
Impact of Polyrhodanine Manganese Ferrite Binary Nanohybrids (PRHD@MnFe2O4) on Osteoblasts and Osteoclasts Activities—A Key Factor in Osteoporosis Treatment

 , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Fabrication of the PRHD@MnFe2O4 Binary Nanohybrids
2.2. Cell Lines and Culture
2.3. Cell Viability Assay
2.4. Fluorescent Microscopy
2.5. Analysis of mRNA Expression Profiles
2.6. Statistical Analysis
3. Results
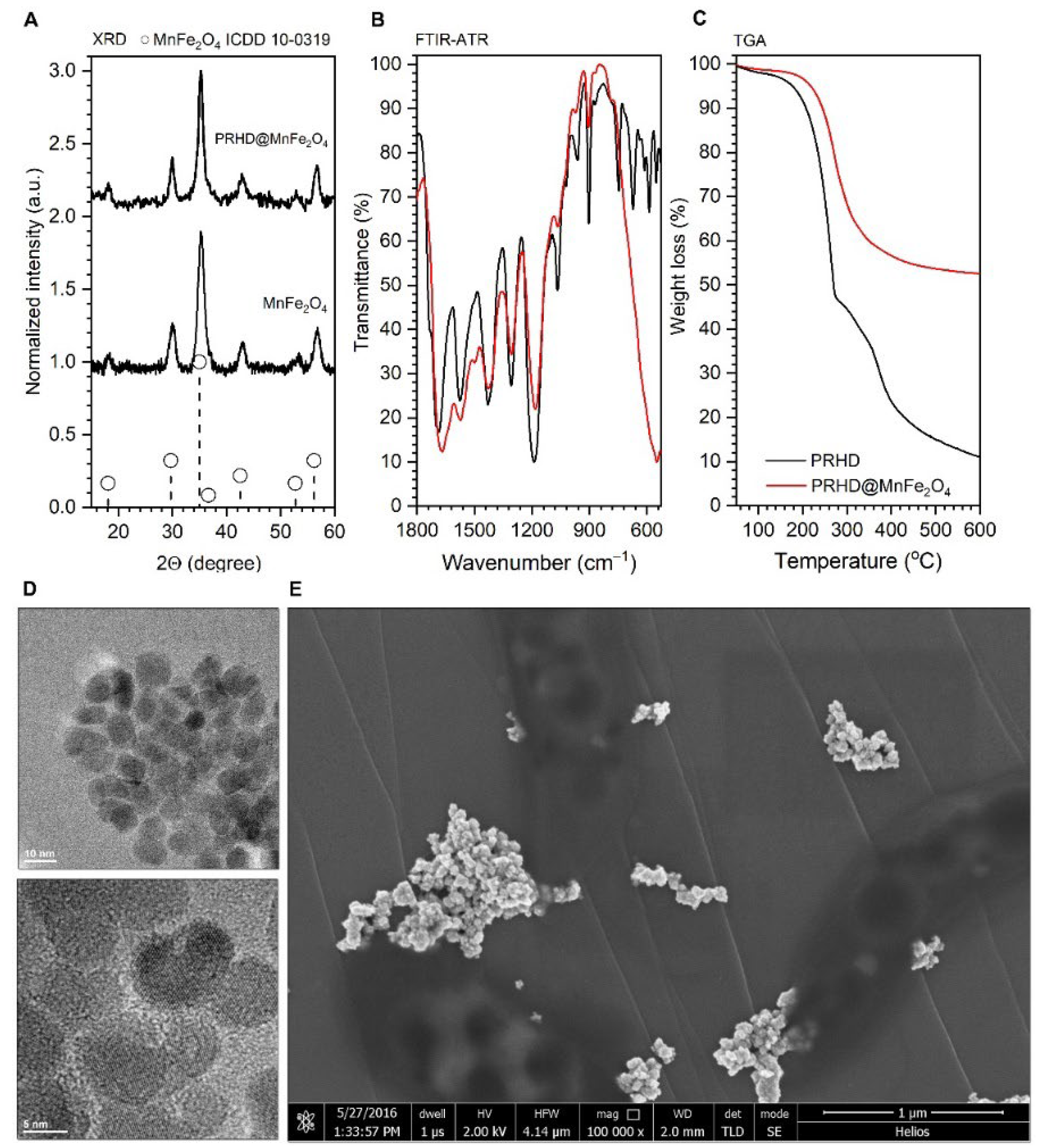
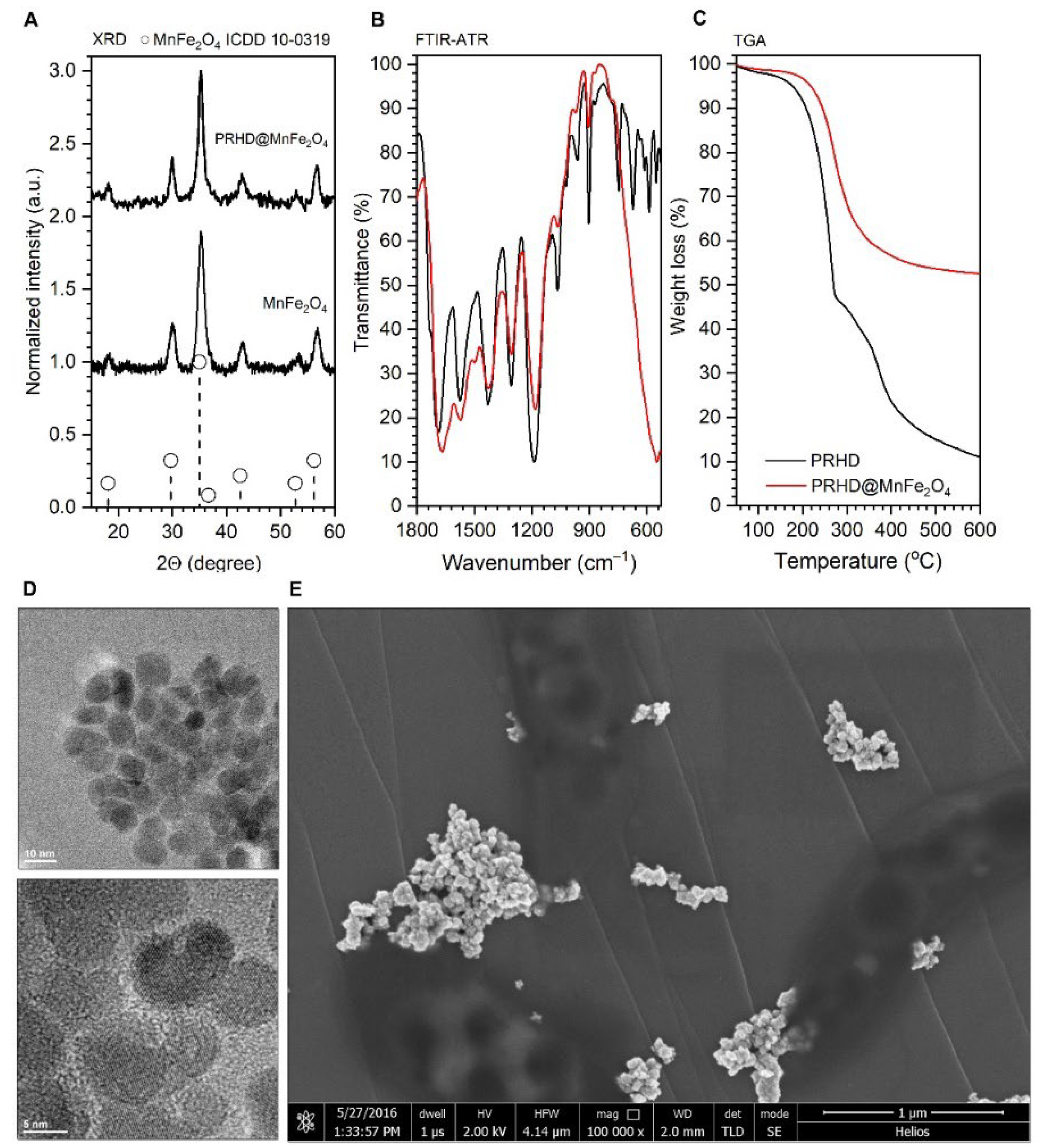
3.1. Physicochemical Properties of the PRHD@MnFe2O4 Binary Hybrids
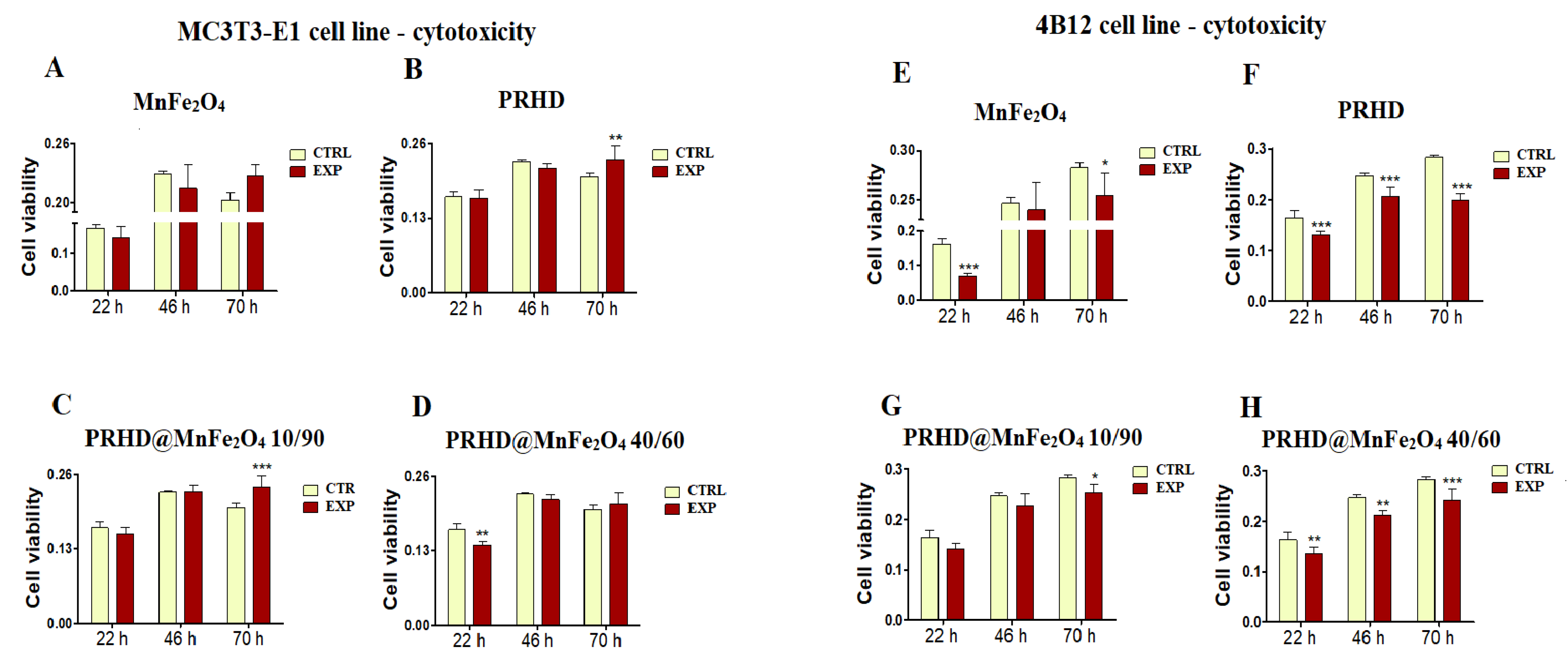
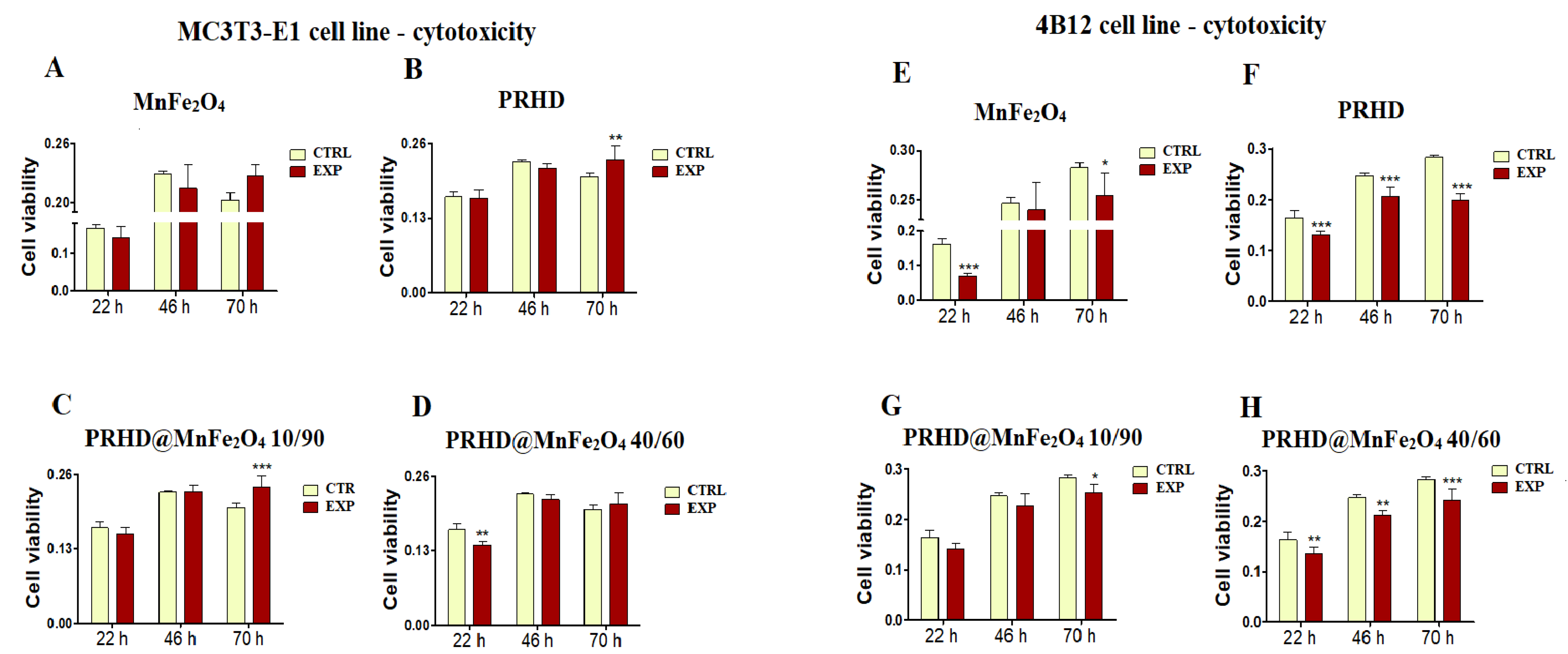
3.2. Cytotoxicity of the PRHD@MnFe2O4 at Different Concentration Ratios towards Osteoblasts and Osteoclasts
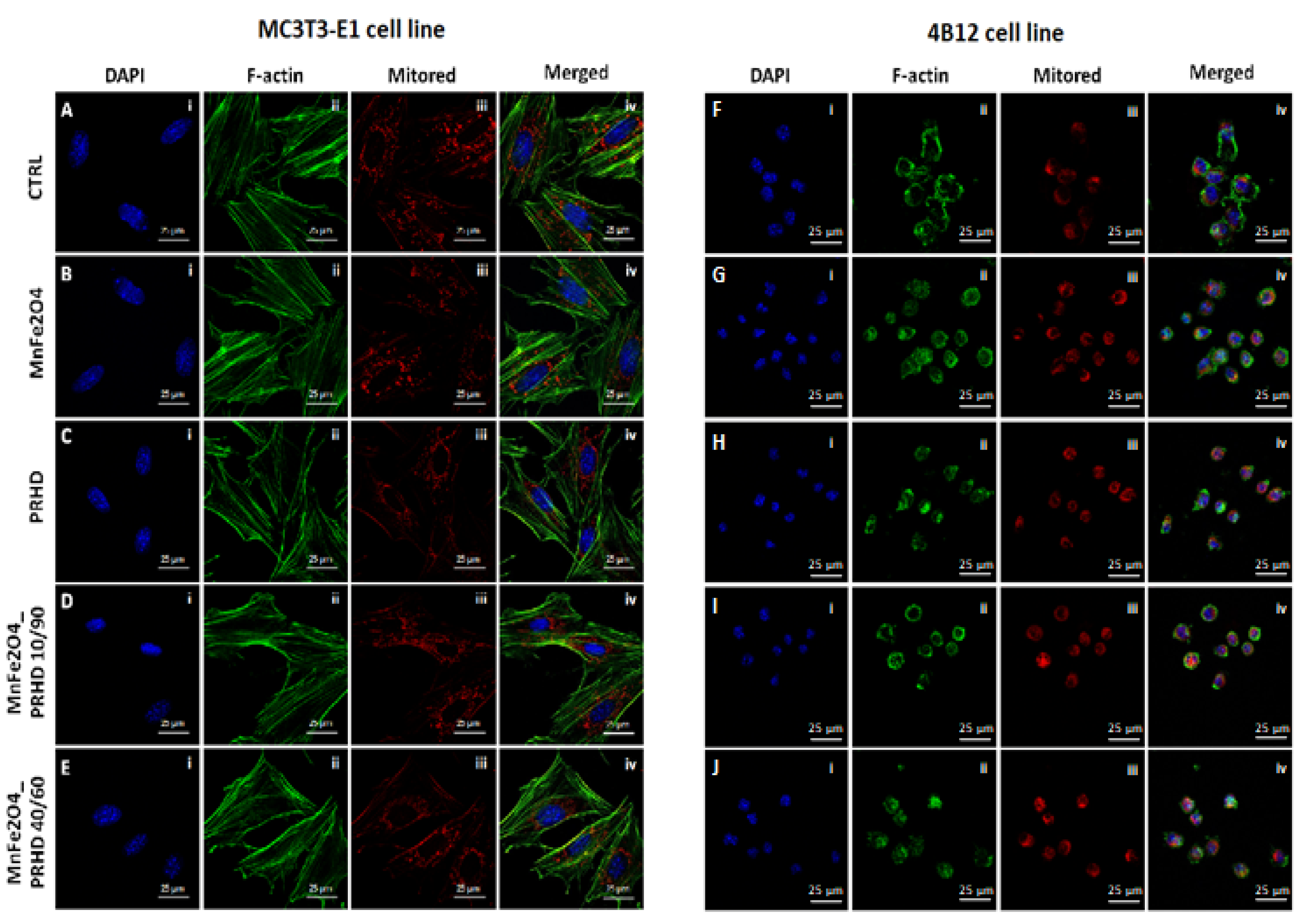
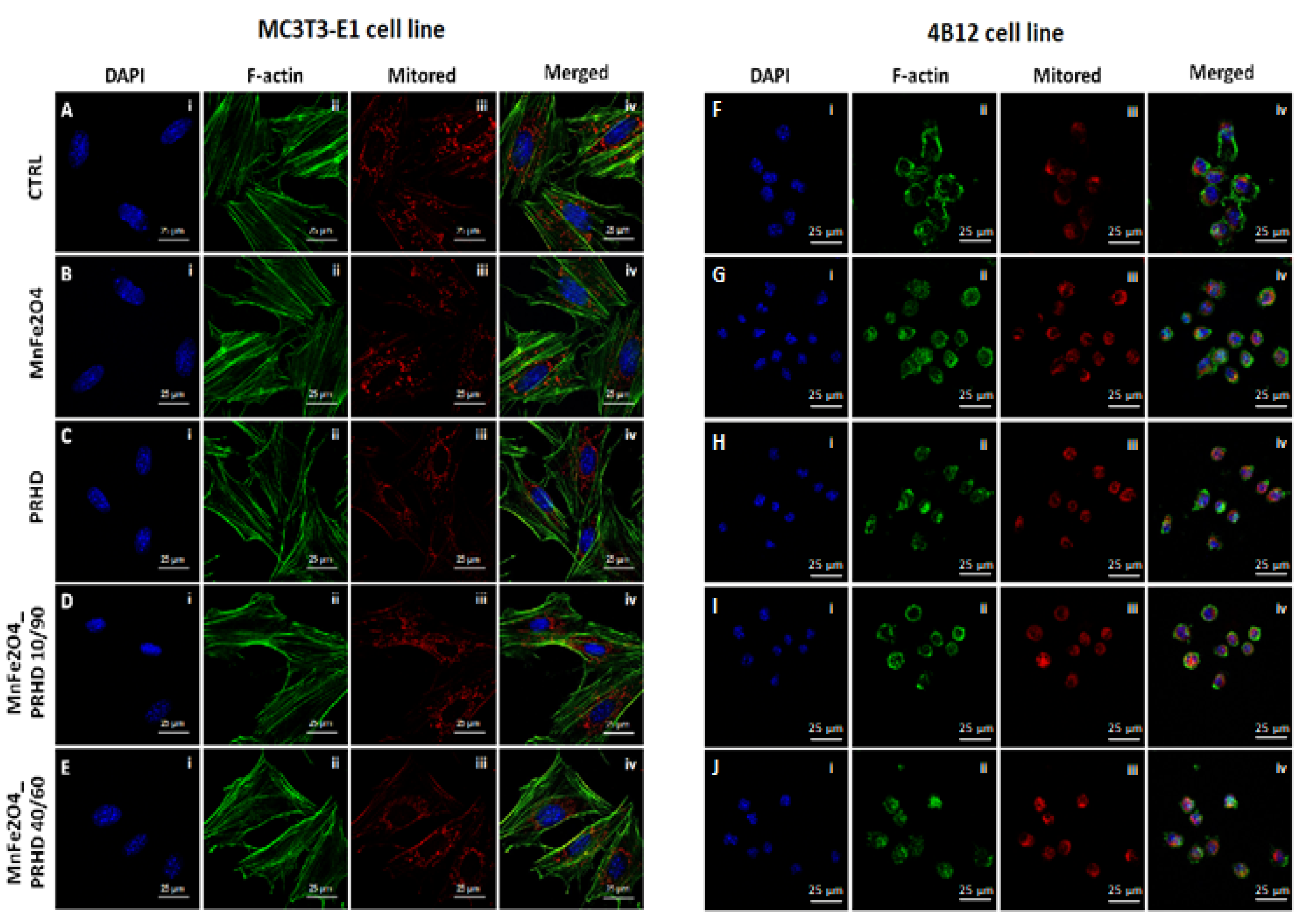
3.3. The Impact of the PRHD@MnFe2O4 on the Morphology and Mitochondrial Status of Osteoblasts and Osteoclasts
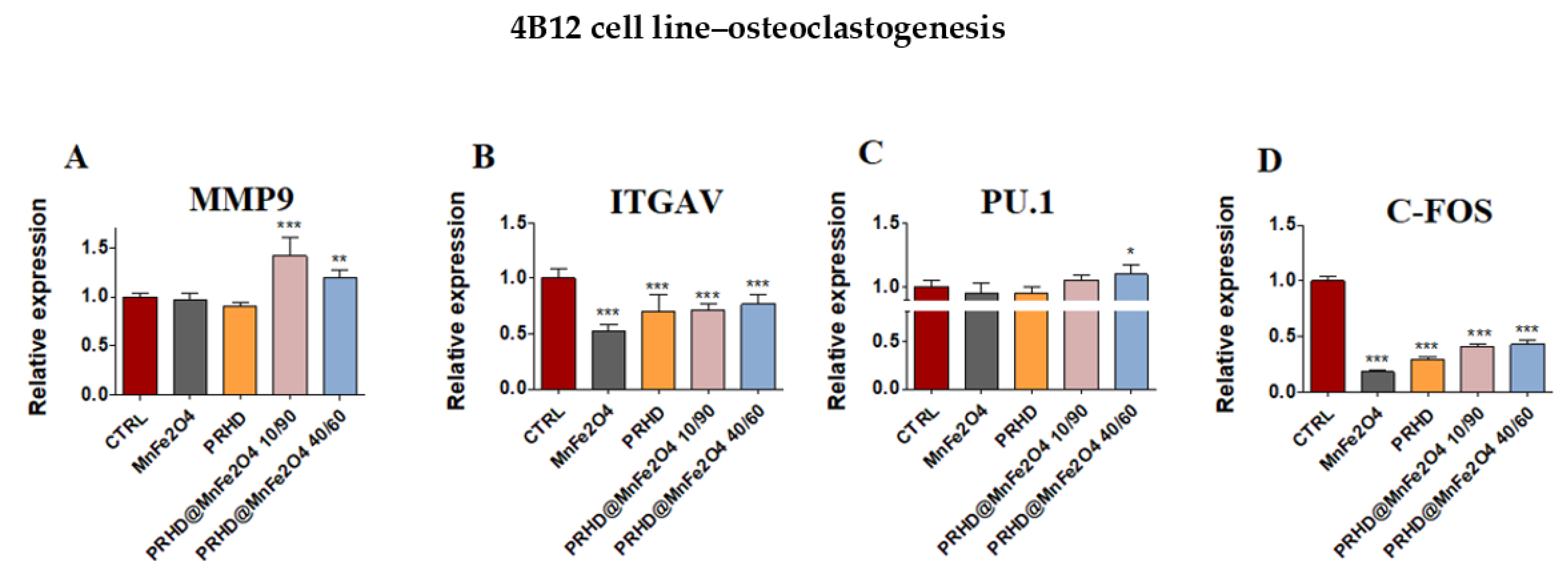
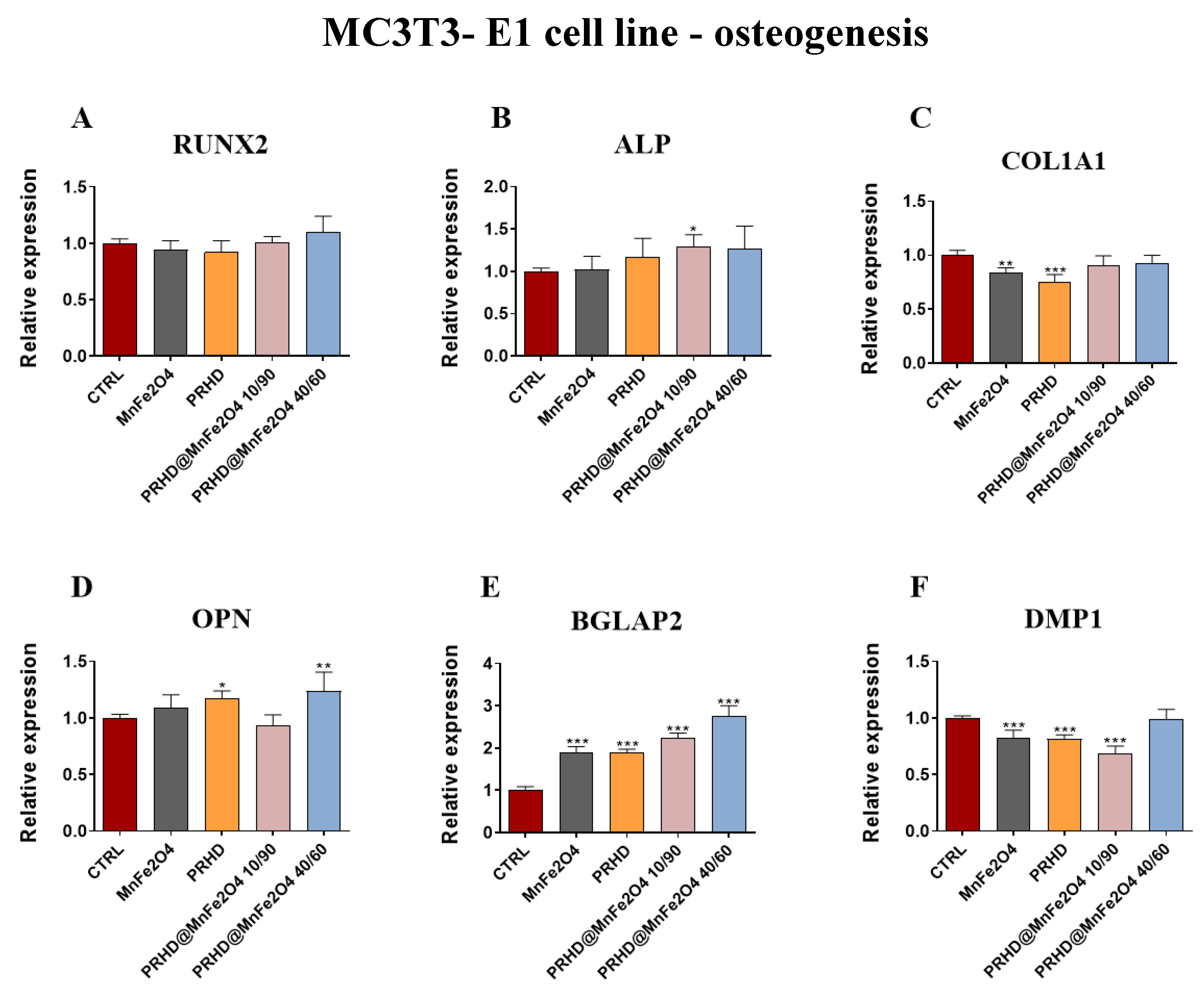
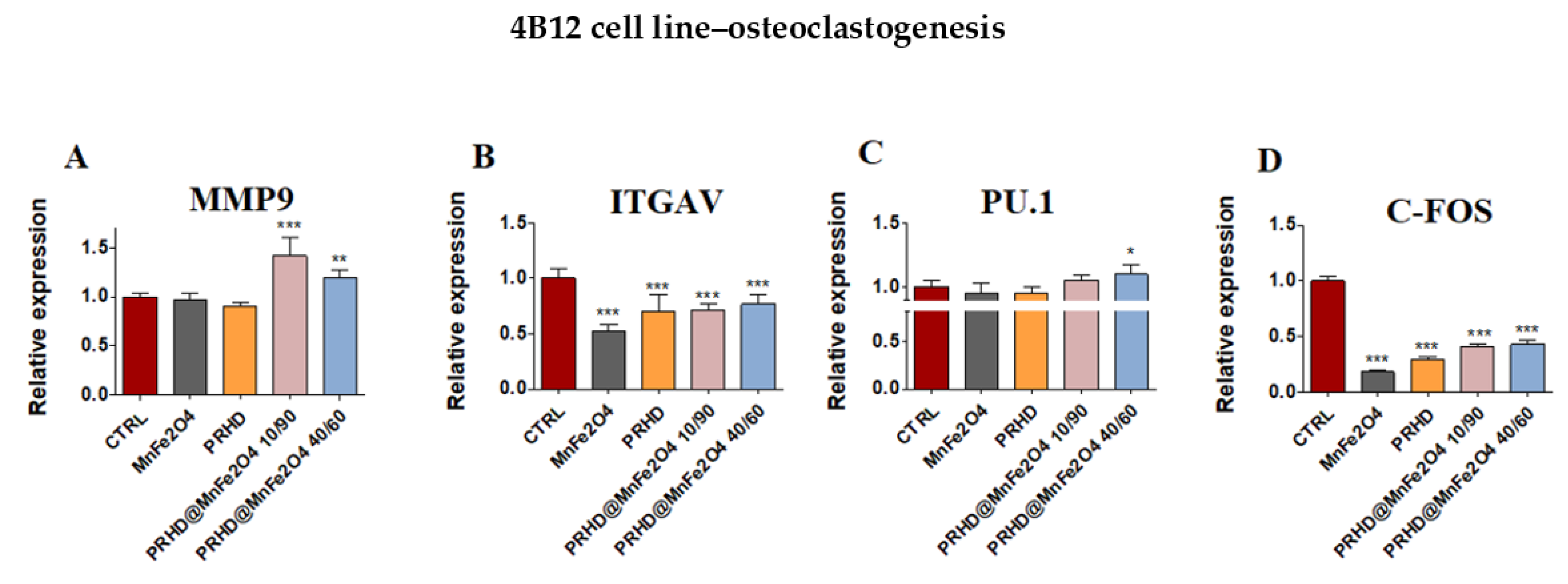
3.4. The Impact of the PRHD@MnFe2O4 on the Gene Expression in Osteoblasts and Osteoclasts
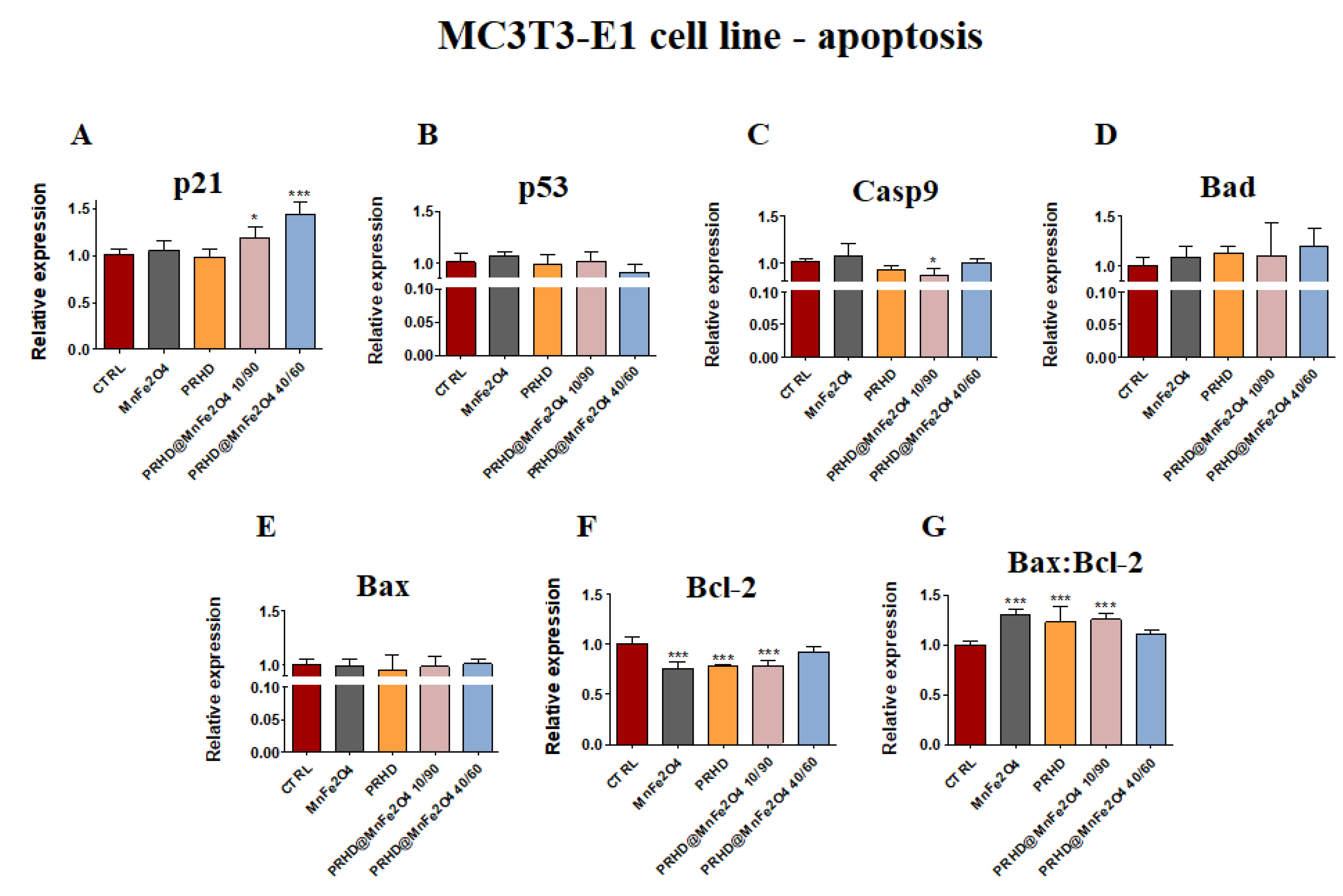
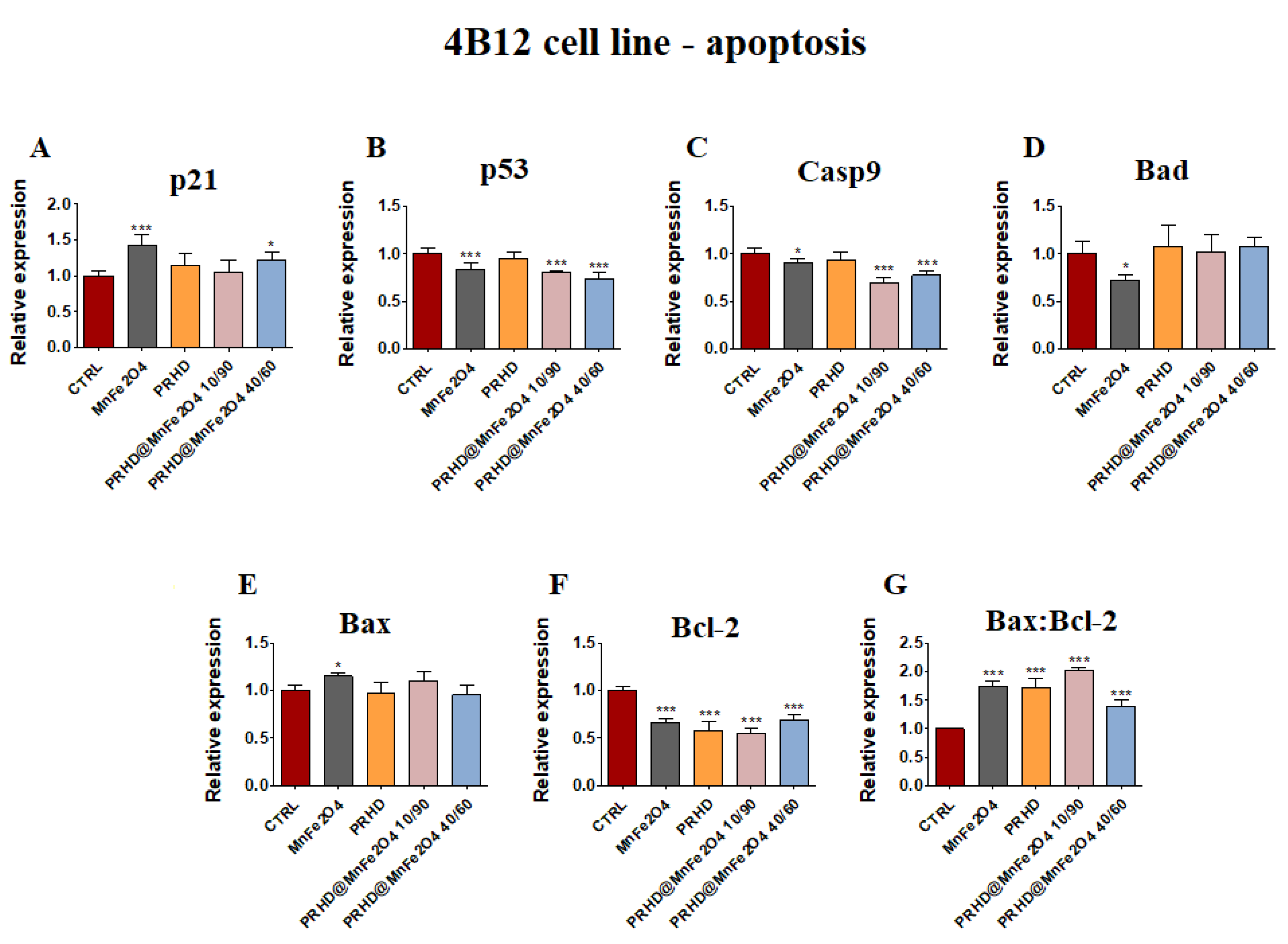
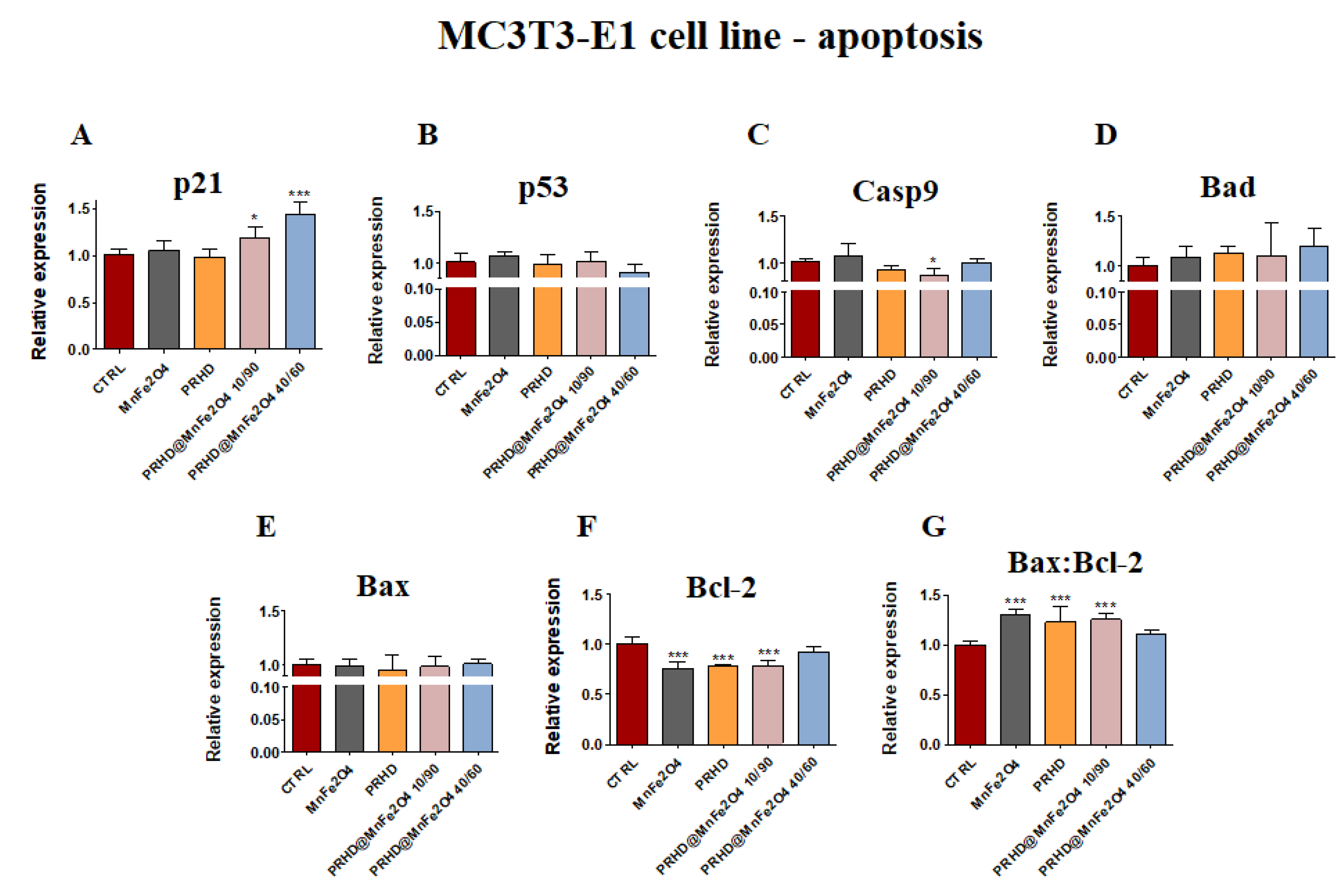
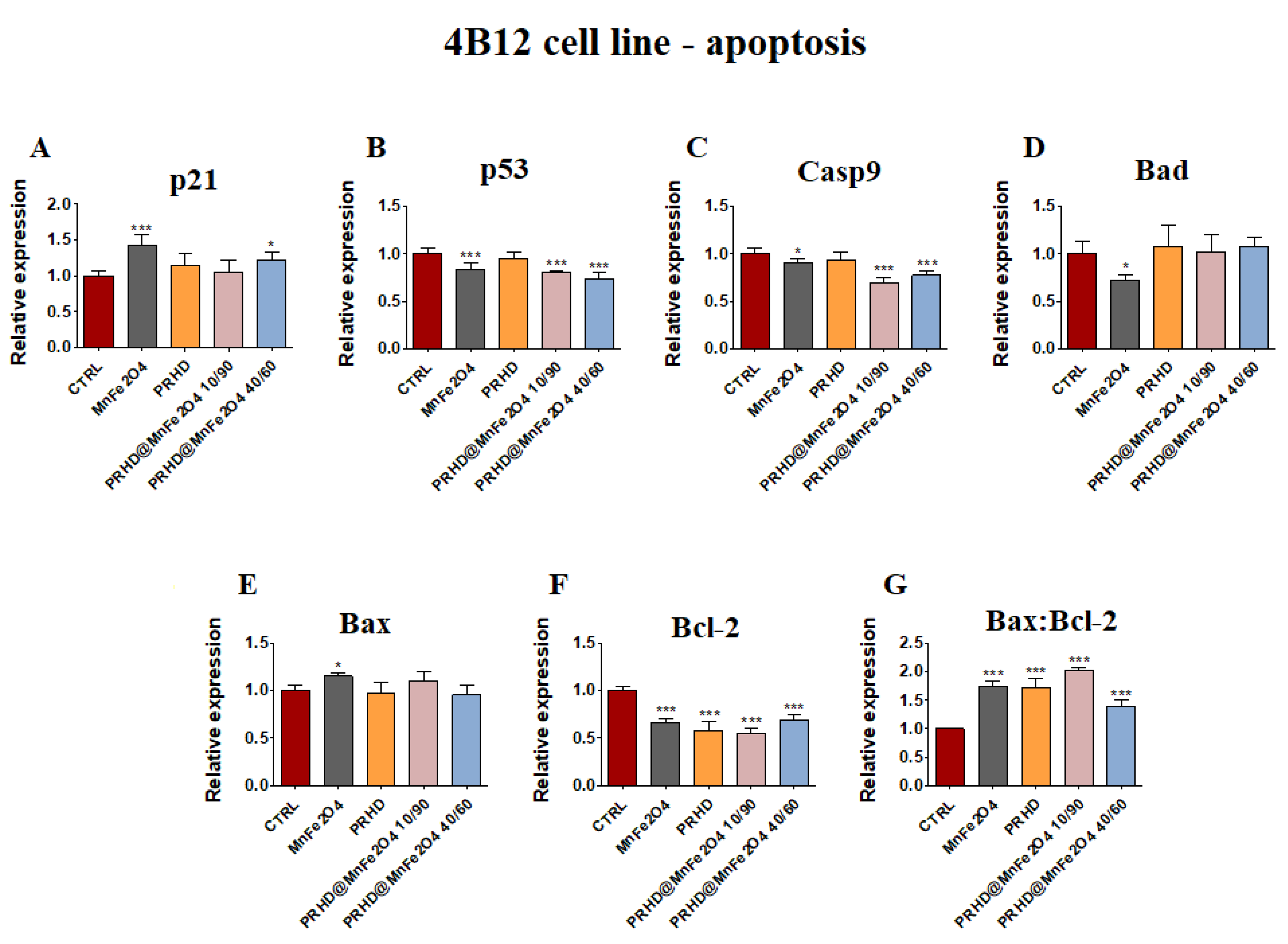
3.4.1. Genes Associated with the Apoptosis
