
Enhanced Cellular Cryopreservation by Biopolymer-Associated Suppression of RhoA/ROCK Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cryopreservation
2.4. Trypan Blue Exclusion Assay
2.5. MTT Assay
2.6. Splat Assay
2.7. Quantitative Real-Time PCR Analysis
2.8. Western Blotting
2.9. Statistical Analysis
3. Results and Discussion
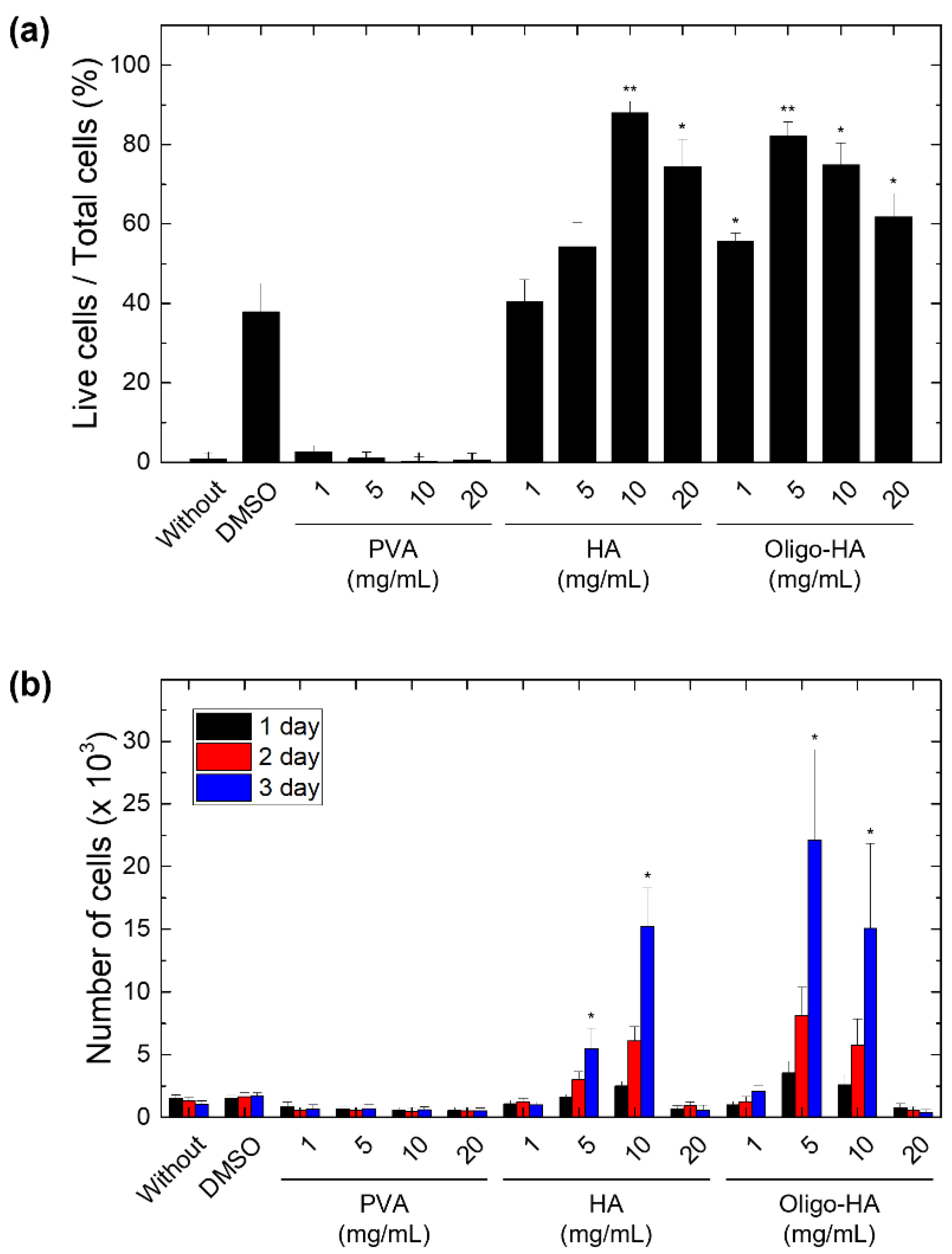
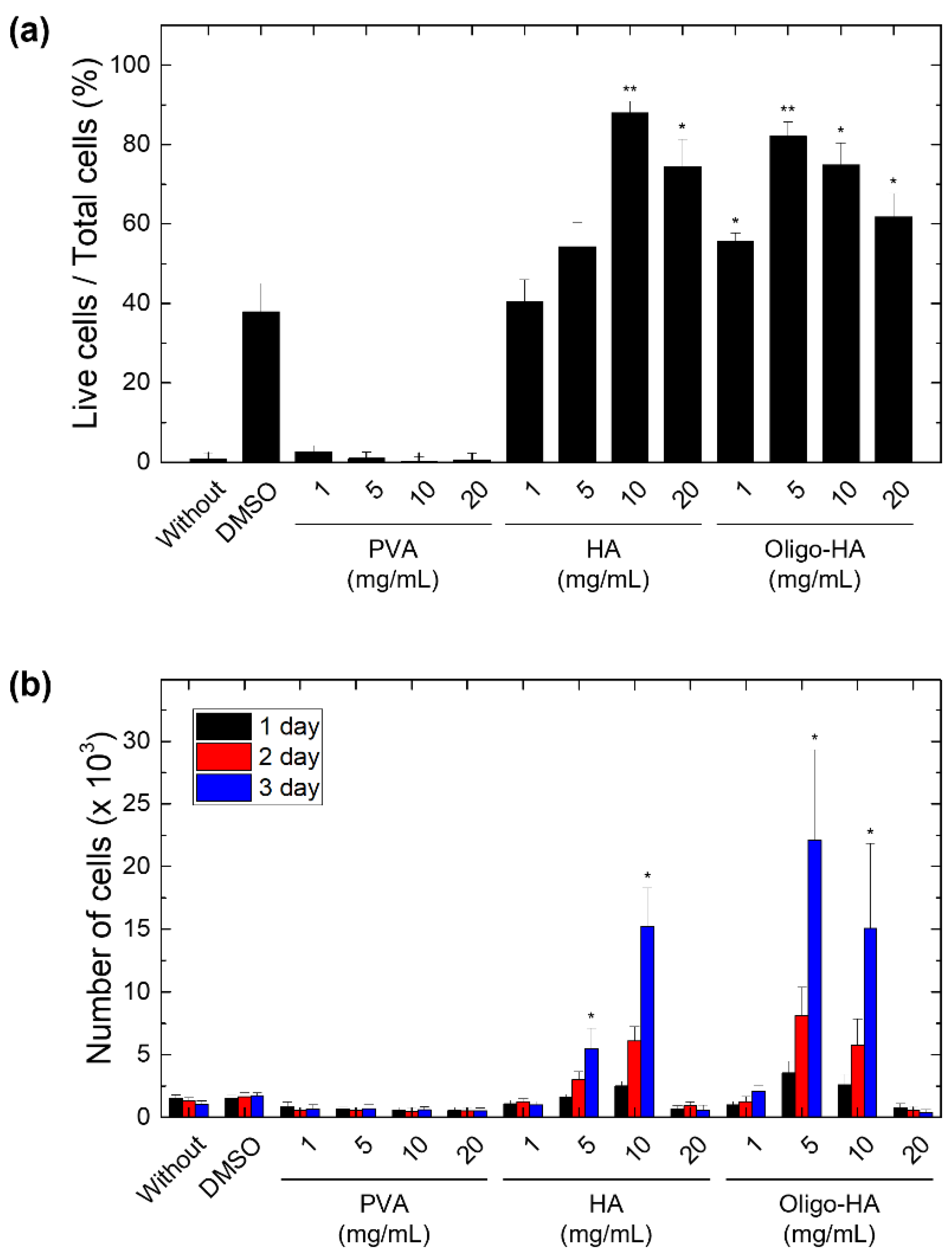
3.1. Effect of Polymeric Supplements on Cell Cryopreservation
3.2. Ice Recrystallization Inhibition Activity of Polymeric Supplements
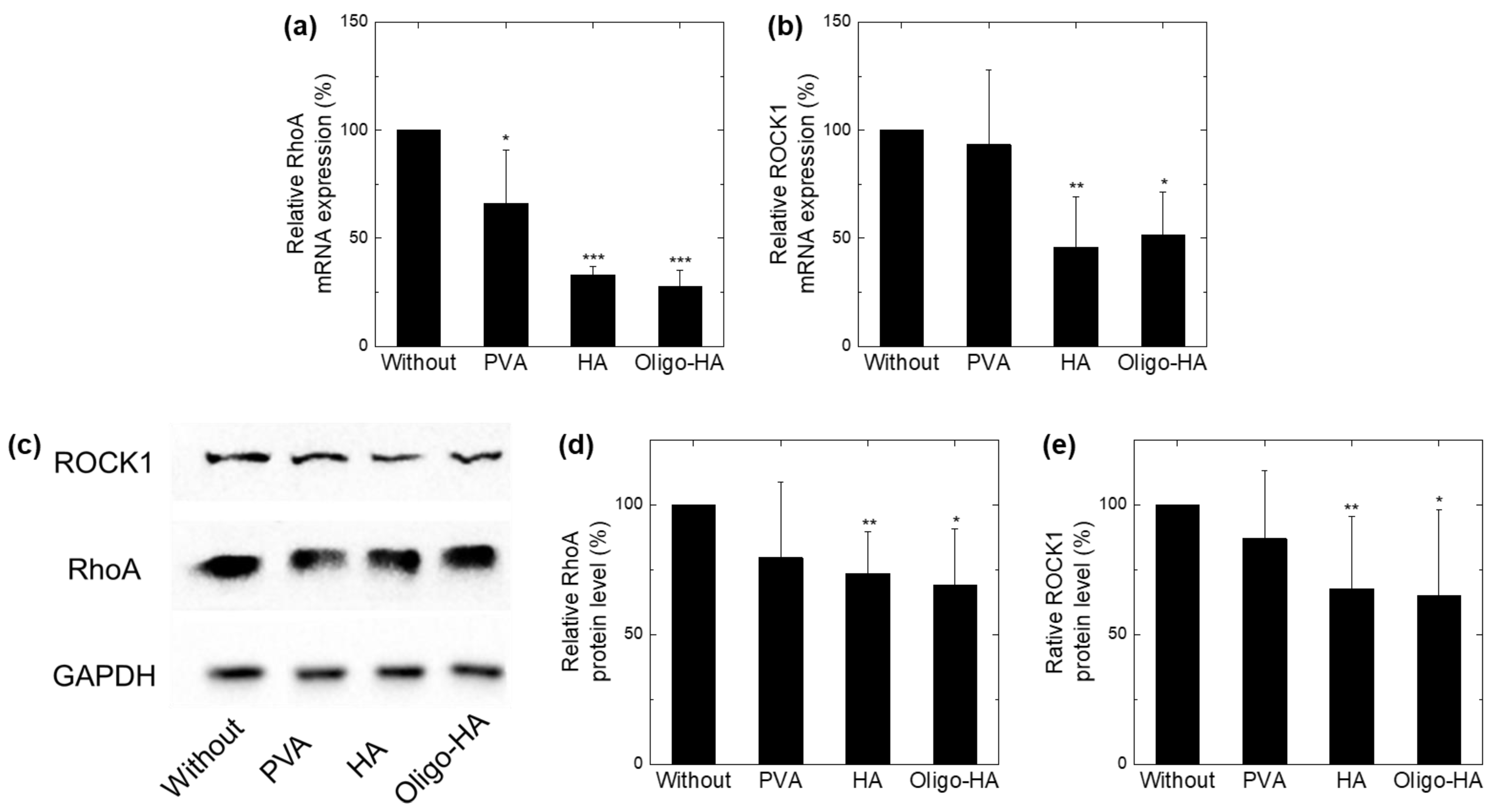
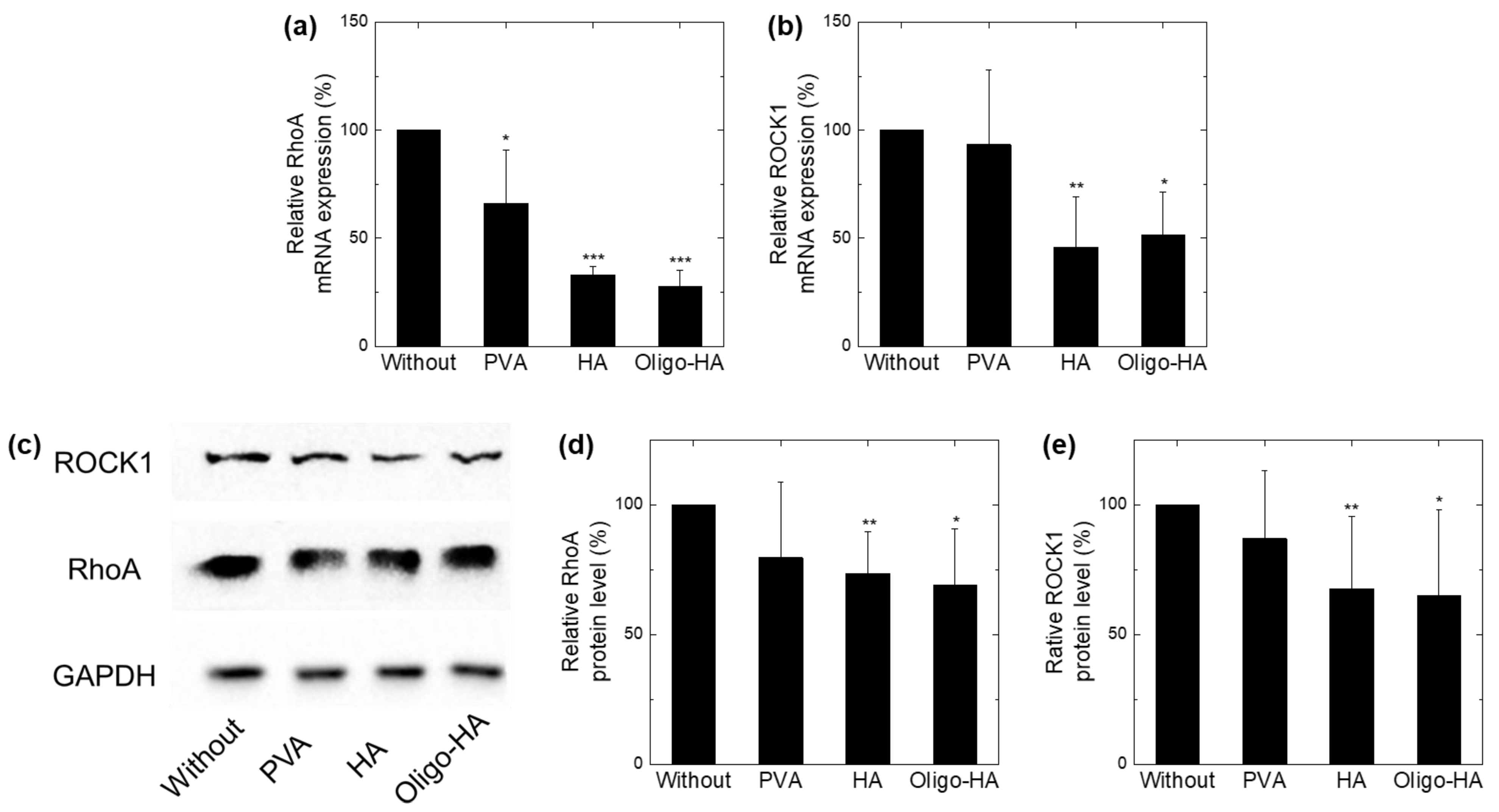
3.3. Downregulation of RhoA/ROCK1 following Polymer Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fuggle, N.R.; Cooper, C.; Oreffo, R.O.C.; Price, A.J.; Kaux, J.F.; Maheu, E.; Cutolo, M.; Honvo, G.; Conaghan, P.G.; Berenbaum, F.; et al. Alternative and complementary therapies in osteoarthritis and cartilage repair. Aging Clin. Exp. Res. 2020, 32, 547–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Yu, S.; Lin, H.; Bi, R. Expression and function of cartilage-derived pluripotent cells in joint development and repair. Stem Cell Res. Ther. 2020, 11, 111. [Google Scholar] [CrossRef]
- Willing, A.E.; Das, M.; Howell, M.; Mohapatra, S.S.; Mohapatra, S. Potential of mesenchymal stem cells alone, or in combination, to treat traumatic brain injury. CNS Neurosci. Ther. 2020, 26, 616–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, D.T.; Nguyen, T.T.; Tien, N.L.B.; Tran, D.K.; Jeong, J.H.; Anh, P.G.; Thanh, V.V.; Truong, D.T.; Dinh, T.C. Recent Progress of Stem Cell Therapy in Cancer Treatment: Molecular Mechanisms and Potential Applications. Cells 2020, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.F.; Cengiz, I.F.; Silva, F.S.; Reis, R.L.; Oliveira, J.M. Scaffolds