
Magnesium Modifies the Structural Features of Enzymatically Mineralized Collagen Gels Affecting the Retraction Capabilities of Human Dermal Fibroblasts Embedded within This 3D System

 , and
, and
Abstract
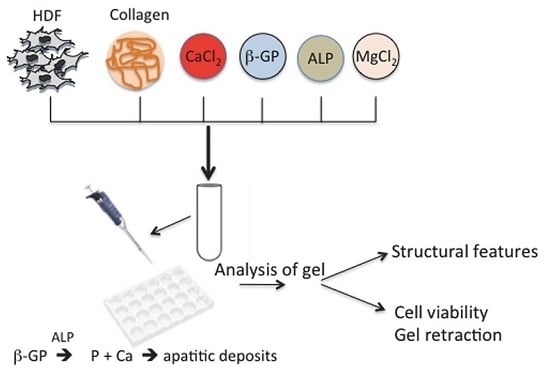
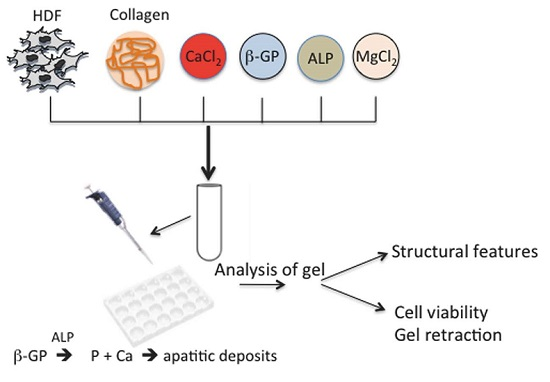
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
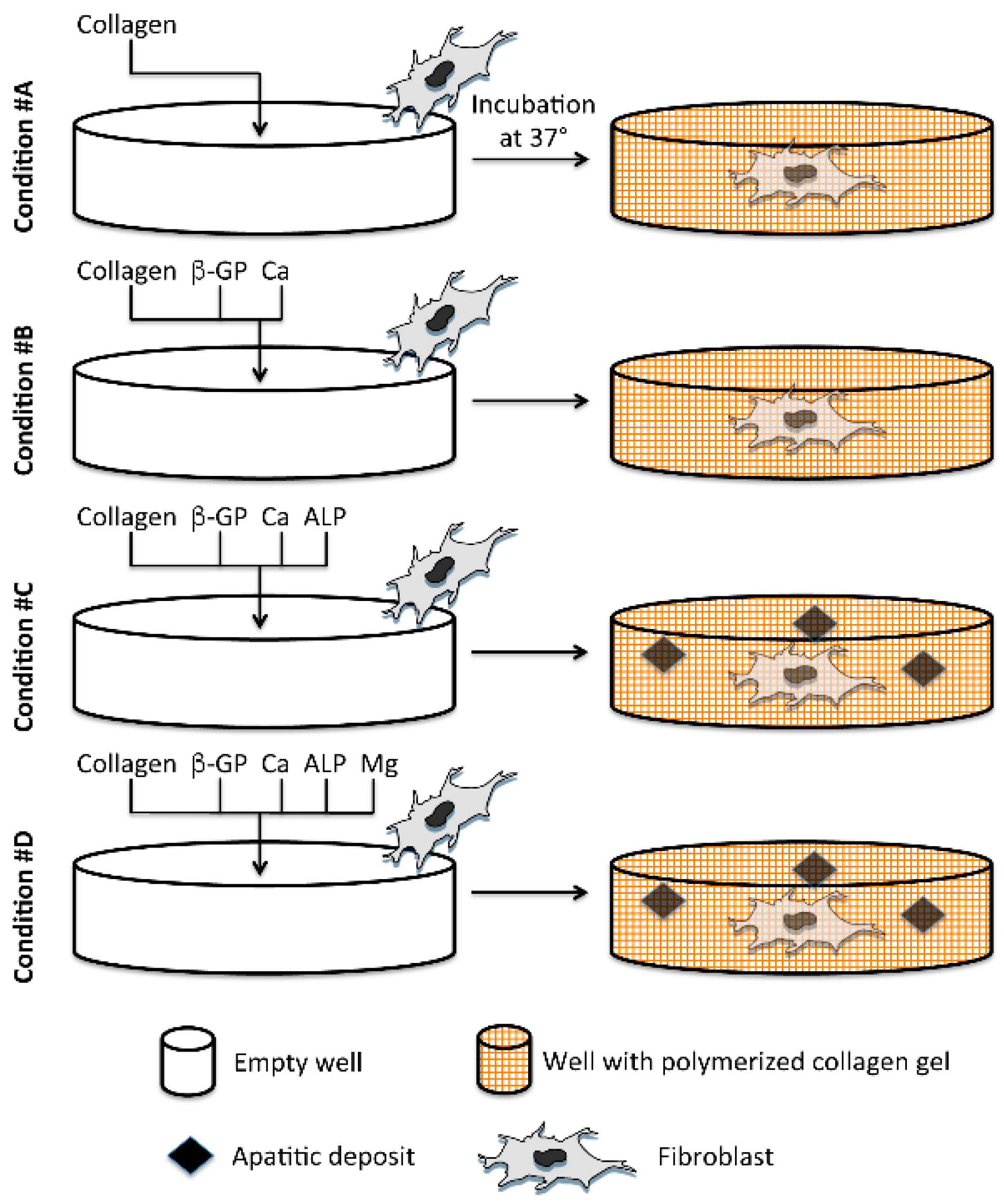
1. Introduction
2. Results
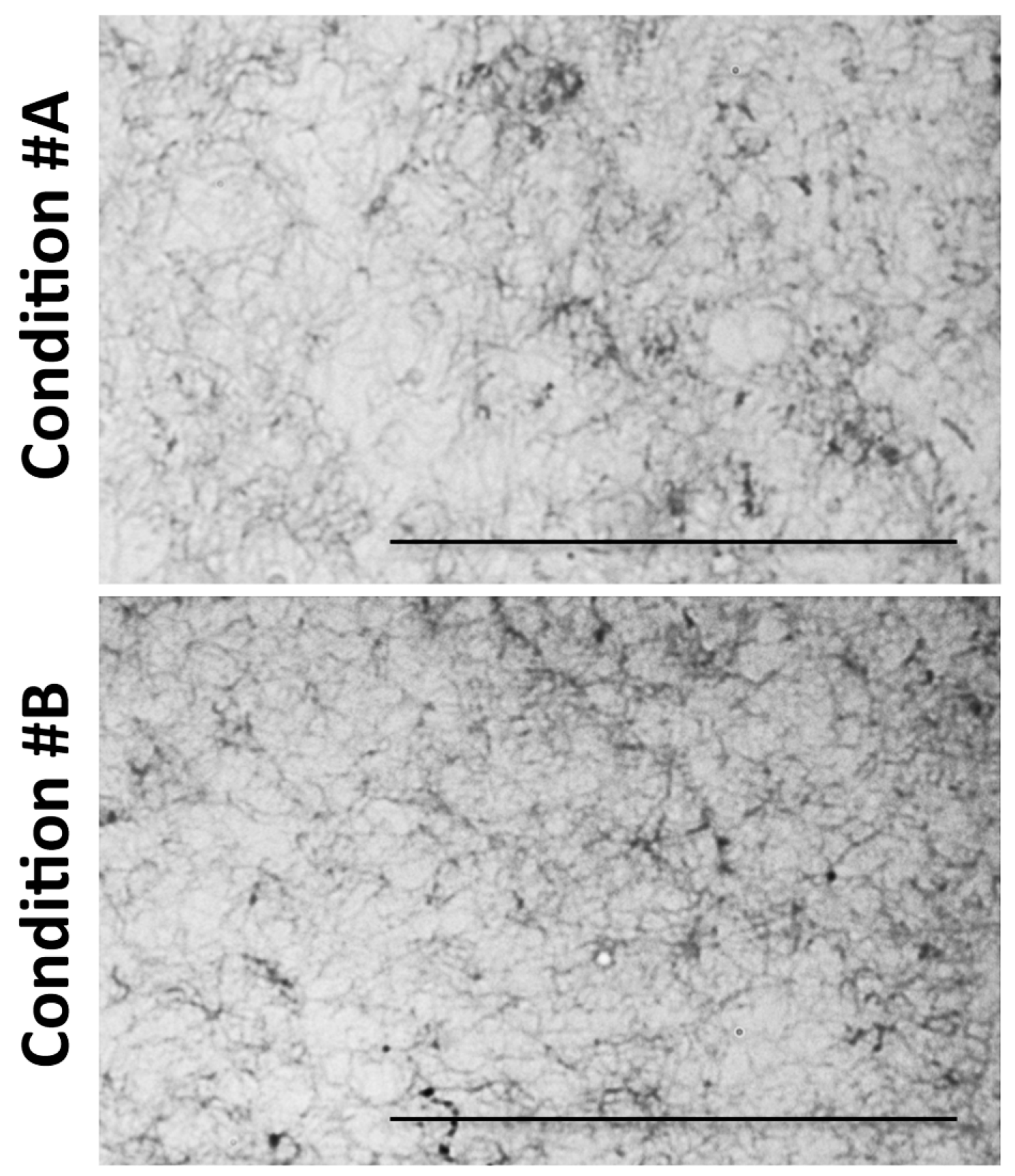
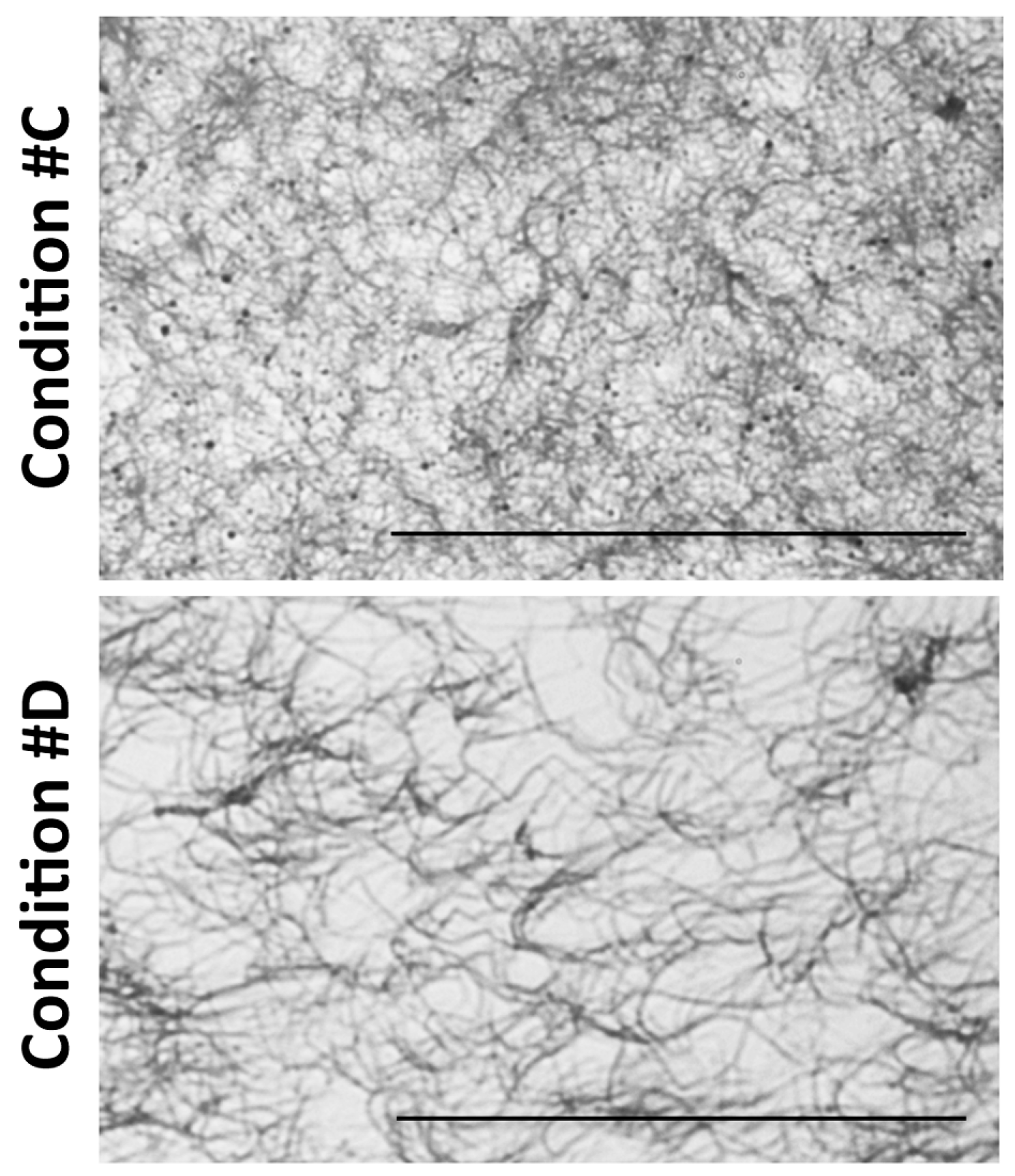
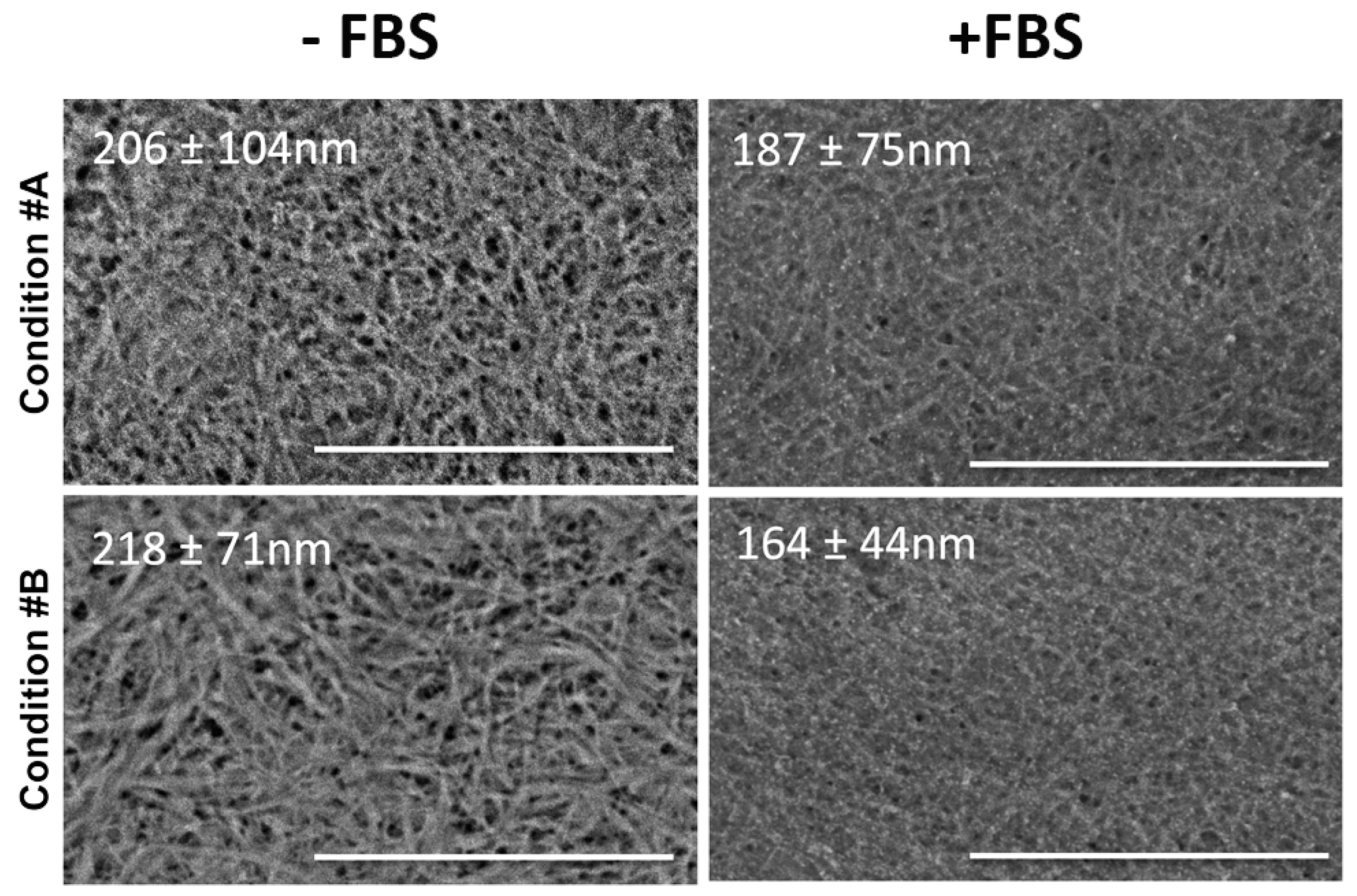
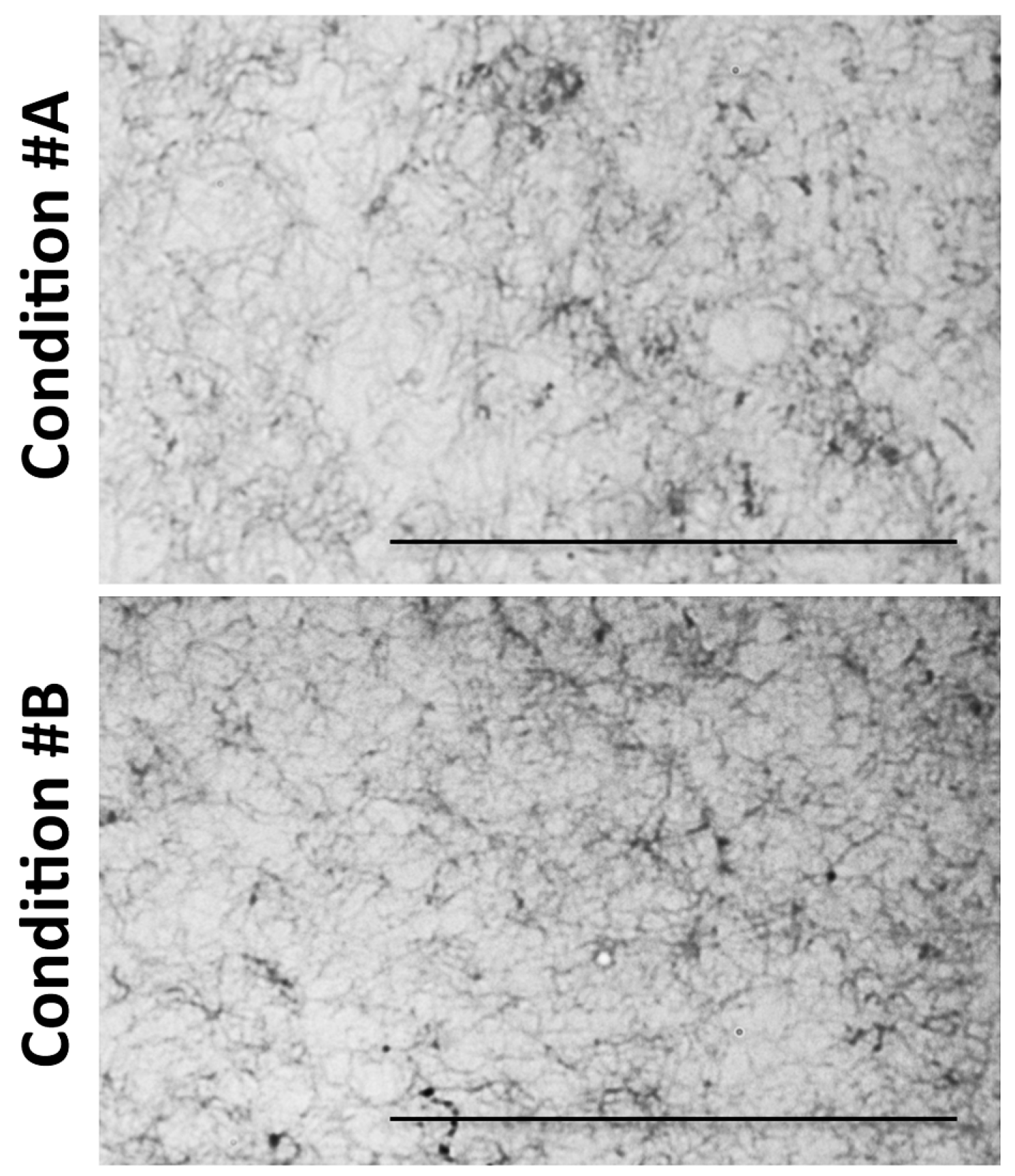
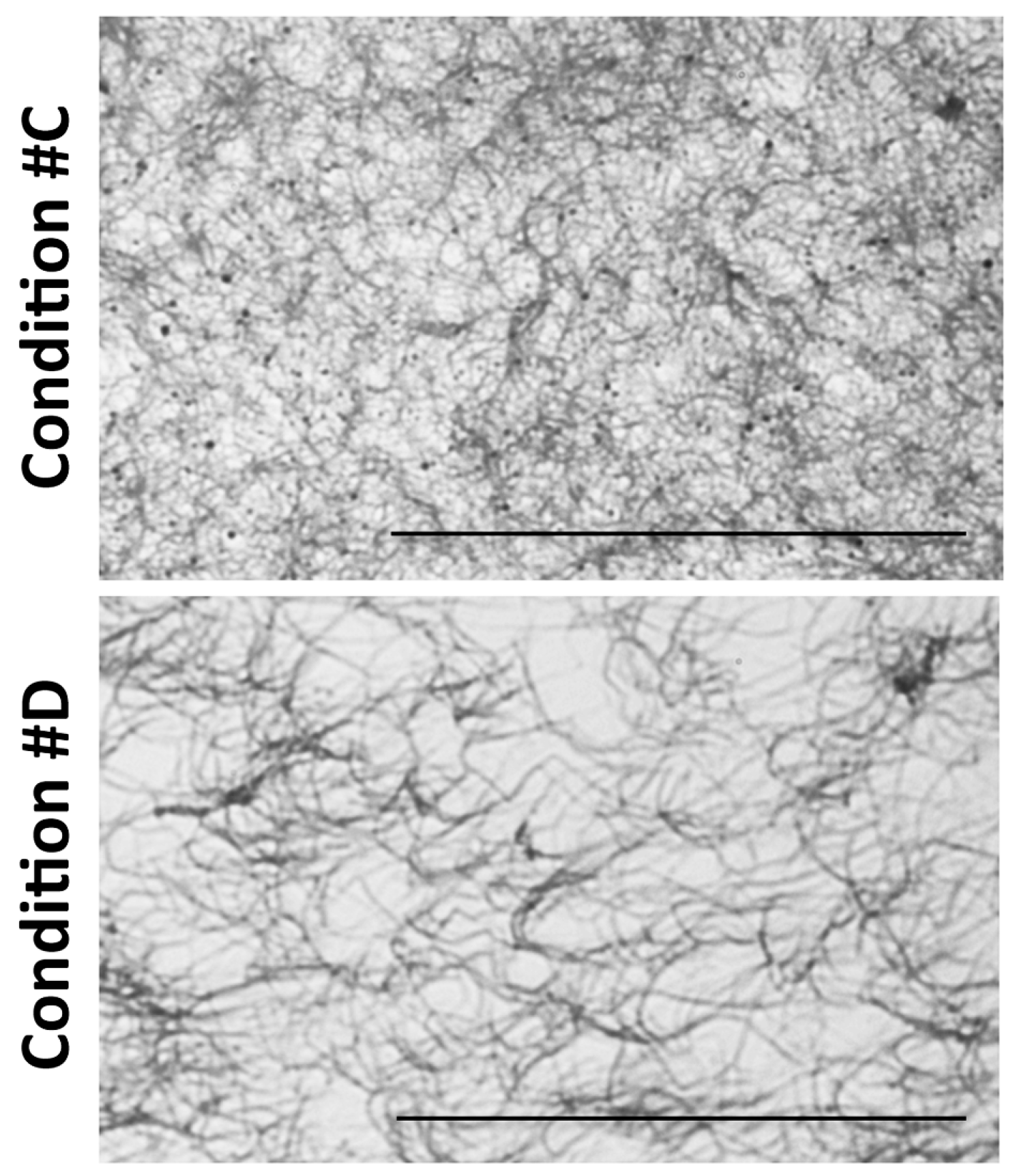
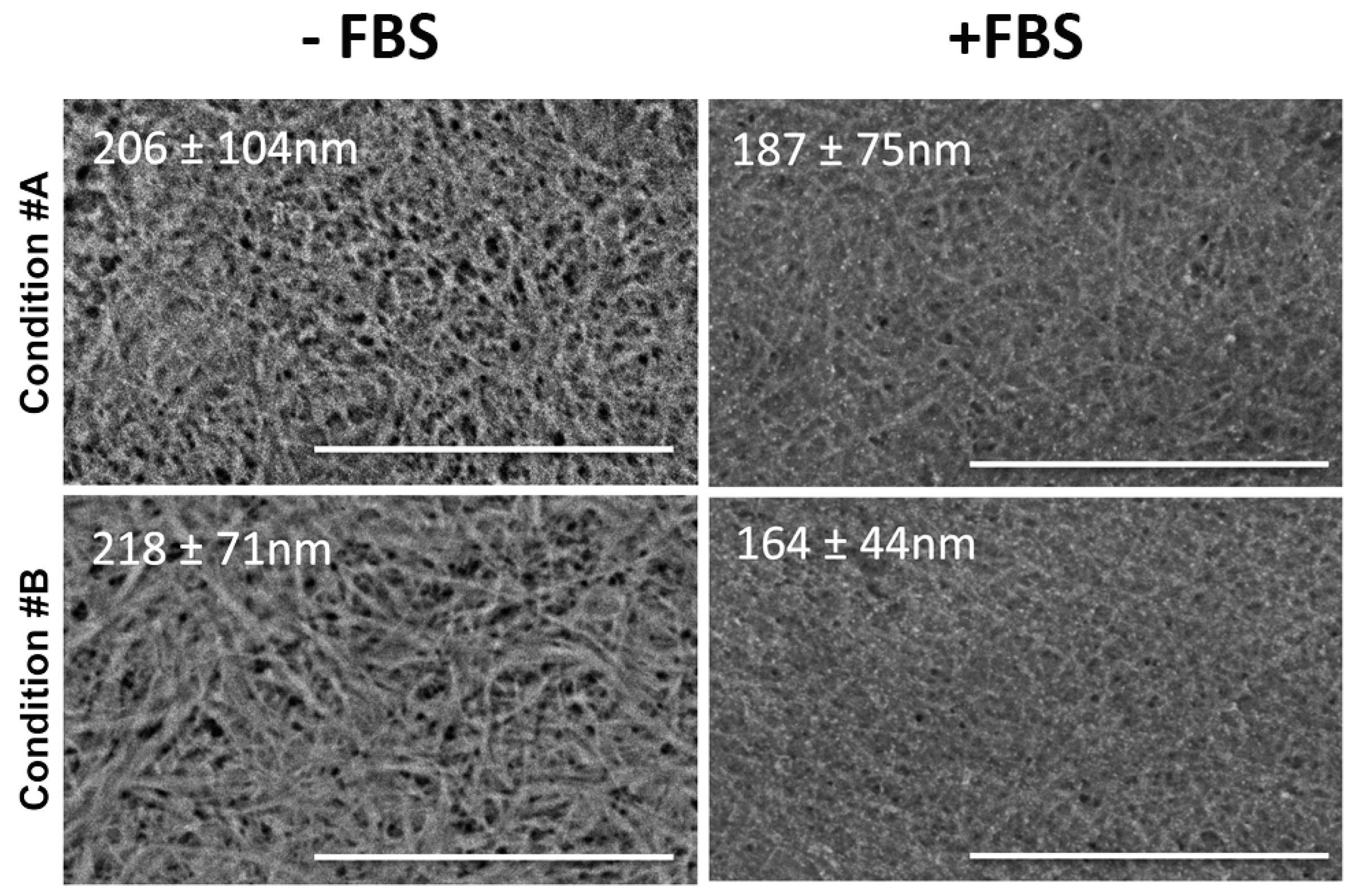
2.1. Light and Scanning Electron Microscopy Reveal a Different Structure of Collagen Gels
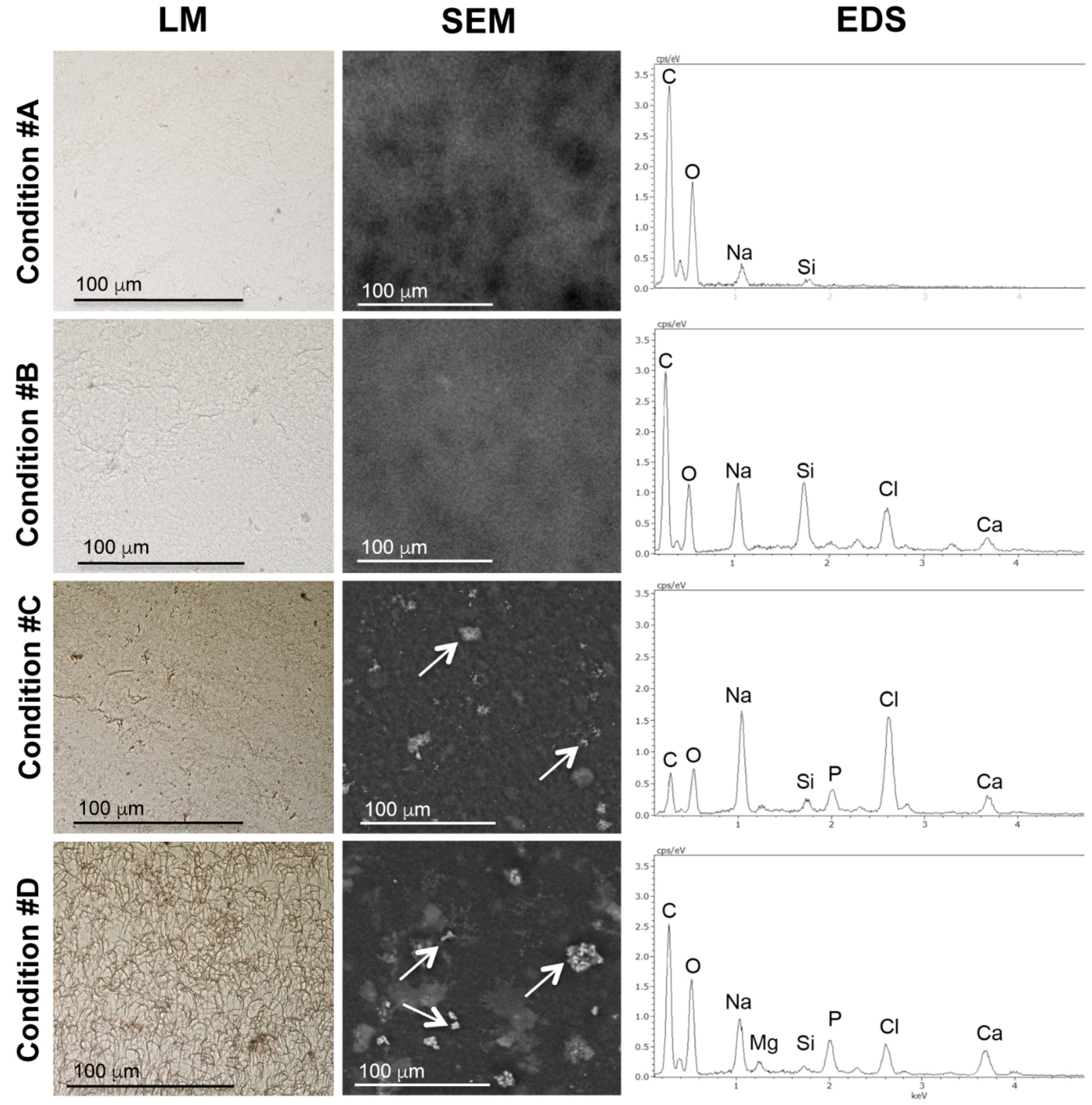
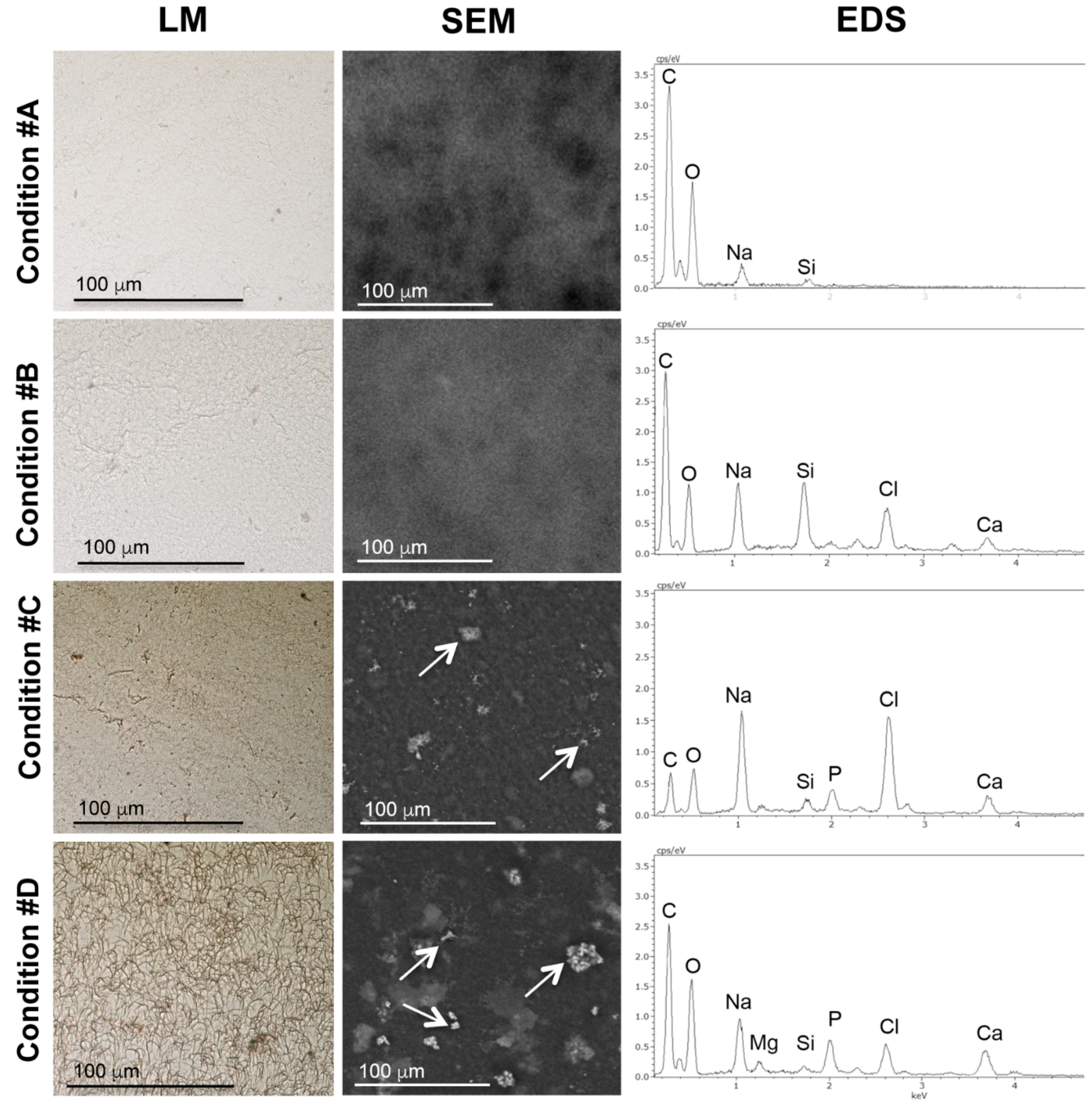
2.2. Mineral Deposition in Collagen Gels Requires ALP Activity
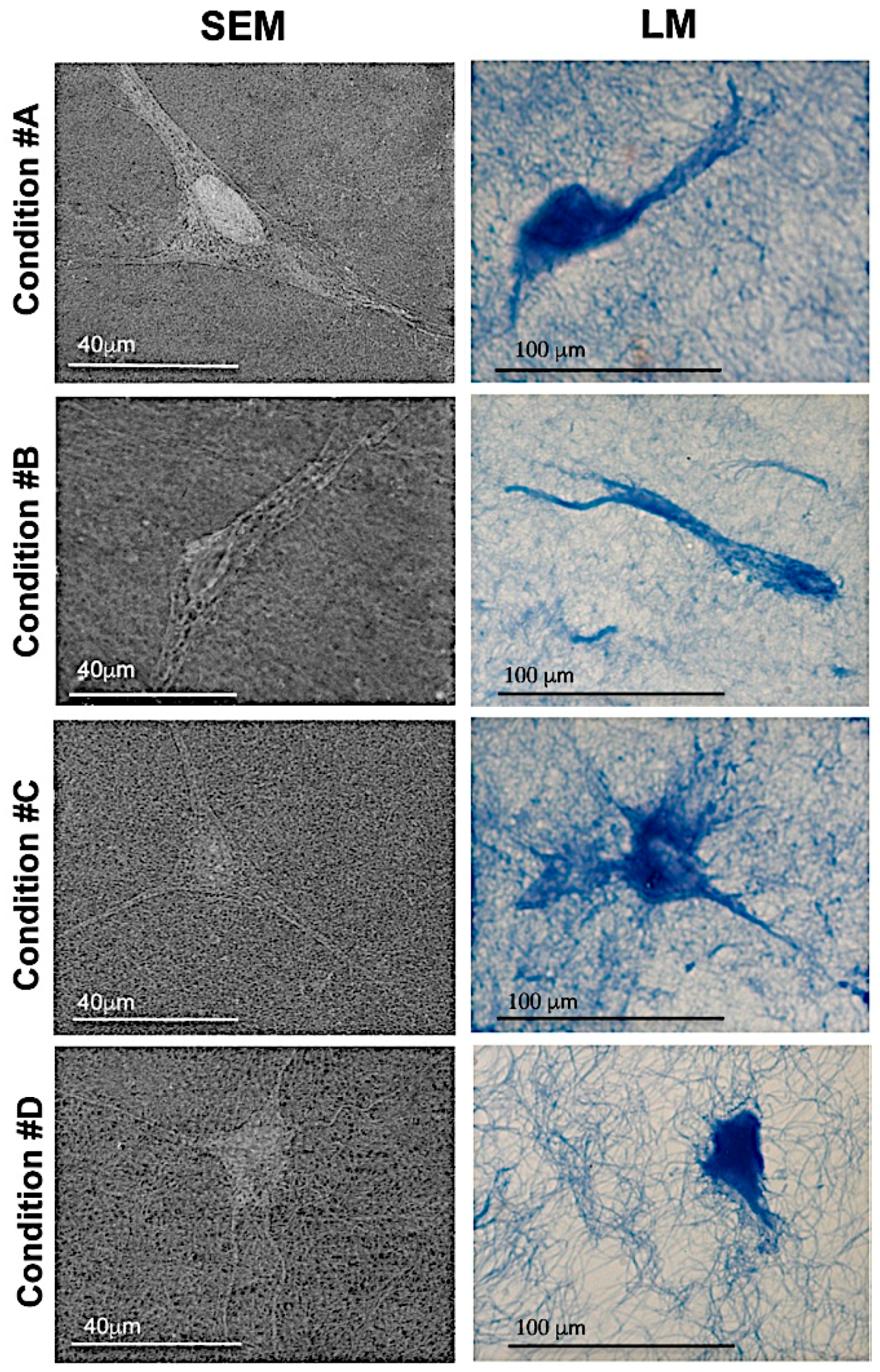
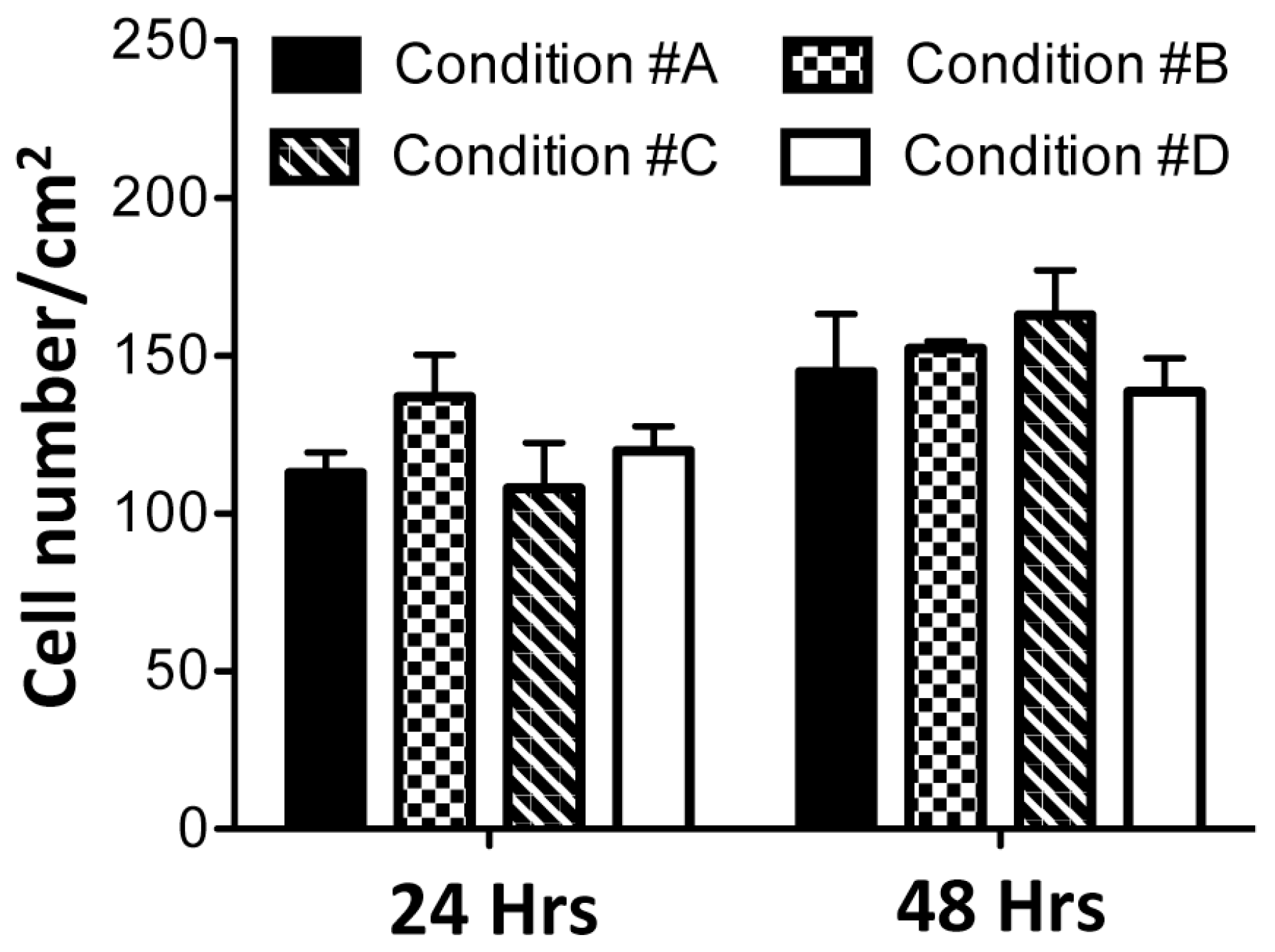
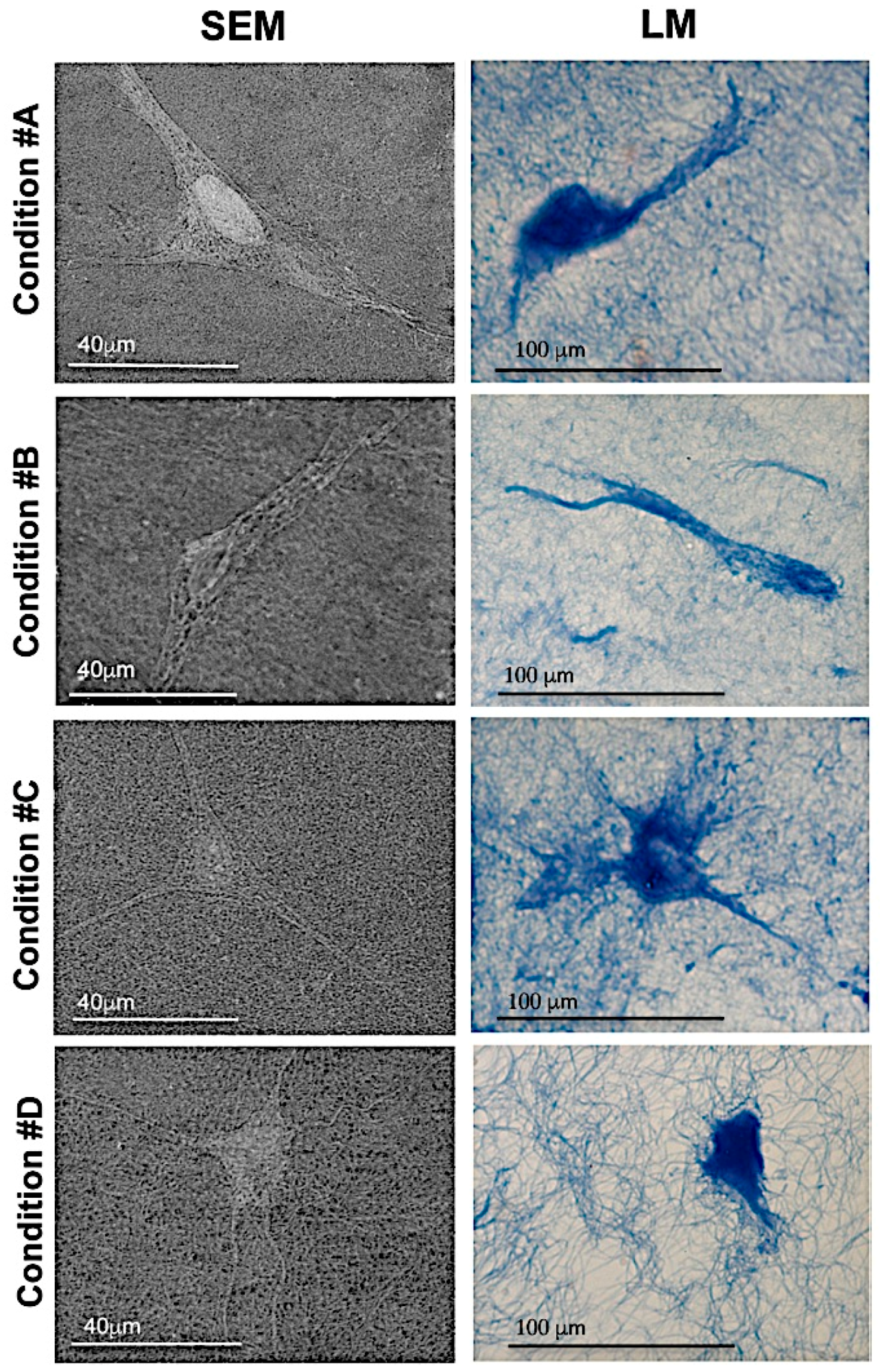
2.3. Homogeneous Distribution but Different Morphology of HDF within Collagen Gels
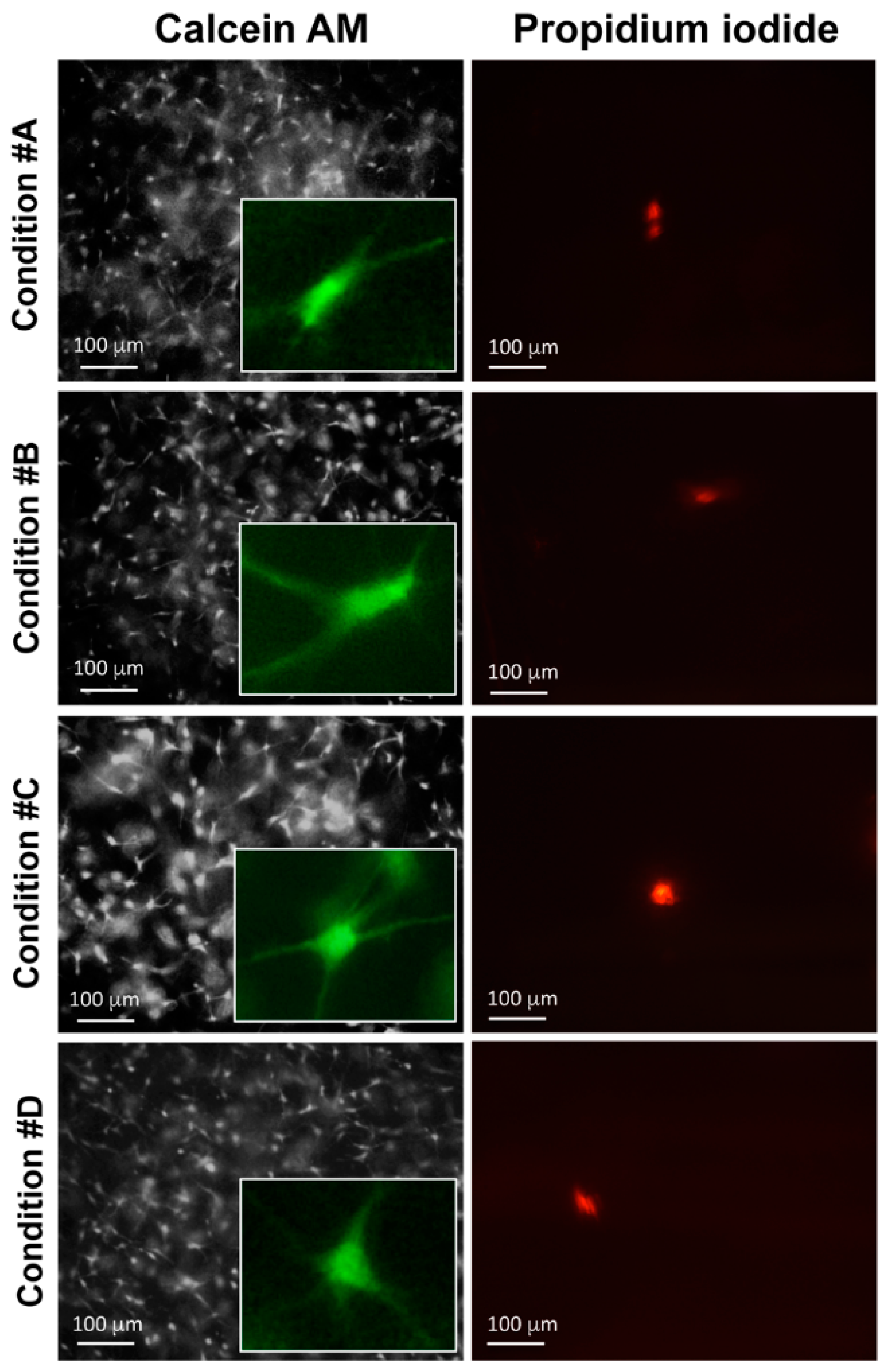
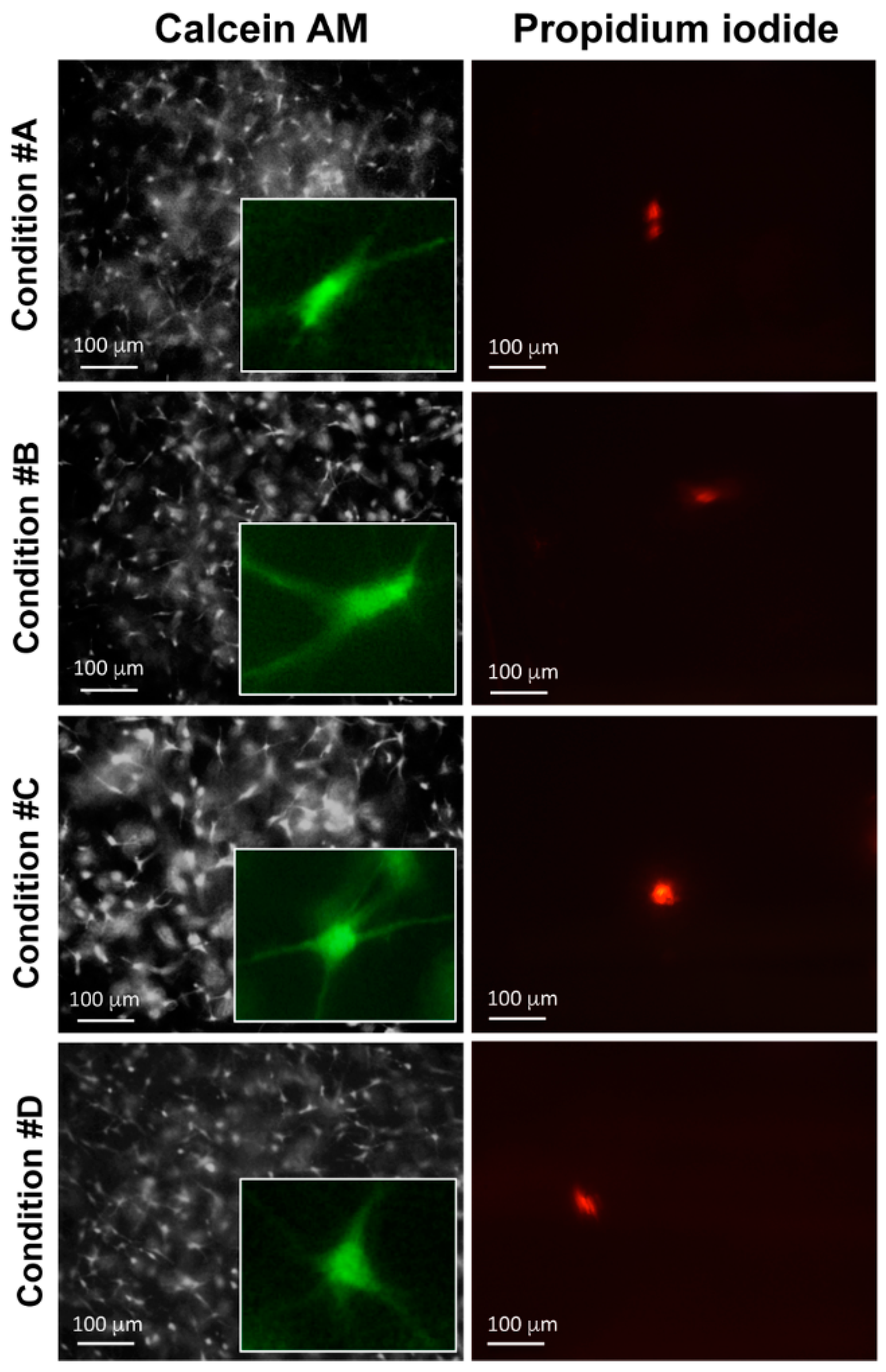
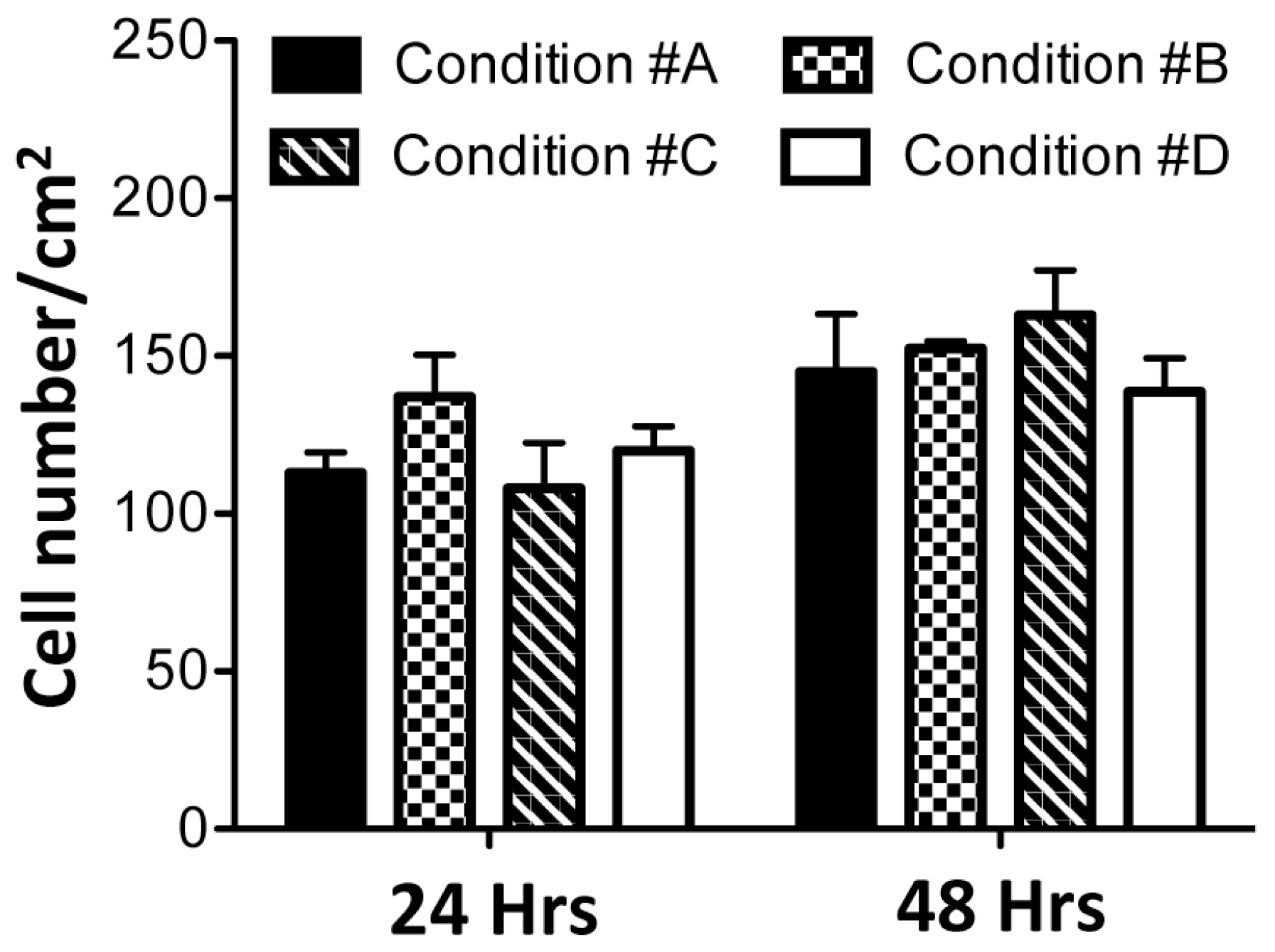
2.4. Enzymatically Mineralized Collagen Gel Is Cytocompatible
3. Discussion
4. Experimental Section
4.1. Preparation of Collagen Gels
4.2. Scanning (Low Vacuum Mode) and Scanning Transmission Electron Microscopy (SEM and SEM-STEM) and Energy-Dispersive Spectroscopy (EDS)
4.3. Histochemistry and Light Microscopy
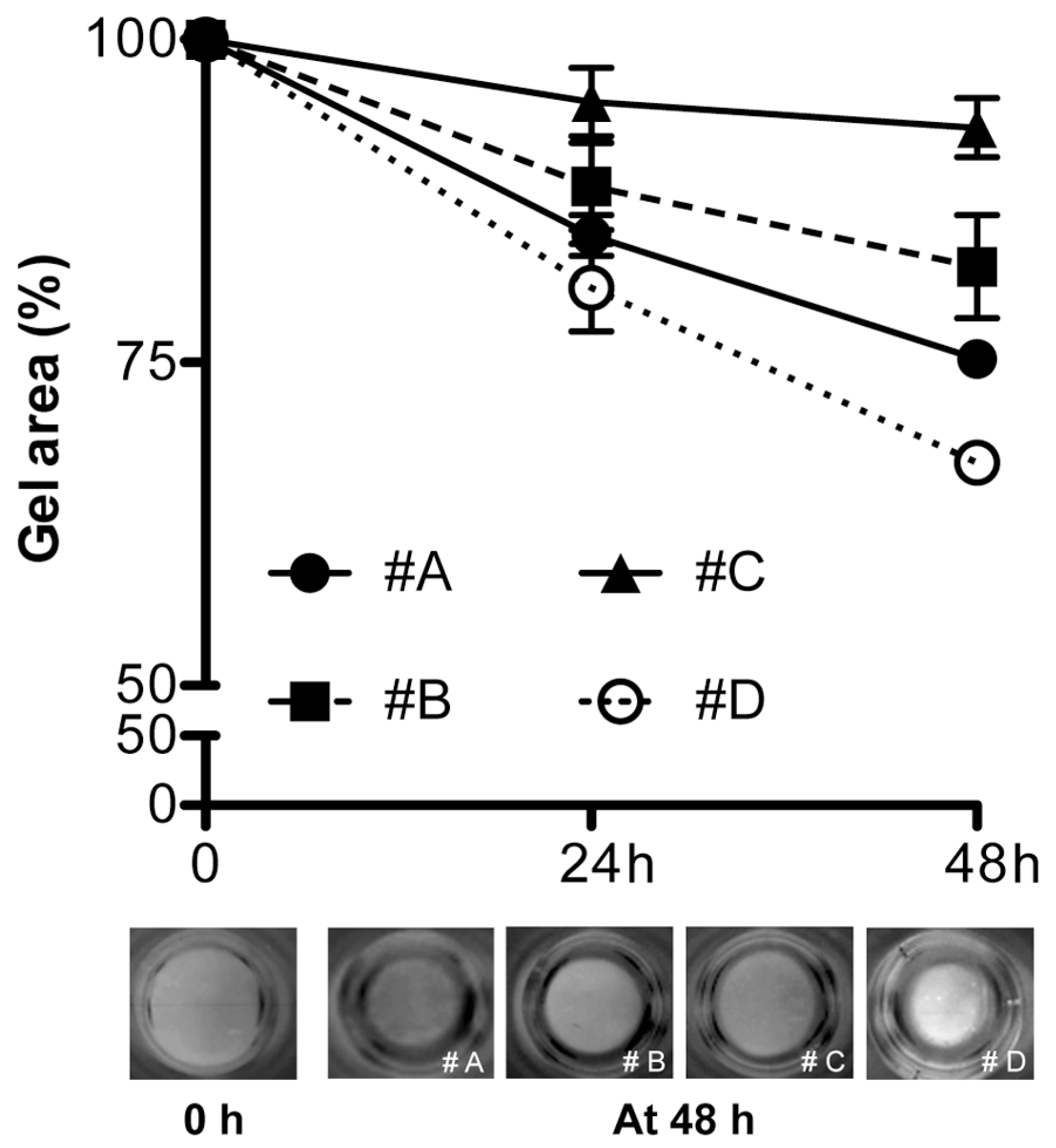
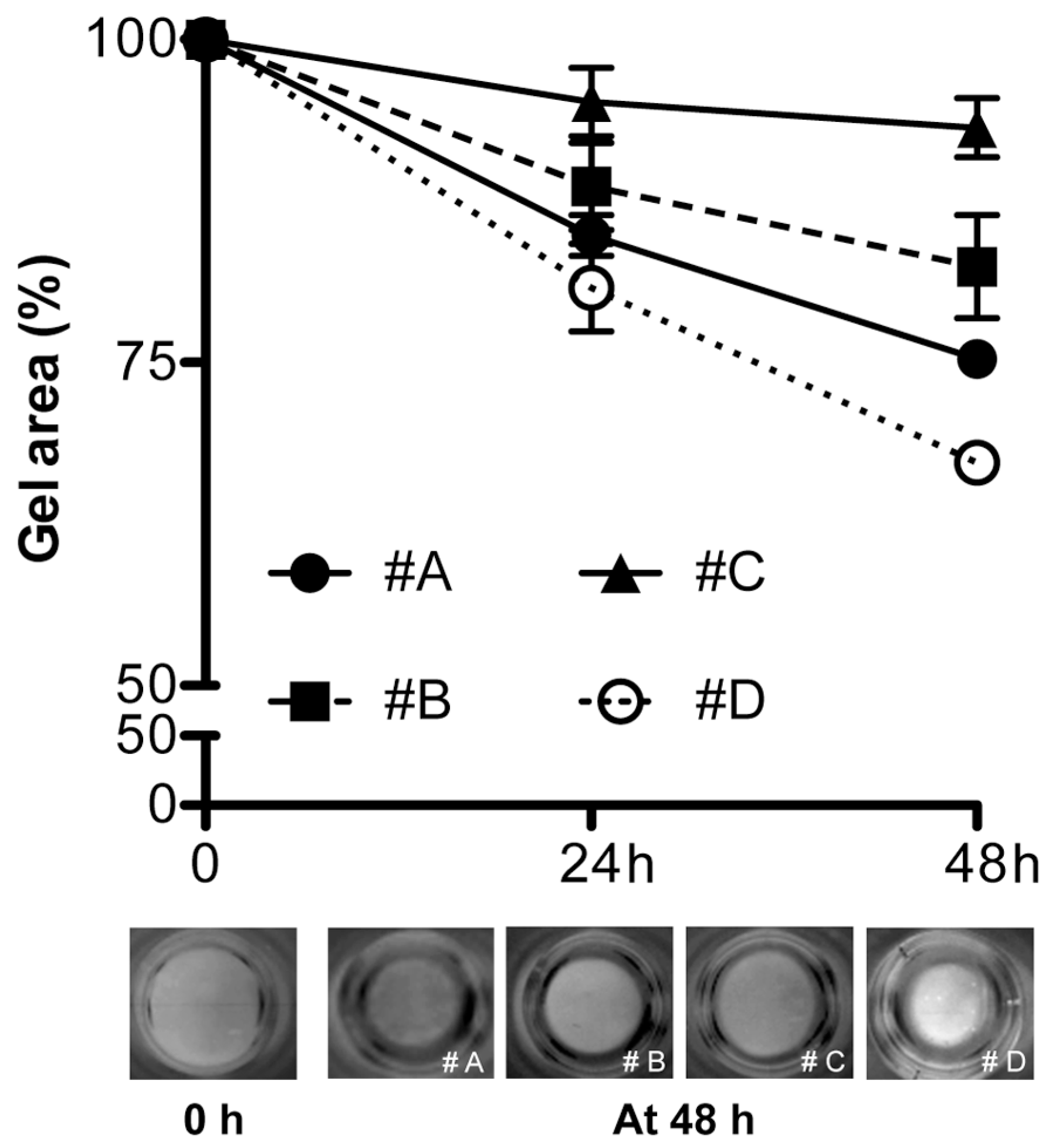
4.4. Gel Retraction and Live/Dead Assay
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Perez, R.A.; Won, J.E.; Knowles, J.C.; Kim, H.W. Naturally and synthetic Smart Composite Biomaterials for Tissue Regeneration. Adv. Drug Deliv. Rev. 2013, 65, 471–496. [Google Scholar] [CrossRef] [PubMed]