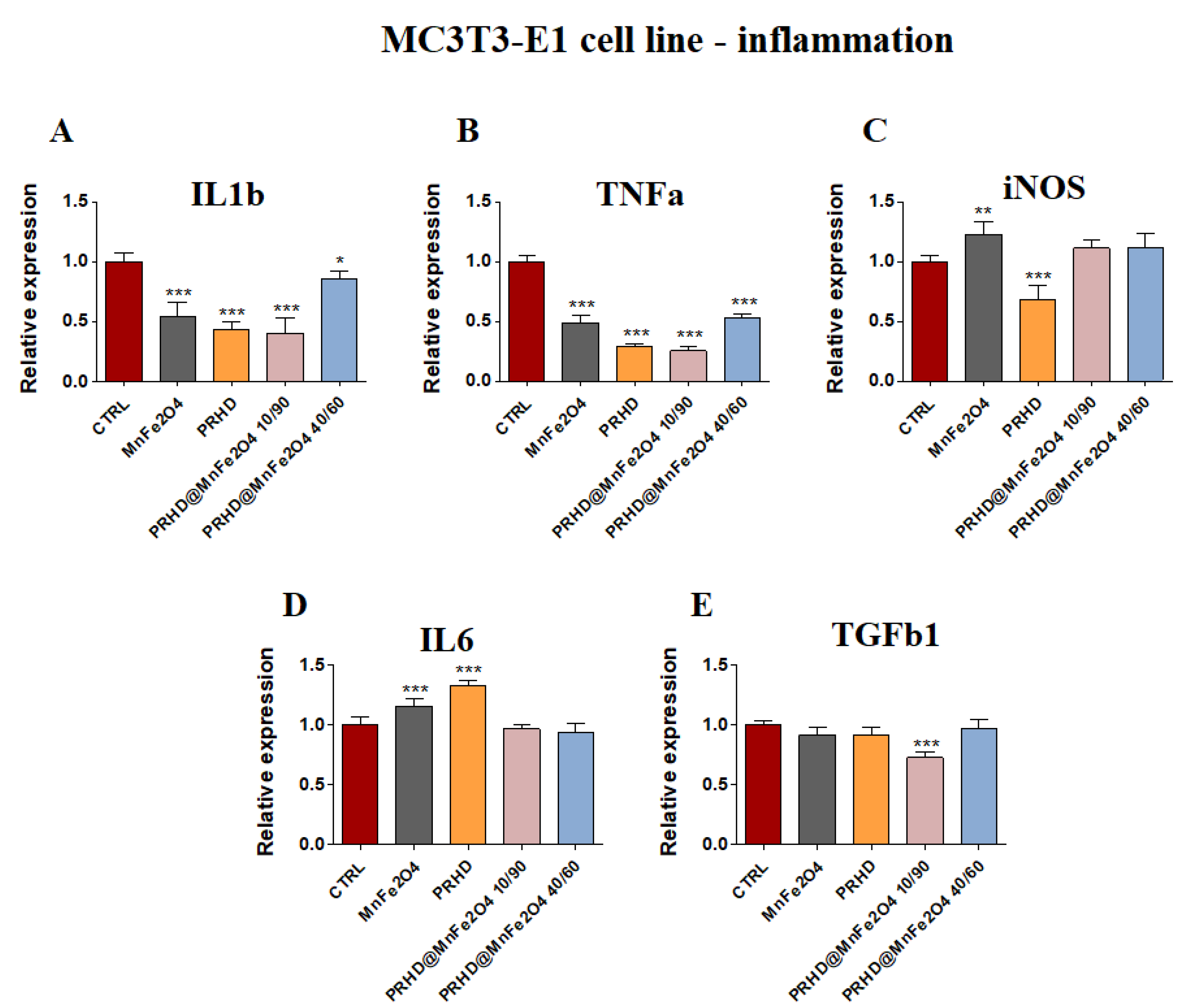
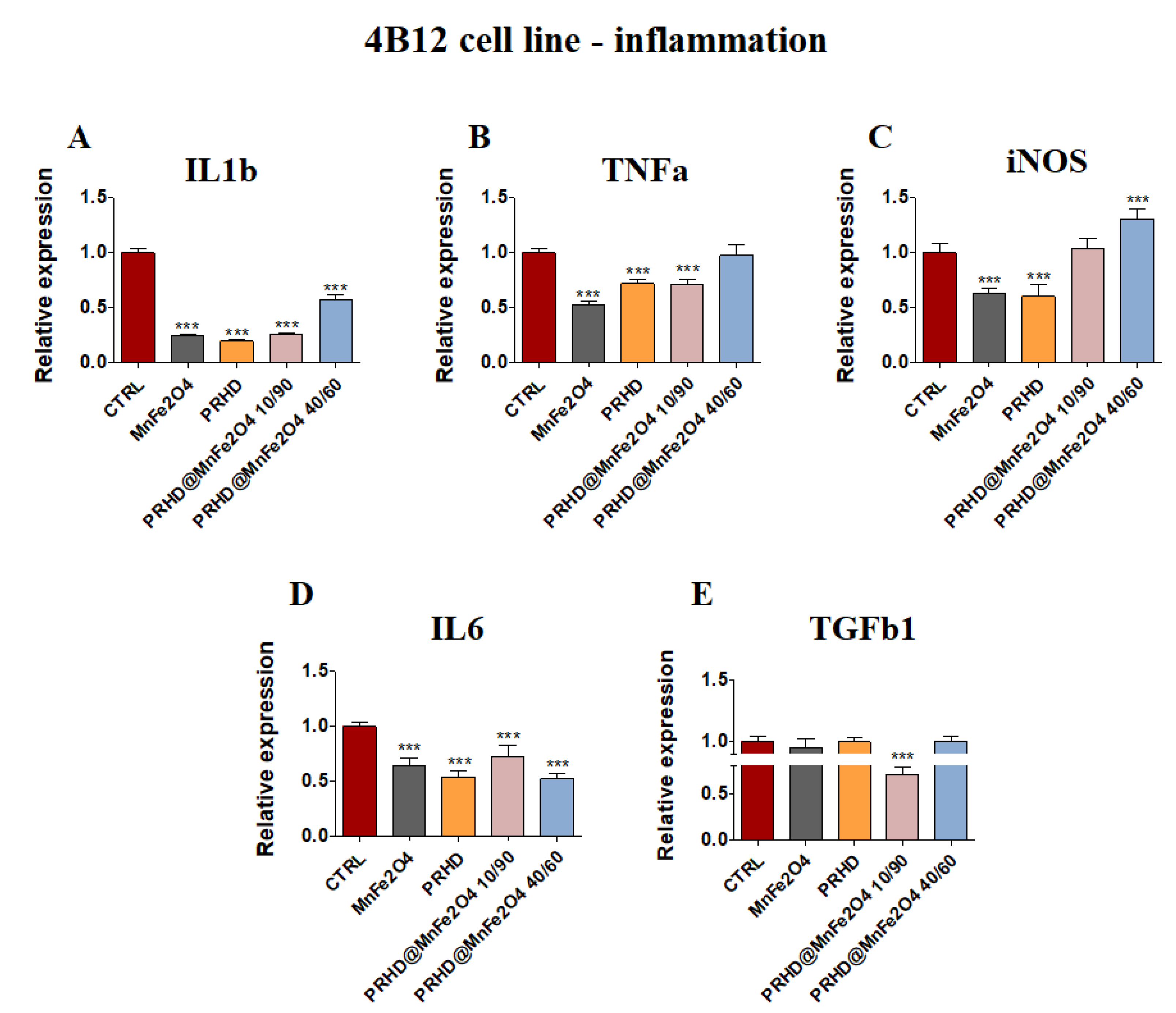
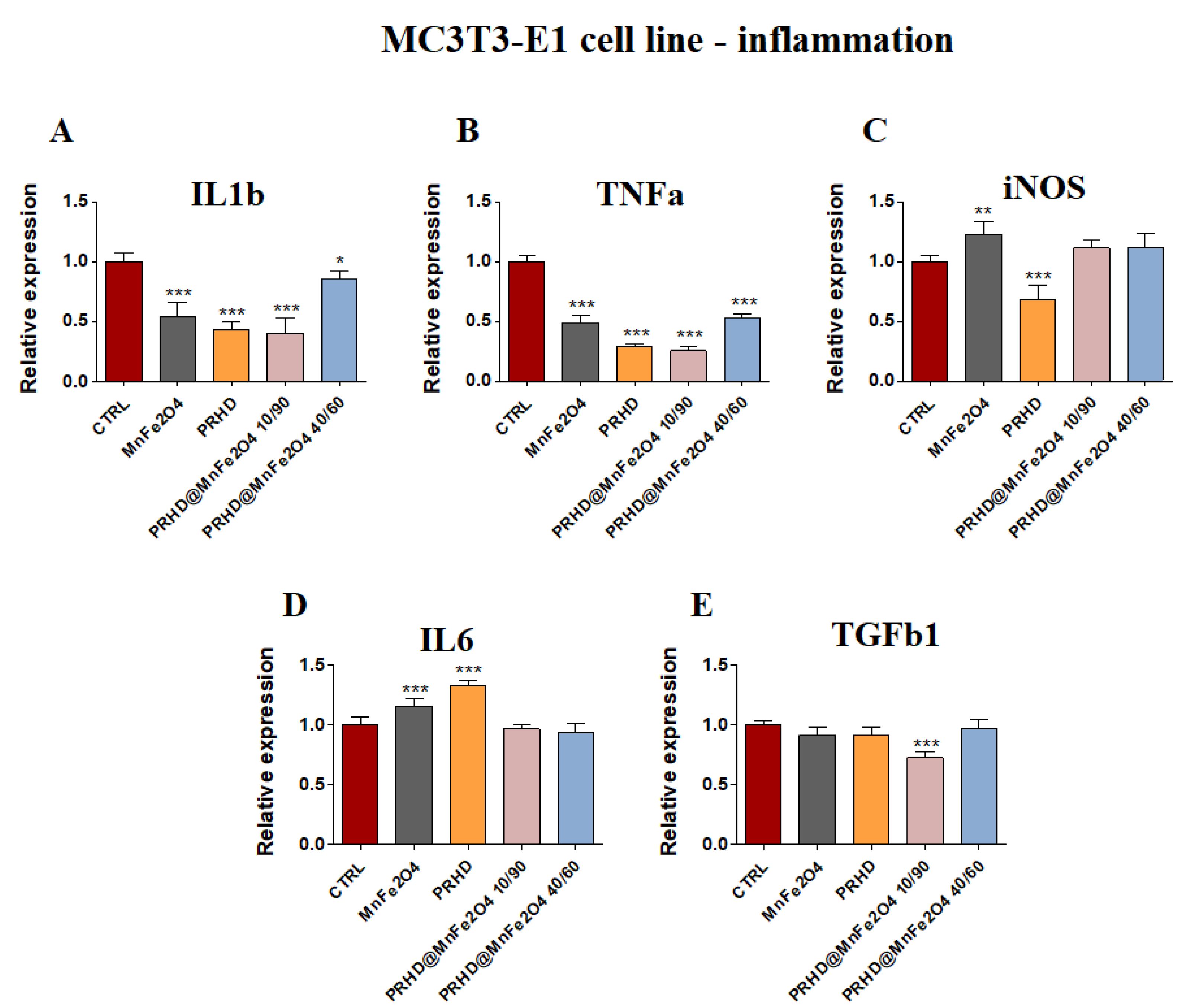
3.4.2. Genes Associated with the Inflammation
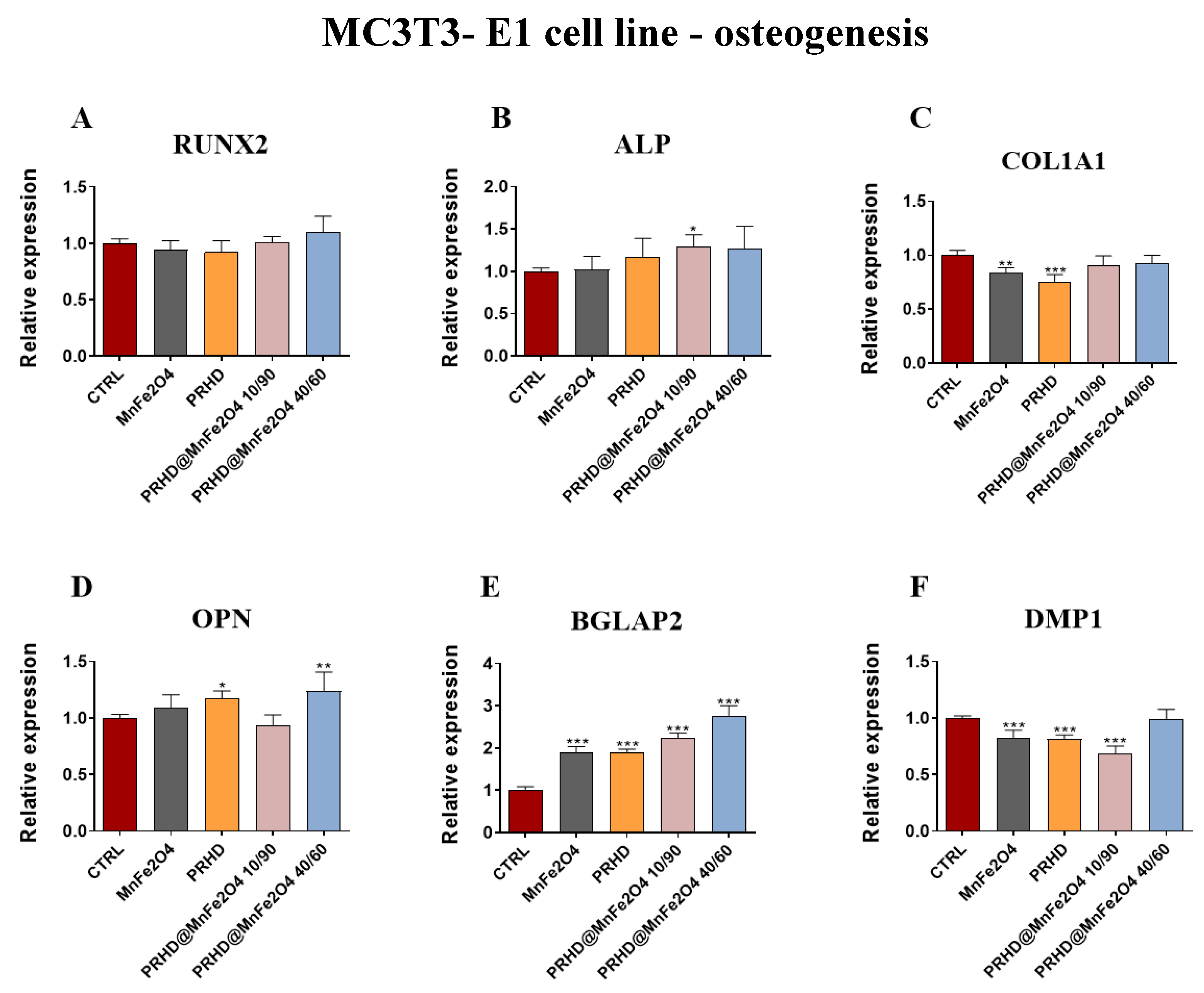
3.4.3. Genes Associated with the Osteogenic Differentiation
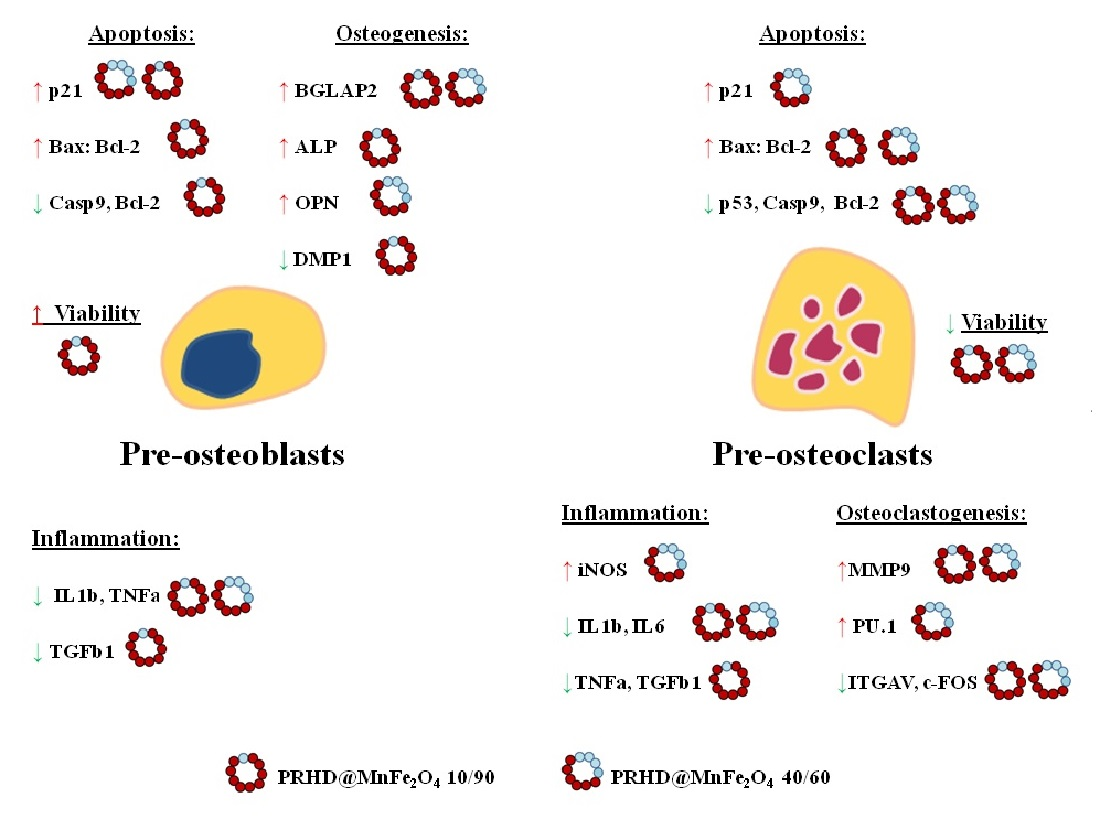
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASCs | adipose stem progenitor cells |
| ATCC | American Type Cell Culture |
| ALP | alkaline phosphatase |
| ATR | attenuated total reflection |
| BAD | Bcl-2 associated agonist of cell death |
| BAX | Bcl-2 associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| BGLAP | bone gamma-carboxyglutamic acid-containing protein (OC) |
| BMD | bone mass |
| BMSCs | bone marrow-derived mesenchymal stem cell progenitor cells |
| CASP | caspase |
| COL1A1 | collagen alpha-1 (I) chain precursor |
| CSCM | 30% calvaria-derived stromal cell conditioned media |
| DEPC | sterile filter water treated with diethyl pyrocarbonate |
| DMP1 | dentin matrix acidic phosphoprotein 1 |
| ECM | extracellular matrix |
| EVs | extracellular vesicles |
| FTIR | Fourier transform infrared spectroscopy |
| FBS | fetal bovine serum |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| IL | interleukin |
| ITGAV | integrin subunit alphaV |
| iNOS | inducible nitric oxide synthase |
| MEMα | minimum essential medium α |
