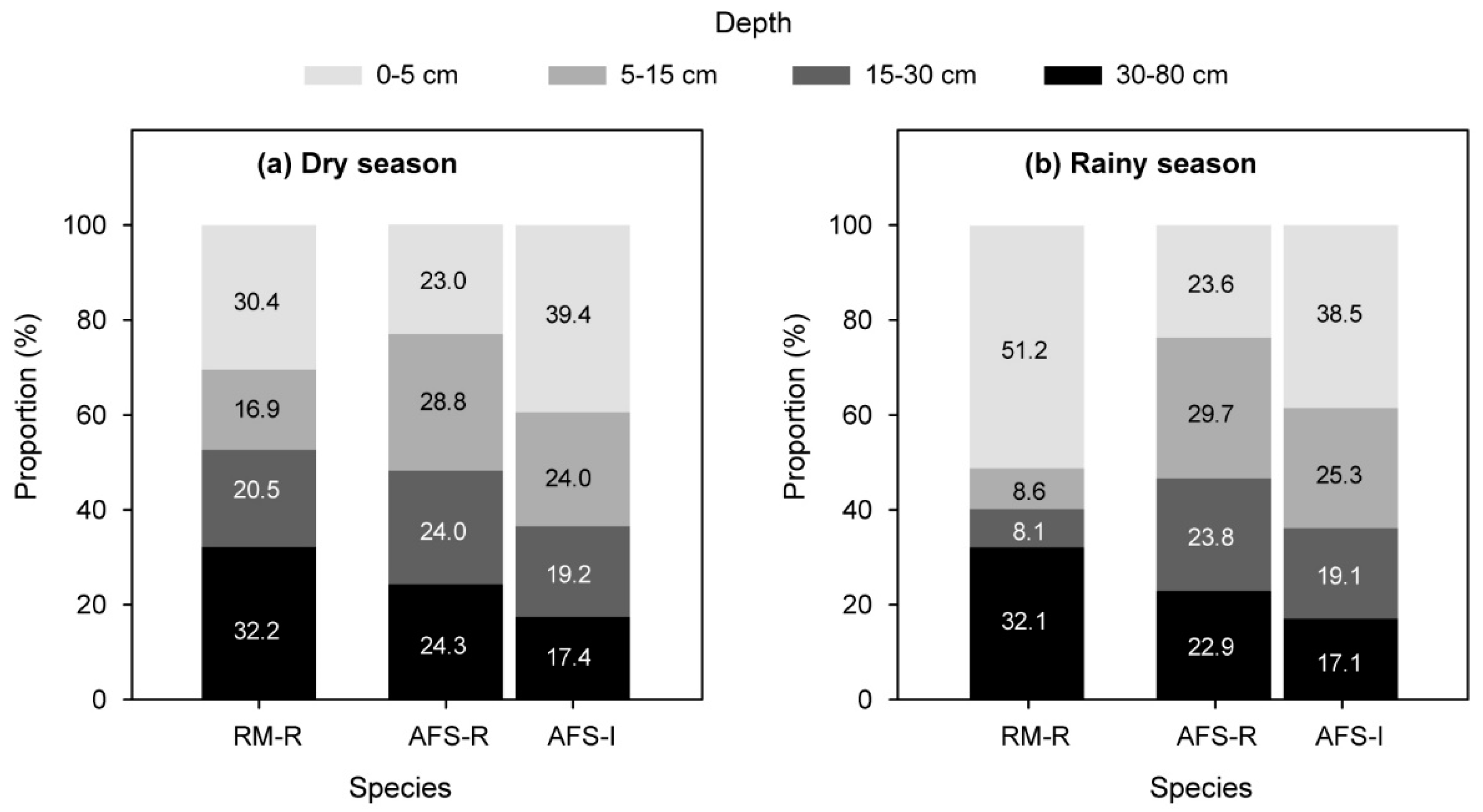
3.2. Plant Water Sources
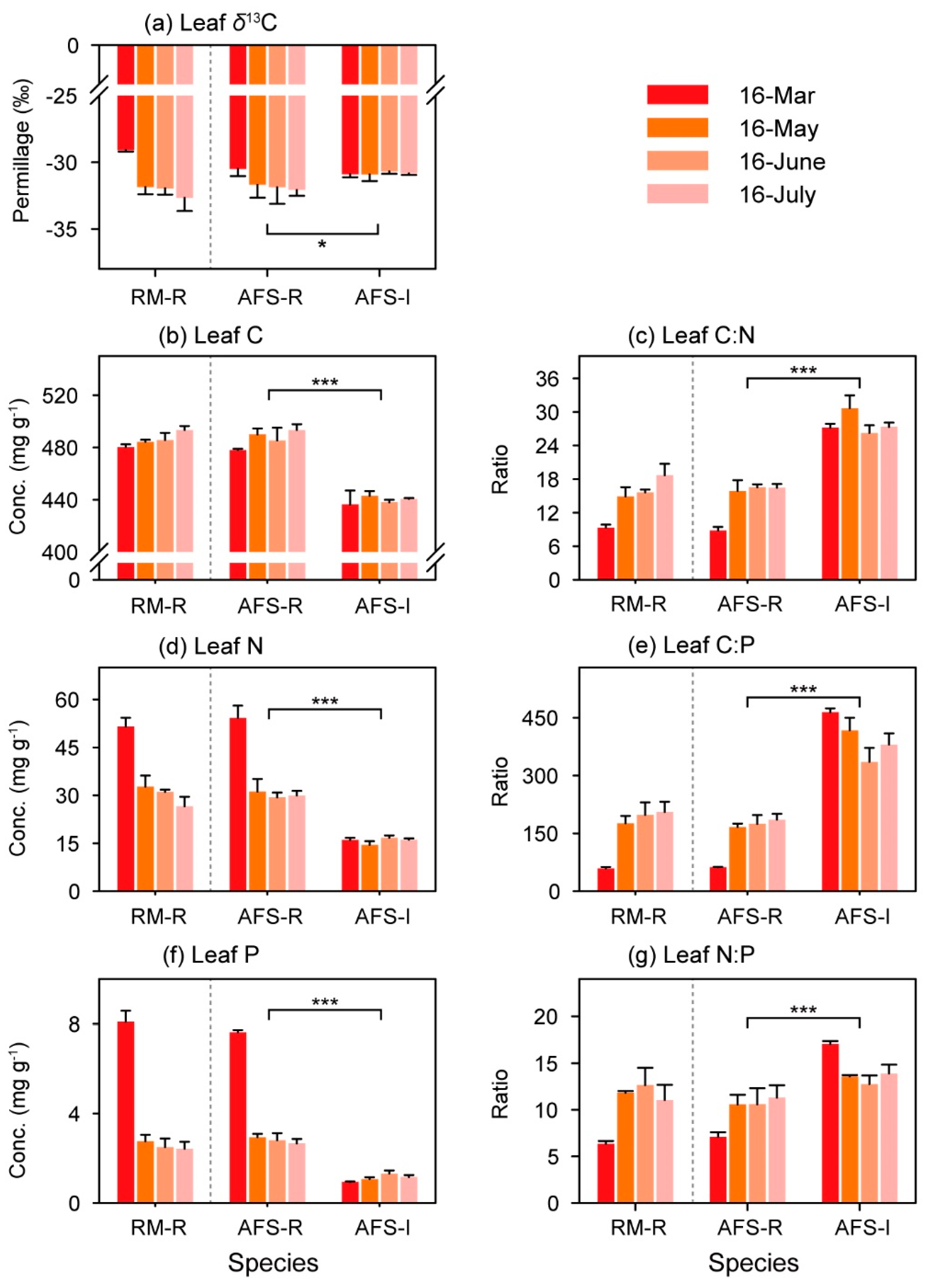
3.3. Leaf δ13C, Nutrient Concentrations, and Ratios
4. Discussion
4.1. Plant Water-Absorbing Patterns
4.2. Plant Water Use Efficiency and Soil Water Conditions
4.3. Plant Leaf Carbon, Nitrogen, and Phosphorus Concentrations and Ratios
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cornish, K. Alternative natural rubber crops: Why should we care? Technol. Innov. 2017, 18, 244–255. [Google Scholar] [CrossRef]
- Mann, C.C. Addicted to rubber. Science 2009, 325, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Castella, J.C. Expansion of rubber (Hevea brasiliensis) in Mainland Southeast Asia: What are the prospects for smallholders? J. Peasant Stud. 2013, 40, 155–170. [Google Scholar] [CrossRef]
- Wigboldus, S.; Hammond, J.; Xu, J.; Yi, Z.F.; He, J.; Klerkx, L.; Leeuwis, C. Scaling green rubber cultivation in Southwest China–An integrative analysis of stakeholder perspectives. Sci. Total Environ. 2017, 580, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kono, Y.; Kobayashi, S. The process of expansion in commercial banana cropping in tropical China: A case study at a Dai village, Mengla County. Agric. Syst. 2014, 124, 32–38. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Corlett, R.T.; Zhai, D. After the rubber boom: Good news and bad news for biodiversity in Xishuangbanna, Yunnan, China. Reg. Environ. Chang. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Li, H.; Ma, Y.; Aide, T.M.; Liu, W. Past, present and future land-use in Xishuangbanna, China and the implications for carbon dynamics. For. Ecol. Manag. 2008, 255, 16–24. [Google Scholar] [CrossRef]
- Mann, C.C. Why We (Still) Can’t Live Without Rubber. Available online: https://www.nationalgeographic.com/magazine/2016/01/southeast-asia-rubber-boom/ (accessed on 20 September 2019).