- Holzwarth, J.M.; Ma, P.X. Biomimetic Nanofibrous Scaffolds for Bone Tissue Engineering. Biomaterials 2011, 32, 9622–9629. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, J.P.; Plunkett, N.A.; O’Brien, F.J. Addition of hydroxyapatite improves stiffness, interconnectivity and osteogenic potential of a highly porous collagen-based scaffold for bone tissue regeneration. Eur. Cells Mater. 2010, 20, 218–230. [Google Scholar]
- Zhou, Y.; Yao, H.; Wang, J.; Wang, D.; Liu, Q.; Li, Z. Greener synthesis of electrospun collagen/hydroxyapatite composite fibers with an excellent microstructure for bone tissue engineering. Int. J. Nanomed. 2015, 10, 3203–3215. [Google Scholar]
- Wang, Y.; Azaïs, T.; Robin, M.; Vallée, A.; Catania, C.; Legriel, P.; Pehau-Arnaudet, G.; Babonneau, V.; Giraud-Guille, M.M.; Nassif, N. The predominant role of collagen in the nucleation, growth, structure and orientation of bone apatite. Nat. Mater. 2012, 11, 724–733. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Veis, A. Phosphorylated proteins and control over apatite nucleation, crystal growth, and inhibition. Chem. Rev. 2008, 108, 4670–4693. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.E.; Bonar, L.C.; Griffin, R.G.; Glimcher, M.J. Characterization of very young mineral phases of bone by solid state 31phosphorus magic angle sample spinning nuclear magnetic resonance and X-ray diffraction. Calcif. Tissue Int. 1992, 50, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Olszta, J.M.; Chenga, X.; Jeea, S.S.; Kumara, R.; Kima, Y.Y.; Kaufmane, M.J.; Douglasa, E.P.; Laurie, B. Bone structure and formation: A new perspective. Mater. Sci. Eng. R 2007, 58, 77–116. [Google Scholar] [CrossRef]