| MMP | matrix metalloproteinase |
| OC | osteocalcin (BGLAP) |
| OPN | osteopontin |
| OS | osteoporosis |
| p21 | cyclin-dependent kinase inhibitor 1 |
| p53 | tumor suppressor factor |
| PU.1 | protein in human encoded by the SPI1 gene |
| PRHD | polyrhodanine |
| PRHD@MnFe2O4 | polyrhodanine manganese ferrite binary nanohybrids |
| RUNX2 | runt-related transcription factor 2 |
| SEM | scanning electron microscope |
| TEM | transmission electron microscope |
| TGA | polymer layer thermogravimetric analysis |
| TGFβ | transforming growth factor β |
| TNFRSF | tumor necrosis factor receptor superfamily |
| TRAIL | tumor necrosis factor-related apoptosis-inducing ligand |
| XRD | X-ray powder diffraction technique |
References
- Srivastava, M.; Cao, J.; Xie, C. Osteoporosis in elderly: Prevention and treatment. Clin. Geriatr. Med. 2002, 18, 529. [Google Scholar] [CrossRef]
- Sózen, T.; Ozisik, L.; Basaran, N.C. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 529. [Google Scholar] [CrossRef] [PubMed]
- Chrischilles, E.; Shireman, T.; Wallace, R. Costs and Health Effects of Osteoporotic Fractures. Bone 1994, 15, 377. [Google Scholar] [CrossRef]
- Kalervo, H.V.; Pirkko, L.H. Estrogen anti-bone metabolism. Maturitas 1996, S65, 5122. [Google Scholar]