and coatings for bone regeneration. J. Mater. Sci. Mater. Med. 2020, 31, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serowoky, M.A.; Arata, C.E.; Crump, J.G.; Mariani, F.V. Skeletal stem cells: Insights into maintaining and regenerating the skeleton. Development 2020, 147, dev179325. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, H.; Kusumoto, D.; Hashimoto, H.; Yuasa, S. Stem Cell Aging in Skeletal Muscle Regeneration and Disease. Int. J. Mol. Sci. 2020, 21, 1830. [Google Scholar] [CrossRef] [Green Version]
- Mossahebi-Mohammadi, M.; Quan, M.; Zhang, J.S.; Li, X. FGF Signaling Pathway: A Key Regulator of Stem Cell Pluripotency. Front. Cell Dev. Biol. 2020, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Coloma, L.; Cabello, R.; Gonzalez, C.; Quicios, C.; Bueno, G.; Garcia, J.V.; Arribas, A.B.; Clasca, F. Cadaveric Models for Renal Transplant Surgery Education: A Comprehensive Review. Curr. Urol. Rep. 2020, 21, 10. [Google Scholar] [CrossRef]
- Tricot, T.; De Boeck, J.; Verfaillie, C. Alternative Cell Sources for Liver Parenchyma Repopulation: Where Do We Stand? Cells 2020, 9, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhou, Y.; Zhang, L.; Wu, L.; Chen, Y.; Xie, D.; Chen, W. Hydrogel Cryopreservation System: An Effective Method for Cell Storage. Int. J. Mol. Sci. 2018, 19, 3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhou, R.; Li, L.; Peters, B.M.; Li, B.; Lin, C.W.; Chuang, T.L.; Chen, D.; Zhao, X.; Xiong, Z.; et al. Viable but non-culturable state and toxin gene expression of enterohemorrhagic Escherichia coli O157 under cryopreservation. Res. Microbiol. 2017, 168, 188–193. [Google Scholar] [CrossRef]
- Polge, C.; Smith, A.U.; Parkes, A.S. Revival of spermatozoa after vitrification and dehydration at low temperatures. Nature 1949, 164, 666. [Google Scholar] [CrossRef]