- Song, Q.H.; Tan, Z.H.; Zhang, Y.P.; Sha, L.Q.; Deng, X.B.; Deng, Y.; Zhao, W. Do the rubber plantations in tropical China act as large carbon sinks? iForest 2013, 7, 42–47. [Google Scholar] [CrossRef]
- Blagodatsky, S.; Xu, J.; Cadisch, G. Carbon balance of rubber (Hevea brasiliensis) plantations: A review of uncertainties at plot, landscape and production level. Agric. Ecosyst. Environ. 2016, 221, 8–19. [Google Scholar] [CrossRef]
- Zhang, M.; Chang, C.; Quan, R. Natural forest at landscape scale is most important for bird conservation in rubber plantation. Biol. Conserv. 2017, 210, 243–252. [Google Scholar] [CrossRef]
- Langenberger, G.; Cadisch, G.; Martin, K.; Min, S.; Waibel, H. Rubber intercropping: A viable concept for the 21st century? Agrofor. Syst. 2017, 91, 577–596. [Google Scholar] [CrossRef]
- John, J.; Nair, A.M. Prospects of allelopathic research in multi-storey cropping systems. In Allelopathy in Ecological Agriculture and Forestry; Narwal, S.S., Hoagland, R.E., Dilday, R.H., Reigosa Roger, M.J., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 159–179. [Google Scholar]
- He, Z.H.; Ge, W.; Yue, G.G.L.; Bik-San Lau, C.; He, M.F.; But, P.P.H. Anti-angiogenic effects of the fruit of Alpinia oxyphylla. J. Ethnopharmacol. 2010, 132, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Liu, Q.F.; Yan, J.K.; Zhang, T.L.; Wang, Q.Y. Seasonal changes of photosynthetic characteristics of Alpinia oxyphylla growing under Hevea brasiliensis. Chin. J. Plant Ecol. 2018, 42, 585–594. [Google Scholar]
- FAO and IAEA. Management of Agroforestry Systems for Enhancing Resource use Efficiency and Crop Productivity. Available online: https://www.iaea.org/publications/8181/management-of-agroforestry-systems-for-enhancing-resource-use-efficiency-and-crop-productivity (accessed on 29 September 2019).
- Craine, J.M.; Dybzinski, R. Mechanisms of plant competition for nutrients, water and light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef]
- Fry, B. Stable Isotope Ecology; Springer: New York, NY, USA, 2006. [Google Scholar]
- White, J.W.; Cook, E.R.; Lawrence, J.R. The DH ratios of sap in trees: Implications for water sources and tree ring DH ratios. Geochim. Cosmochim. Acta 1985, 49, 237–246. [Google Scholar] [CrossRef]
- Silvertown, J.; Araya, Y.; Gowing, D. Hydrological niches in terrestrial plant communities: A review. J. Ecol. 2015, 103, 93–108. [Google Scholar] [CrossRef]
- Youri, R.; Mathieu, J. Reviews and syntheses: Isotopic approaches to quantify root water uptake: A review and comparison of methods. Biogeosciences 2017, 14, 2199–2224. [Google Scholar]
- Chapin, F.S., III; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer: New York, NY, USA, 2011. [Google Scholar]
- Everard, K.; Seabloom, E.W.; Harpole, W.S.; de Mazancourt, C. Plant water use affects competition for nitrogen: Why drought favors invasive species in California. Am. Nat. 2009, 1, 85–97. [Google Scholar] [CrossRef]
- Guardiola-Claramonte, M.; Troch, P.A.; Ziegler, A.D.; Giambelluca, T.W.; Durcik, M.; Vogler, J.B.; Nullet, M.A. Hydrologic effects of the expansion of rubber (Hevea brasiliensis) in a tropical catchment. Ecohydrol. 2010, 3, 306–314. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Chen, C. Can intercropping with the world’s three major beverage plants help improve the water use of rubber trees? J. Appl. Ecol. 2016, 53, 1787–1799. [Google Scholar] [CrossRef]