- Henriksen, K.; Bollerslev, J.; Everts, V.; Karsdal, M.A. Osteoblast Activity and subtypes as a function of physiology and pathology—Implications for future. Endocr. Rev. 2011, 32, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; He, C. TNF-α and IL-6: The Link between Immune and Bone System. Curr. Drug Targets 2019, 21, 213. [Google Scholar] [CrossRef]
- Xu, F. The Roles of Epigenetics Regulation in Bone Metabolism and Osteoporosis. Front. Cell Dev. Biol. 2021, 8, 1928. [Google Scholar] [CrossRef]
- Zachanowicz, E.; Kulpa-Greszta, M.; Tomaszewska, A.; Gazińska, M.; Marędziak, M.; Marycz, K.; Pązik, R. Multifunctional Properties of Binary Polyrhodanine Manganese Ferrite Nanohybrids—From the Energy Converters to Biological Activity. Polymers 2020, 12, 2934. [Google Scholar] [CrossRef]
- Marycz, K.; Grzesiak, J.; Wrzeszcz, K.; Golonka, P. Adipose stem cell combined with plasma-based implant bone tissue differentiation in vitro and in a horse with a phalanx digitalis distalis fracture: A case report. Vet. Med. 2012, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Pązik, R.; Piasecka, E.; Małecka, M.; Kessler, V.D.; Idzikowski, B.; Śniadecki, Z.; Wiglusz, R. Facile non-hydrolytic synthesis of highly water dispersible, surfactant free nanoparticles of synthetic MFe2O4 (M-Mn2+, Fe2+, Co2+, Ni2+) ferrite spinel by a modified Bradley reaction. RSC Adv. 2013, 30, 3. [Google Scholar] [CrossRef]