- Silver, F.H.; Landis, W.J. Deposition of apatite in mineralizing vertebrate extracellular matrices: A model of possible nucleation sites on type I collagen. Connect. Tissue Res. 2001, 52, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.S.; Fang, P.A.; Zhang, X.; Jayaraman, T.; Sfeir, C.; Beniash, E. Primary structure and phosphorylation of dentin matrix protein 1 (DMP1) and dentin phosphophoryn (DPP) uniquely determine their role in biomineralization. Biomacromolecules 2011, 12, 2933–2945. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Goda, T.; Takeuchi, N.; Einaga, H.; Tanabe, T. Preparation of collagen/calcium phosphate multilayer sheet using enzymatic mineralization. Biomaterials 2004, 25, 5481–5489. [Google Scholar] [CrossRef] [PubMed]
- Douglas, T.E.; Messersmith, P.B.; Chasan, S.; Mikos, A.G.; de Mulder, E.L.; Dickson, G.; Schaubroeck, D.; Balcaen, L.; Vanhaecke, F.; Dubruel, P.; et al. Enzymatic mineralization of hydrogels for bone tissue engineering by incorporation of alkaline phosphatase. Macromol. Biosci. 2012, 12, 1077–1089. [Google Scholar] [CrossRef] [PubMed]
- Uchihashi, K.; Aoki, S.; Matsunobu, A.; Toda, S. Osteoblast migration into type I collagen gel and differentiation to osteocyte-like cells within a self-produced mineralized matrix: A novel system for analyzing differentiation from osteoblast to osteocyte. Bone 2013, 52, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, C.E.; Marelli, B.; Donelli, I.; Alessandrino, A.; Freddi, G.; Nazhat, S.N. Multilayered dense collagen–silk fibroin hybrid: A platform for mesenchymal stem cell differentiation towards chondrogenic and osteogenic lineages. J. Tissue Eng. Regen. Med. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Pastakia, M.; Fenn, M.B.; Kishore, V. Saos-2 Cell Mediated Mineralization on Collagen Gels: Effect of Densification and Bioglass Incorporation. J. Biomed. Mater. Res. A 2016, 104, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Williams, D.J. Incorporation of hydroxyapatite sol into collagen gel to regulate the contraction mediated by human bone marrow-derived stromal cells. IEEE Trans. Nanobiosci. 2010, 9, 1–11. [Google Scholar]