- Iussig, B.; Maggiulli, R.; Fabozzi, G.; Bertelle, S.; Vaiarelli, A.; Cimadomo, D.; Ubaldi, F.M.; Rienzi, L. A brief history of oocyte cryopreservation: Arguments and facts. Acta Obstet. Gynecol. Scand. 2019, 98, 550–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovelock, J.; Bishop, M. Prevention of freezing damage to living cells by dimethyl sulphoxide. Nature 1959, 183, 1394–1395. [Google Scholar] [CrossRef]
- Capicciotti, C.J.; Doshi, M.; Ben, R.N. Ice recrystallization inhibitors: From biological antifreezes to small molecules. Recent Dev. Study Recryst. 2013, 177, 224. [Google Scholar]
- Ruiz-Delgado, G.J.; Mancias-Guerra, C.; Tamez-Gomez, E.L.; Rodriguez-Romo, L.N.; Lopez-Otero, A.; Hernandez-Arizpe, A.; Gomez-Almaguer, D.; Ruiz-Arguelles, G.J. Dimethyl sulfoxide-induced toxicity in cord blood stem cell transplantation: Report of three cases and review of the literature. Acta Haematol. 2009, 122, 1–5. [Google Scholar] [CrossRef]
- Yong, K.W.; Pingguan-Murphy, B.; Xu, F.; Abas, W.A.; Choi, J.R.; Omar, S.Z.; Azmi, M.A.; Chua, K.H.; Wan Safwani, W.K. Phenotypic and functional characterization of long-term cryopreserved human adipose-derived stem cells. Sci. Rep. 2015, 5, 9596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivakumar, S.B.; Bharti, D.; Subbarao, R.B.; Jang, S.J.; Park, J.S.; Ullah, I.; Park, J.K.; Byun, J.H.; Park, B.W.; Rho, G.J. DMSO- and Serum-Free Cryopreservation of Wharton’s Jelly Tissue Isolated From Human Umbilical Cord. J. Cell. Biochem. 2016, 117, 2397–2412. [Google Scholar] [CrossRef]