- Amano, S.; Sekine, K.; Bonewald, L.; Ohmori, Y. A Novel Osteoblast Precursor Cell Line, 4B12, Recapitulates the Features of Primary Osteoblast Differentation and Function: Enhanced Transfection Efficiency before and after Differentation. J. Cell. Physiol. 2009, 221, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alicka, M.; Major, R.; Wysocki, M.; Marycz, K. Clinical Medicine Adipose-Derived Mesenchymal Stem Cells Isolated from Patients with Type 2 Diabetes Show Reduced “Stemness” through an Altered Secretome Profile, Impaired Anti-Oxidative Protection, and Mitochondrial Dynamics Deterioration. J. Clin. Med. 2019, 8, 765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomczyński, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 2, 581. [Google Scholar] [CrossRef] [PubMed]
- Kardas, G.; Solmaz, R. Electrochemical synthesis and characterization of a new conducting polymer: Polyrhodanine. Appl. Surf. Sci. 2007, 253, 3402. [Google Scholar] [CrossRef]
- Bhatti, R.S.; Shah, S.; Suresh; Krishan, P.; Sandhu, J.S. Recent pharmacological developments on rhodanines and 2,4-thiazolidinediones. Int. J. Med. Chem. 2013, 2013, 793260. [Google Scholar] [CrossRef] [Green Version]
- Abussetta, A.; Alumairi, J.; Alkaabi, M.Y.; Al Ajeil, R.; Shaidim, A.A.; Akram, D.; Pajak, J.; Ghattas, M.A.; Atatresh, N.; AlNeyadi, S.S. Design, Synthesis, in Vitro antibacterial activity, and docking studies of new rhodanine derivatives. Open J. Med. Chem. 2020, 10, 15–34. [Google Scholar]