- Ronchetti, I.; Boraldi, F.; Annovi, G.; Cianciulli, P.; Quaglino, D. Fibroblast involvement in soft connective tissue calcification. Front. Genet. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Motte, S.; Kaufman, L.J. Pore size variable type I collagen gels and their interaction with glioma cells. Biomaterials 2010, 31, 5678–5688. [Google Scholar] [CrossRef] [PubMed]
- Sapudom, J.; Rubner, S.; Martin, S.; Kurth, T.; Riedel, S.; Mierke, C.T.; Pompe, T. The phenotype of cancer cell invasion controlled by fibril diameter and pore size of 3D collagen networks. Biomaterials 2015, 52, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Arnoult, O.; Smith, M.E.; Wnek, G.E. Electrospinning of collagen nanofiber scaffolds from benign solvents. Macromol. Rapid Commun. 2009, 30, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Roeder, B.A.; Kokini, K.; Voytik-Harbin, S.L. Fibril microstructure affects strain transmission within collagen extracellular matrices. J. Biomech. Eng. 2009, 131. [Google Scholar] [CrossRef] [PubMed]
- LaRusso, J.; Li, Q.; Jiang, Q.; Uitto, J. Elevated dietary magnesium prevents connective tissue mineralization in a mouse model of pseudoxanthoma elasticum (Abcc6(-/-)). J. Investig. Dermatol. 2009, 129, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, Y.; Li, P.; Song, Y. Anion-specific effects on the assembly of collagen layers mediated by magnesium ion on mica surface. J. Phys. Chem. B 2014, 118, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Eidelman, N.; Boyde, A.; Bushby, A.J.; Howell, P.G.; Sun, J.; Newbury, D.E.; Miller, F.W.; Robey, P.G.; Rider, L.G. Microstructure and mineral composition of dystrophic calcification associated with the idiopathic inflammatory myopathies. Arthritis Res. Ther. 2009, 11. [Google Scholar] [CrossRef] [PubMed]