- Arutyunyan, I.V.; Strokova, S.O.; Makarov, A.V.; Mullabaeva, S.M.; Elchaninov, A.V.; Lokhonina, A.V.; Abramov, A.A.; Fatkhudinov, T.K. DMSO-Free Cryopreservation of Human Umbilical Cord Tissue. Bull. Exp. Biol. Med. 2018, 166, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Fayter, A.E.R.; Gibson, M.I. Ice Recrystallization Inhibiting Polymers Enable Glycerol-Free Cryopreservation of Microorganisms. Biomacromolecules 2018, 19, 3371–3376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, H.; Yang, X.; Ke, L.; Hu, Y. The properties, biotechnologies, and applications of antifreeze proteins. Int. J. Biol. Macromol. 2020, 153, 661–675. [Google Scholar] [CrossRef]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A review of the actions and applications of cryoprotective solutes that modulate cell recovery from ultra-low temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef]
- Robles, V.; Valcarce, D.G.; Riesco, M.F. The Use of Antifreeze Proteins in the Cryopreservation of Gametes and Embryos. Biomolecules 2019, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biggs, C.I.; Bailey, T.L.; Ben, G.; Stubbs, C.; Fayter, A.; Gibson, M.I. Polymer mimics of biomacromolecular antifreezes. Nature Commun. 2017, 8, 1546. [Google Scholar] [CrossRef] [Green Version]
- Bari, J.; Islam, M.; Alam, M.H.; Khatun, A.; Hashem, M.; Moniruzzaman, M. Effect of Polyvinylpyrrolidone on Vitrification of Buffalo (Bubalus bubalis) Oocytes. J. Buffalo Sci. 2020, 9, 152–158. [Google Scholar] [CrossRef]
- Li, H.; Li, P.; Yang, Z.; Gao, C.; Fu, L.; Liao, Z.; Zhao, T.; Cao, F.; Chen, W.; Peng, Y.; et al. Meniscal Regenerative Scaffolds Based on Biopolymers and Polymers: Recent Status and Applications. Front. Cell Dev. Biol. 2021, 9, 661802. [Google Scholar] [CrossRef]
- Salwowska, N.M.; Bebenek, K.A.; Żądło, D.A.; Wcisło-Dziadecka, D.L. Physiochemical properties and application of hyaluronic acid: A systematic review. J. Cosmet. Dermatol. 2016, 15, 520–526. [Google Scholar] [CrossRef]