- Makridis, A.; Topouridou, K.; Tziomaki, M.; Sakellari, D.; Simeonidis, K.; Angelakeris, M.; Yavropoulou, M.P.; Yovos, J.G.; Kalogirou, O. in Vitro application of Mn-ferrite nanoparticles as novel magnetic hyperthermia agents. J. Mater. Chem. B 2014, 47, 8390. [Google Scholar] [CrossRef]
- Redlich, K.; Smolen, J.S. Inflammatory bone loss: Pathogenesis and therapeutic intervention. Nat. Rev. Drug Discov. 2012, 11, 234. [Google Scholar] [CrossRef]
- Manolagasid, S.C. Osteocalcin promotes bone mineralization but is not a hormone. PLoS Genet. 2020, 16, e1008714. [Google Scholar]
- Wennberg, C.; Hessle, L.; Lundberg, P.; Mauro, S.; Narisawa, S.; Lerner, U.H.; Millan, J.L. Functional characterization of osteoblasts and osteoclasts from alkaline phosphatase knockout mice. J. Bone Miner. Res. 2000, 15, 1879. [Google Scholar] [CrossRef]
- Kwak, J.S.; Lee, Y.; Yang, J.; Kim, S.K.; Shin, Y.; Kim, H.J.; Choi, J.H.; Im, Y.J.; Kim, M.J.; Lee, Y.K.; et al. Characterization of rhodamine derivatives as potential disease-modifying drugs for experimental mouse osteoarthritis. Osteoarthr. Cartil. 2022, 14, 1. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ → 3′) | Reverse (5′ → 3′) | Length of Amplicon |
|---|---|---|---|
| p21 | TGTTCCACACAGGAGCAAAG | AACACGCTCCCAGACGTAGT | 175 |
| p53 | AGTCACAGCACATGACGGAGG | GGAGTCTTCCAGTGTGATGATGG | 287 |