- Delogne, C.; Lawford, P.V.; Habesch, S.M.; Carolan, V.A. Characterization of the calcification of cardiac valve bioprostheses by environmental scanning electron microscopy and vibrational spectroscopy. J. Microsc. 2007, 228, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Yamanishi, H.; Sato, V. The removal of non-collagen components from newborn calf dermis with magnesium chloride solution. J. Biochem. 1976, 79, 131–144. [Google Scholar] [PubMed]
- Brana, C.; Benham, C.; Sundstrom, L. A method for characterising cell death in vitro by combining propidium iodide staining with immunohistochemistry. Brain Res. Protoc. 2002, 10, 109–114. [Google Scholar] [CrossRef]
- Addison, W.N.; Nelea, V.; Chicatun, F.; Chien, Y.C.; Tran-Khanh, N.; Buschmann, M.D.; Nazhat, S.N.; Kaartinen, M.T.; Vali, H.; Tecklenburg, M.M.; et al. Extracellular matrix mineralization in murine MC3T3-E1 osteoblast cultures: An ultrastructural, compositional and comparative analysis with mouse bone. Bone 2015, 71, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Widbiller, M.; Lindner, S.R.; Buchalla, W.; Eidt, A.; Hiller, K.A.; Schmalz, G.; Galler, K.M. Three-dimensional culture of dental pulp stem cells in direct contact to tricalcium silicate cements. Clin. Oral Investig. 2016, 20, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Steitz, S.A.; Speer, M.Y.; Curinga, G.; Yang, H.Y.; Haynes, P.; Aebersold, R.; Schinke, T.; Karsenty, G.; Giachelli, C.M. Smooth muscle cell phenotypic transition associated with calcification: Upregulation of Cbfa1 and downregulation of smooth muscle lineage markers. Circ. Res. 2001, 89, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Boraldi, F.; Annovi, G.; Vermeer, C.; Schurgers, L.J.; Trenti, T.; Tiozzo, R.; Guerra, D.; Quaglino, D. Matrix gla protein and alkaline phosphatase are differently modulated in human dermal fibroblasts from PXE patients and controls. J. Investig. Dermatol. 