- Reddy, N.; Reddy, R.; Jiang, Q. Crosslinking biopolymers for biomedical applications. Trends Biotechnol. 2015, 33, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Uebersax, L.; Merkle, H.P.; Meinel, L. Biopolymer-based growth factor delivery for tissue repair: From natural concepts to engineered systems. Tissue Eng. Part B Rev. 2009, 15, 263–289. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, S.H.; Ma, K.; Zhao, B.Y.; Yan, B.; Pei, C.B.; Zhou, Y.; Wang, H.H.; Wang, H.Y.; Ma, L.H.; et al. Rho/ROCK signaling pathway and anti-cryodamage ability of human sperm. Zhonghua Natl. J. Androl. 2019, 25, 322–328. [Google Scholar]
- Gu, M.; Ni, H.; Sheng, X.; Pauciullo, A.; Liu, Y.; Guo, Y. RhoA phosphorylation mediated by Rho/RhoA-associated kinase pathway improves the anti-freezing potentiality of murine hatched and diapaused blastocysts. Sci. Rep. 2017, 7, 6705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebbagh, M.; Renvoizé, C.; Hamelin, J.; Riché, N.; Bertoglio, J.; Bréard, J. Caspase-3-mediated cleavage of ROCK I induces MLC phosphorylation and apoptotic membrane blebbing. Nat. Cell Biol. 2001, 3, 346–352. [Google Scholar] [CrossRef]
- Jiang, W.; Li, M.; Chen, Z.; Leong, K.W. Cell-laden microfluidic microgels for tissue regeneration. Lab Chip 2016, 16, 4482–4506. [Google Scholar] [CrossRef] [Green Version]
- Lotfi, S.; Mehri, M.; Sharafi, M.; Masoudi, R. Hyaluronic acid improves frozen-thawed sperm quality and fertility potential in rooster. Anim. Reprod. Sci. 2017, 184, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Khetan, S.; Corey, O. Maintenance of stem cell viability and differentiation potential following cryopreservation within 3-dimensional hyaluronic acid hydrogels. Cryobiology 