| Casp9 | CCGGTGGACATTGGTTCTGG | GCCATCTCCATCAAAGCCGT | 278 |
| Bad | ACATTCATCAGCAGGGACGG | ATCCCTTCATCCTCCTCGGT | 115 |
| Bax | AGGACGCATCCACCAAGAAGC | GGTTCTGATCAGCTCGGGCA | 251 |
| Bcl-2 | GGATCCAGGATAACGGAGGC | ATGCACCCAGAGTGATGCAG | 141 |
| Runx2 | TCCGAAATGCCTCTGCTGTT | GCCACTTGGGGAGGATTTGT | 130 |
| Alp | TTCATAAGCAGGCGGGGGAG | TGAGATTCGTCCCTCGCTGG | 198 |
| Col1A1 | CCAGCCGCAAAGAGTCTACA | CAGGTTTCCACGTCTCACCA | 175 |
| Opn | AGACCATGCAGAGAGCGAG | GCCCTTTCCGTTGTTGTCCT | 340 |
| Bglap2 | CTCCTGAGAGTCTGACAAAGCCTT | GCTGTGACATCCATTACTTGC | 320 |
| Dmp1 | CCCAGAGGCACAGGCAAATA | TCCTCCCCAATGTCCTTCTT | 211 |
| Mmp9 | TTGCCCCTACTGGAAGGTATTAT | GAGAATCTCTGAGCAATCCTTGA | 172 |
| Pu.1 | GAGAAGCTGATGGCTTGGAG | TTGTGCTTGGACGAGAACTG | 175 |
| Itgav | ACAATGTAAGCCCAGTTGTGTCT | TTTGTAAGGCCACTGGAGATTTA | 236 |
| c-fos | CCAGTCAAGAGCATCAGCAA | TAAGTAGTGCAGCCCGGAGT | 248 |
| Il1b | TGCCACCTTTTGACAGTGATG | TGATGTGCTGCTGCGAGATT | 138 |
| Tnfa | ACAGAAAGCATGATCCGCGA | CTTGGTGGTTTGCTACGACG | 295 |
| iNos | GACAAGCTGCATGTGACATC | GCTGGTAGGTTCCTGTTGTT | 325 |
| Il6 | GAGGATACCACTCCCAACAGACC | AAGTGCATCATCGTTGTTCATACA | 141 |
| Tgfb1 | GGAGAGCCCTGGATACCAAC | CAACCCAGGTCCTTCCTAAA | 94 |