2013, 133, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Boraldi, F.; Annovi, G.; Bartolomeo, A.; Quaglino, D. Fibroblasts from patients affected by Pseudoxanthoma elasticum exhibit an altered PPi metabolism and are more responsive to pro-calcifying stimuli. J. Dermatol. Sci. 2014, 74, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Banks, E.; Nakajima, S.; Shapiro, L.C.; Tilevitz, O. Fibrous apatite on modified collagen. Science 1997, 198, 1164–1166. [Google Scholar] [CrossRef]
- Doi, Y.; Horiguchi, T.; Moriwaki, Y.; Kitago, H.; Kajimoto, T.; Iwayama, Y. Formation of apatite-collagen complexes. J. Biomed. Mater. Res. 1996, 31, 43–49. [Google Scholar] [CrossRef]
- Grinnell, F. Fibroblast-collagen-matrix contraction: Growth-factor signalling and mechanical loading. Trends Cell Biol. 2000, 10, 362–365. [Google Scholar] [CrossRef]
- Freyman, T.M.; Yannas, I.V.; Yokoo, R.; Gibson, L.J. Fibroblast contractile force is independent of the stiffness which resists the contraction. Exp. Cell Res. 2002, 272, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Kunz, W.; Henle, J.; Ninham, B.W. Zur Lehre von der Wirkung der Salze’ (about the science of the effect of salts): Franz Hofmeister’s historical papers. Curr. Opin. Colloid Interface Sci. 2004, 9, 19–37. [Google Scholar] [CrossRef]
- Farraro, K.F.; Kim, K.E.; Woo, S.L.; Flowers, J.R.; McCullough, M.B. Revolutionizing orthopaedic biomaterials: The potential of biodegradable and bioresorbable magnesium-based materials for functional tissue engineering. J. Biomech. 2014, 47, 1979–1986. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S. Morphogenesis of calcification in porcine bioprosthesis: Insight from high resolution electron microscopic investigation at molecular and atomic resolution. J. Electron. Microsc. 1993, 42, 156–165. [Google Scholar]