2019, 90, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, S.; Cordoba, M. Hyaluronic acid-induced capacitation involves protein kinase C and tyrosine kinase activity modulation with a lower oxidative metabolism in cryopreserved bull sperm. Theriogenology 2018, 122, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Sbracia, M.; Grasso, J.; Sayme, N.; Stronk, J.; Huszar, G. Hyaluronic acid substantially increases the retention of motility in cryopreserved/thawed human spermatozoa. Hum. Reprod. 1997, 12, 1949–1954. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.A.; Mendel, G.; Wauthier, E.; Barbier, C.; Reid, L.M. Hyaluronan-supplemented buffers preserve adhesion mechanisms facilitating cryopreservation of human hepatic stem/progenitor cells. Cell Transplant. 2012, 21, 2257–2266. [Google Scholar] [CrossRef]
- Liu, X.; Pan, Y.; Liu, F.; He, Y.; Zhu, Q.; Liu, Z.; Zhan, X.; Tan, S. A Review of the Material Characteristics, Antifreeze Mechanisms, and Applications of Cryoprotectants (CPAs). J. Nanomater. 2021. [Google Scholar] [CrossRef]
- Vail, N.S.; Stubbs, C.; Biggs, C.I.; Gibson, M.I. Ultralow dispersity poly (vinyl alcohol) reveals significant dispersity effects on ice recrystallization inhibition activity. ACS Macro Lett. 2017, 6, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Bodo, M.; Pezzetti, F.; Baroni, T.; Carinci, F.; Arena, N.; Nicoletti, I.; Becchetti, E. Hyaluronic acid modulates growth, morphology and cytoskeleton in embryonic chick skin fibroblasts. Int. J. Dev. Biol. 2002, 37, 349–352. [Google Scholar]