- Parnell, A.C.; Phillips, D.L.; Bearhop, S.; Semmens, B.X.; Ward, E.J.; Moore, J.W.; Inger, R. Bayesian stable isotope mixing models. Environmetrics 2013, 24, 387–399. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing Computer Software; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- George, S.; Suresh, P.R.; Wahid, P.A.; Nair, R.B.; Punnoose, K.I. Active root distribution pattern of Hevea brasiliensis determined by radioassay of latex serum. Agrofor. Syst. 2009, 76, 275–281. [Google Scholar] [CrossRef]
- Priyadarshan, P. Biology of Hevea Rubber; CABI: Wallingford, UK, 2011. [Google Scholar]
- Kobayashi, N.; Kumagai, T.O.; Miyazawa, Y.; Matsumoto, K.; Tateishi, M.; Lim, T.K.; Yin, S. Transpiration characteristics of a rubber plantation in central Cambodia. Tree Physiol. 2014, 34, 285–301. [Google Scholar] [CrossRef]
- Giambelluca, T.W.; Mudd, R.G.; Liu, W.; Ziegler, A.D.; Kobayashi, N.; Kumagai, T.O.; Yin, S. Evapotranspiration of rubber (Hevea brasiliensis) cultivated at two plantation sites in Southeast Asia. Water Resour. Res. 2016, 52, 660–679. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Chen, C. How do plants share water sources in a rubber-tea agroforestry system during the pronounced dry season. Agric. Ecosyst. Environ. 2017, 236, 69–77. [Google Scholar] [CrossRef]
- Bleby, T.M.; Mcelrone, A.J.; Jackson, R.B. Water uptake and hydraulic redistribution across large woody root systems to 20 m depth. Plant Cell Environ. 2010, 33, 2132–2148. [Google Scholar] [CrossRef]
- Callaway, R.M. The detection of neighbors by plants. Trends Ecol. Evol. 2002, 17, 104–105. [Google Scholar] [CrossRef]
- Jiang, X.J.; Liu, W.; Wu, J.; Wang, P.; Liu, C.; Yuan, Z.Q. Land Degradation Controlled and Mitigated by Rubber-based Agroforestry Systems through Optimizing Soil Physical Conditions and Water Supply Mechanisms: A Case Study in Xishuangbanna, China. Land Degrad. Dev. 2017, 28, 2277–2289. [Google Scholar] [CrossRef]
- Pierret, A.; Maeght, J.L.; Clément, C.; Montoroi, J.P.; Hartmann, C.; Gonkhamdee, S. Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef]
- Liu, C.; Jin, Y.; Liu, C.; Tang, J.; Wang, Q.; Xu, M. Phosphorous fractions in soils of rubber-based agroforestry systems: Influence of season, management and stand age. Sci. Total Environ. 2018, 616, 1576–1588. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, M.H. Carbon isotopes in photosynthesis. BioScience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Kohn, M.J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo) ecology and (paleo) climate. Proc. Natl. Acad. Sci. USA 2010, 107, 19691–19695. [Google Scholar] [CrossRef] [PubMed]
- Ehleringer, J.R.; Roden, J.; Dawson, T.E. Assessing ecosystem-level water relations through stable isotope ratio analyses. In Methods in Ecosystem Science; Sala, O.E., Jackson, R.B., Mooney, H.A., Howarth, R.W., Eds.; Springer: New York, NY, USA, 2000; pp. 181–198. [Google Scholar]
- Marshall, J.D.; Brooks, J.R.; Lajtha, K. Sources of variation in the stable isotopic composition of plants. In Stable Isotopes in Ecology and Environmental Science; Michener, R., Lajtha, K., Eds.; John Wiley Sons: Chichester, UK, 2007; pp. 22–60. [Google Scholar]
- Sardans, J.; Rivas-Ubach, A.; Penuelas, J. The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives. Biogeochemistry 2012, 111, 1–39. [Google Scholar] [CrossRef]
- Sasaki, T.; Yoshihara, Y.; Jamsran, U.; Ohkuro, T. Ecological stoichiometry explains larger-scale facilitation processes by shrubs on species coexistence among understory plants. Ecol. Eng. 2010, 36, 1070–1075. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Oxford, UK, 2002. [Google Scholar]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.A.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley Sons: New York, NY, USA, 1995. [Google Scholar]