- Huang-Lee, L.H.H.; Cheung, D.T.; Nimni, M.E. Biochemical changes and cytotoxicity associated with the degradation of polymeric glutaraldehyde derived cross-links. J. Biomed. Mater. Res. 1990, 24, 1185–1201. [Google Scholar] [CrossRef] [PubMed]
- Mavrilasa, D.; Apostolakia, A.; Kapolosb, J.; Koutsoukosb, P.G.; Melachrinouc, M.; Zolotac, V.; Dougenisd, D. Development of bioprosthetic heart valve calcification in vitro and in animal models: Morphology and composition. J. Cryst. Growth 1999, 4, 554–562. [Google Scholar] [CrossRef]
- Louvet, L.; Bazin, D.; Büchel, J.; Steppan, S.; Passlick-Deetjen, J.; Massy, Z.A. Characterisation of calcium phosphate crystals on calcified human aortic vascular smooth muscle cells and potential role of magnesium. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Gorgieva, S.; Strancar, J.; Kokol, V. Evaluation of surface/interface-related physicochemical and microstructural properties of gelatin 3D scaffolds, and their influence on fibroblast growth and morphology. J. Biomed. Mater. Res. Part A 2014, 102, 3986–3997. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, M.; Wittel, F.K.; Herrmann, H.J. Simulation of flow of mixtures through anisotropic porous media using a lattice Boltzmann model. Eur. Phys. J. E Soft Matter. 2010, 32, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Menezes, G.C.; Miron-Mendoza, M.; Ho, C.H.; Jiang, H.; Grinnell, F. Oncogenic Ras-transformed human fibroblasts exhibit differential changes in contraction and migration in 3D collagen matrices. Exp. Cell Res. 2008, 314, 3081–3091. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F.; Ho, C.H. The effect of growth factor environment on fibroblast morphological response to substrate stiffness. Biomaterials 2013, 34, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Tamariz, E.; Grinnell, F. Modulation of fibroblast morphology and adhesion during collagen matrix remodelling. Mol. Biol. Cell 2002, 13, 3915–3929. [Google Scholar] [CrossRef] [PubMed]