- Harvima, I.T.; Heikura, H.; Hyttinen, M.; Naukkarinen, A. Hyaluronic acid inhibits the adherence and growth of monolayer keratinocytes but does not affect the growth of keratinocyte epithelium. Arch. Dermatol. Res. 2006, 298, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Boeckel, D.G.; Shinkai, R.S.; Grossi, M.L.; Teixeira, E.R. In vitro evaluation of cytotoxicity of hyaluronic acid as an extracellular matrix on OFCOL II cells by the MTT assay. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2014, 117, e423–e428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, A.; Toner, M. Cryo-injury and biopreservation. Ann. N. Y. Acad. Sci. 2005, 1066, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Muldrew, K.; McGann, L.E. Mechanisms of intracellular ice formation. Biophys. J. 1990, 57, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Mazur, P. Life in the Frozen State; CRC Press: Boca Raton, FL, USA, 2004; pp. 3–65. [Google Scholar]
- Misra, S.; Toole, B.P.; Ghatak, S. Hyaluronan constitutively regulates activation of multiple receptor tyrosine kinases in epithelial and carcinoma cells. J. Biol. Chem. 2006, 281, 34936–34941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, S.; Ghatak, S.; Toole, B.P. Regulation of MDR1 expression and drug resistance by a positive feedback loop involving hyaluronan, phosphoinositide 3-kinase, and ErbB2. J. Biol. Chem. 2005, 280, 20310–20315. [Google Scholar] [CrossRef] [Green Version]
- Misra, S.; Obeid, L.M.; Hannun, Y.A.; Minamisawa, S.; Berger, F.G.; Markwald, R.R.; Toole, B.P.; Ghatak, S. Hyaluronan constitutively regulates activation of COX-2-mediated cell survival activity in intestinal epithelial and colon carcinoma cells. J. Biol. Chem. 2008, 283, 14335–14344. [Google Scholar] [CrossRef] [Green Version]
- Misra, S.; Hascall, V.C.; Markwald, R.R.; Ghatak, S. Interactions between hyaluronan and its receptors (CD44, RHAMM) regulate the activities of inflammation and cancer. Front. Immunol. 2015, 6, 201. [Google Scholar] [CrossRef] [Green Version]