| Gapdh | TGCACCACCAACTGCTTAG | GGATGCAGGGATGATGTTC | 177 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marycz, K.; Kowalczuk, A.; Turlej, E.; Zachanowicz, E.; Tomaszewska, A.; Kulpa-Greszta, M.; Kępska, M.; Steczkiewicz, Z.; Pązik, R. Impact of Polyrhodanine Manganese Ferrite Binary Nanohybrids (PRHD@MnFe2O4) on Osteoblasts and Osteoclasts Activities—A Key Factor in Osteoporosis Treatment. Materials 2022, 15, 3990. https://doi.org/10.3390/ma15113990
Marycz K, Kowalczuk A, Turlej E, Zachanowicz E, Tomaszewska A, Kulpa-Greszta M, Kępska M, Steczkiewicz Z, Pązik R. Impact of Polyrhodanine Manganese Ferrite Binary Nanohybrids (PRHD@MnFe2O4) on Osteoblasts and Osteoclasts Activities—A Key Factor in Osteoporosis Treatment. Materials. 2022; 15(11):3990. https://doi.org/10.3390/ma15113990
Chicago/Turabian StyleMarycz, Krzysztof, Anna Kowalczuk, Eliza Turlej, Emilia Zachanowicz, Anna Tomaszewska, Magdalena Kulpa-Greszta, Martyna Kępska, Zofia Steczkiewicz, and Robert Pązik. 2022. "Impact of Polyrhodanine Manganese Ferrite Binary Nanohybrids (PRHD@MnFe2O4) on Osteoblasts and Osteoclasts Activities—A Key Factor in Osteoporosis Treatment" Materials 15, no. 11: 3990. https://doi.org/10.3390/ma15113990
APA StyleMarycz, K., Kowalczuk, A., Turlej, E., Zachanowicz, E., Tomaszewska, A., Kulpa-Greszta, M., Kępska, M., Steczkiewicz, Z., & Pązik, R. (2022). Impact of Polyrhodanine Manganese Ferrite Binary Nanohybrids (PRHD@MnFe2O4) on Osteoblasts and Osteoclasts Activities—A Key Factor in Osteoporosis Treatment. Materials, 15(11), 3990. https://doi.org/10.3390/ma15113990