- Craine, J.M.; Fargione, J.; Sugita, S. Supply pre-emption, not concentration reduction, is the mechanism of competition for nutrients. New Phytol. 2005, 166, 933–940. [Google Scholar] [CrossRef]
- Craine, J.M. Competition for nutrients and optimal root allocation. Plant Soil. 2006, 285, 171–185. [Google Scholar] [CrossRef]
- Keddy, P.; Fraser, L.H.; Wisheu, I.C. A comparative approach to examine competitive response of 48 wetland plant species. J. Veg. Sci. 1998, 9, 777–786. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Effects | δ2H | δ18O | SWC | |||||
|---|---|---|---|---|---|---|---|---|
| d.f. | F | P | F | P | F | P | ||
| (A) | Season | 1 | 47.66 | *** | 35.02 | *** | 35.66 | *** |
| Site | 1 | 0.39 | 9.5 | ** | 0.78 | |||
| Depth | 3 | 18.69 | *** | 21.29 | *** | 31.05 | *** | |
| Season * Site | 1 | 0.01 | 2.02 | 9.48 | ** | |||
| Season * Depth | 3 | 21.22 | *** | 18.52 | *** | 6.73 | ** | |
| Site * Depth | 3 | 0.74 | 0.25 | 1.92 | ||||
| Season * Site * Depth | 3 | 0.52 | 0.95 | 2.11 | ||||
| (B) | Season | 1 | 37.21 | *** | 21.43 | *** | 7.19 | * |
| Depth | 3 | 9.85 | *** | 17.94 | *** | 16.55 | *** | |
| Season * Depth | 3 | 22.71 | *** | 29.47 | *** | 2.7 | ||
| (C) | Season | 1 | 17.79 | *** | 17.62 | *** | 28.88 | *** |
| Depth | 3 | 9.66 | *** | 8.56 | ** | 16.46 | *** | |
| Season * Depth | 3 | 5.52 | ** | 3.64 | * | 5.12 | * | |
| (D) | Site | 1 | 0.38 | 2.68 | 1.54 | |||
| Depth | 3 | 106 | *** | 72.3 | *** | 2.85 | ||
| Site * Depth | 3 | 0.47 | 0.45 | 0.74 | ||||
| (E) | Site | 1 | 0.16 | 6.83 | * | 18.13 | *** | |
| Depth | 3 | 0.95 | 1.7 | 76.91 | *** | |||
| Site * Depth | 3 | 0.66 | 0.66 | 6.64 | ** | |||
| Tested Effects | δ13C | C | N | P | C/N | C/P | N/P | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| d.f. | F | P | F | P | F | P | F | P | F | P | F | P | F | P | ||
| (A) | Time | 3 | 19.86 | *** | 7.66 | ** | 50.44 | *** | 162.91 | *** | 24.62 | *** | 25.82 | *** | 15.21 | ** |
| (B) | Time | 3 | 2.01 | 3.82 | 50.58 | *** | 400.68 | *** | 34.6 | *** | 47.41 | *** | 7.12 | * | ||
| (C) | Time | 3 | 0.39 | 0.79 | 4.01 | 8.24 | ** | 5.57 | * | 10.81 | ** | 22.85 | *** | |||
| (D) | Time | 3 | 13.96 | *** | 8.8 | ** | 99.61 | *** | 436.11 | *** | 54.91 | *** | 63.05 | *** | 20.82 | |
| Site | 1 | 0.21 | 0.15 | 0.38 | 0.28 | 0.17 | 2.47 | 1.3 | ||||||||
| Time * Site | 3 | 2.06 | 0.77 | 1.41 | 2.06 | 2.23 | 0.57 | 1.65 | ||||||||
| (E) | Time | 3 | 1.58 | 3.79 | * | 47.78 | *** | 298.91 | *** | 18.42 | *** | 3.37 | * | 0.88 | ||
| Species | 1 | 7.05 | * | 395.01 | *** | 534.04 | *** | 1893.82 | *** | 665.24 | *** | 731.62 | *** | 119.17 | *** | |
| Time * Species | 3 | 2.1 | 0.9 | 46.58 | *** | 366.16 | *** | 14.35 | *** | 32.76 | *** | 20.8 | *** | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Zeng, H.; Chen, C.; Liu, W.; Jiang, X. Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition. Forests 2019, 10, 924. https://doi.org/10.3390/f10100924
Wu J, Zeng H, Chen C, Liu W, Jiang X. Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition. Forests. 2019; 10(10):924. https://doi.org/10.3390/f10100924
Chicago/Turabian StyleWu, Junen, Huanhuan Zeng, Chunfeng Chen, Wenjie Liu, and Xiaojin Jiang. 2019. "Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition" Forests 10, no. 10: 924. https://doi.org/10.3390/f10100924
APA StyleWu, J., Zeng, H., Chen, C., Liu, W., & Jiang, X. (2019). Intercropping the Sharp-Leaf Galangal with the Rubber Tree Exhibits Weak Belowground Competition. Forests, 10(10), 924. https://doi.org/10.3390/f10100924