- Razinia, Z.; Castagnino, P.; Xu, T.; Vázquez-Salgado, A.; Puré, E.; Assoian, R.K. Stiffness-dependent motility and proliferation uncoupled by deletion of CD44. Sci. Rep. 2017, 7, 16499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, P.S.; Majumdar, S.; Jacob, M.; Hayden, J.; Mrass, P.; Weninger, W.; Assoian, R.K.; Puré, E. Fibroblast migration is mediated by CD44-dependent TGFβ activation. J. Cell Sci. 2008, 121, 1393–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govindaraju, P.; Todd, L.; Shetye, S.; Monslow, J.; Puré, E. CD44-dependent inflammation, fibrogenesis, and collagenolysis regulates extracellular matrix remodeling and tensile strength during cutaneous wound healing. Matrix Biol. 2019, 75–76, 314–330. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, T.W.; Lee, G.W.; An, S.; Seong, K.-Y.; Lee, J.S.; Yang, S.Y. Enhanced Cellular Cryopreservation by Biopolymer-Associated Suppression of RhoA/ROCK Signaling Pathway. Materials 2021, 14, 6056. https://doi.org/10.3390/ma14206056
Lee TW, Lee GW, An S, Seong K-Y, Lee JS, Yang SY. Enhanced Cellular Cryopreservation by Biopolymer-Associated Suppression of RhoA/ROCK Signaling Pathway. Materials. 2021; 14(20):6056. https://doi.org/10.3390/ma14206056
Chicago/Turabian StyleLee, Tae Wook, Gyeong Won Lee, Seonyeong An, Keum-Yong Seong, Jong Soo Lee, and Seung Yun Yang. 2021. "Enhanced Cellular Cryopreservation by Biopolymer-Associated Suppression of RhoA/ROCK Signaling Pathway" Materials 14, no. 20: 6056. https://doi.org/10.3390/ma14206056
APA StyleLee, T. W., Lee, G. W., An, S., Seong, K.-Y., Lee, J. S., & Yang, S. Y. (2021). Enhanced Cellular Cryopreservation by Biopolymer-Associated Suppression of RhoA/ROCK Signaling Pathway. Materials, 14(20), 6056. https://doi.org/10.3390/ma14206056