- Quaglino, D.; Boraldi, F.; Barbieri, D.; Croce, A.; Tiozzo, R.; Pasquali Ronchetti, I. Abnormal phenotype of in vitro dermal fibroblasts from patients with Pseudoxanthoma elasticum (PXE). Biochim. Biophys. Acta 2000, 1501, 51–62. [Google Scholar] [CrossRef]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boraldi, F.; Bartolomeo, A.; Annovi, G.; Debret, R.; Quaglino, D. Magnesium Modifies the Structural Features of Enzymatically Mineralized Collagen Gels Affecting the Retraction Capabilities of Human Dermal Fibroblasts Embedded within This 3D System. Materials 2016, 9, 477. https://doi.org/10.3390/ma9060477
Boraldi F, Bartolomeo A, Annovi G, Debret R, Quaglino D. Magnesium Modifies the Structural Features of Enzymatically Mineralized Collagen Gels Affecting the Retraction Capabilities of Human Dermal Fibroblasts Embedded within This 3D System. Materials. 2016; 9(6):477. https://doi.org/10.3390/ma9060477
Chicago/Turabian StyleBoraldi, Federica, Angelica Bartolomeo, Giulia Annovi, Romain Debret, and Daniela Quaglino. 2016. "Magnesium Modifies the Structural Features of Enzymatically Mineralized Collagen Gels Affecting the Retraction Capabilities of Human Dermal Fibroblasts Embedded within This 3D System" Materials 9, no. 6: 477. https://doi.org/10.3390/ma9060477
APA StyleBoraldi, F., Bartolomeo, A., Annovi, G., Debret, R., & Quaglino, D. (2016). Magnesium Modifies the Structural Features of Enzymatically Mineralized Collagen Gels Affecting the Retraction Capabilities of Human Dermal Fibroblasts Embedded within This 3D System. Materials, 9(6), 477. https://doi.org/10.3390/ma9060477