
Developments for Collagen Hydrolysate in Biological, Biochemical, and Biomedical Domains: A Comprehensive Review



Abstract
1. Introduction
2. Species-Based Literature
2.1. Species as Collagen Source
2.1.1. Bovine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Source Organ | Subject | Type | Peptide Sequence/Amino Acid Contents | MW | Biological Function |
|---|---|---|---|---|---|---|
| Bovine/calve/yak | Calves blood extract [9] | Rabbits | Not given | Not given | Not given | Bone regeneration |
| Bone and calcium citrate [10] | Tail suspended ovariectomized rats | I | Not given | Not given | Increased serum osteocalcin levels | |
| Yak bone [12] | Ovariectomized rats | I | Not given | Not given | Bone formation biomarkers | |
| Yak bones collagen [13] | Not given | I | GPSGPAGKDGRIGQPG GDRGETGPAGPAGPIGPV | <3 KDa | Osteoblast proliferation | |
| Bovine Achilles tendon [14] | Not given | I | 44 peptides GERGFPGLPGPS | 29 peptides with <1 KDa | Angiotensin-converting enzyme (ACE)-inhibitory activity | |
| Bovine Achilles tendon [16] | Not given | I | (1) Gly-Asp-Hyl-Gly-Glu-Thr-Gly-Glu-Gln-Gly-Asp-Arg, (2) Phe-Leu-Pro-Gln-Pro-Pro-Gln-Glu-Lys-Ala-His-Asp-Gly-Gly-Arg | Several peptides <2 KDa | Antimicrobial and radical scavenging | |
| Bovine lung tissue [17] | Not given | Not clear | Not given | Not given | anti-inflammatory | |
| Bovine Nuchal ligament [15] | Not given | I | Gly-Pro-Arg-Gly-Phe (not ACE inhibited) Tyr-Trp and Leu-Arg-Tyr (ACE Inhibited) | Not given | ACE inhibitor peptides | |
| Porcine | Porcine skin [18] | Mouse lymphoma L5178YTK ± (not a mouse) | Majorly IPeptides of ii, iii and iv | Not given | <K3Da | Chronic wound healing |
| porcine leg bones [19] | Osteoblast MC3T3-E1 cells (not a mouse) | I | Not given | 1 to >5 KDa | Overcome osteoporosis effects on bones | |
| Fresh pig bone [20] | Osteoblast MC3T3-E1 cells (not a mouse) | I | Not given | Not given | Upregulating osteoblasts | |
| Pig skin gelatin water extracts [21] | Male ICR mice | Not given | Not given | <3 KDa | Cure of cognitive function | |
| Porcine gelatin [22] | Human | Not given | Not given | 1200 Da | language cognitive function and brain structure | |
| Pig skin gelatin [23] | Human neuroblastoma cell line (SH-SY5Y) | Not given | Not given | >50 KDa | Antioxidative and neuroprotective effect | |
| Marine | Pacific oyster (Crassostrea gigas) [24] | SKH hairless mice (Mice dorsal skins) | I | Ile-Val-Val-Pro-Lys | 554.72 Da | Anti-photoaging |
| Fresh water Salmo Salar skin Deep sea Tilapia Nilotica skin [25] | Rat | Not given | Asp, Glu, Ser, His, Gly, Thr, Arg, Ala, Tyr, Val, Met, Phe, Ile, Leu, Lys, Pro, Hyp | 512.63 Da for Tilapia Nilotica | Altering Cutaneous Microbiome Colonization for wound healing | |
| Redlip Croaker (Pseudosciaena polyactis) Scales [26] | Not given | I | DGPEGR, GPEGPMGLE, EGPFGPEG, YGPDGPTG, GFIGPTE, and IGPLGA | 629.6, 885.9, 788.9, 762.7, 733.8, and 526.6 Da | Anti-oxidation scavenging | |
| Soft shell of turtle (Carapax Trionycis) [27] | SD rats | Not given | Amino acid w.r.t mg/100 g Ser > Gly > Tyr > Ala > Glu > Asp > Cys > Val> Arg > Lys > Thr > Ile > Met > Leu > Try > His > Phe | Not given | Pre-biotic potential | |
| Swim bladders of Atlantic cod (Gadus morhua) [28] | Human fibro blast cell (MRC-5) | I | Not given | ACS 182.9 ± 7.9 PSC 28.7 ± 1.7 KDa | High thermal properties | |
| Mackerel fillets (Scomber scombrus) [29] | Not given | Not given | Aspartic acid (Asp), Serine (Ser), Glutamic acid (Glu), Glycine (Gly), Arginine (Arg), Threonine (Thr), Alanine (Ala), Proline (Pro), Tyrosine (Tyr), Valine (Val), Methionine (Met), Lysine (Lys), Isoleucine (Ile), Leucine (Leu), Phenylalanine (Phe) | Not given | Antioxidation and antihypertension | |
| Asian swamp eels [30] | Not given | Not given | Asp, Glu, Lys, His, Arg, Asn, Gln, Ser and Thr | Not given | Emulsifier and stabilizer in food products | |
| Sharks [31] | Mice | Not given | Asn, Thr, Ser, Glu, Pro, Gly, Ala, Val, Cys, Met, Leu,Ile, Tyr, Phe, His, Lys, Arg, Hyp. | <800 Da | Osteoblast proliferation | |
| Chicken | Sternal cartilage [32] | Not given | II | Glu, Ala, Pro, and Hyp,Cys, Met, Ile, Tyr, Phe, His | 245 KDa | Type II collagen |
| Chicken Heads (gelatin) [33] | Not given | Not given | Not given | Not given | Good use of by-product | |
| Skin (gelatin) [34] | Not given | B | Hydroxyproline | Not given | Good use of by-product | |
| Frog | Rana chensinensis skin [35] | Not given | Not given | Ala, Arg, Asp, Cys, Glu, Gly, His, Iso, Leu, Lys, Met, Phe, Hyp, Pro, Ser, Thr Tyr, Val | 106 KDa | Food stuff and medical |
| Duck | Breast [36] | Not given | Not given | Asp, Thr, Ser, Glu, Gly, Ala, Cys, Val, Met, Ile, Leu, Tyr, Phe, Lys, His, Arg, Pro | 491–715 Da | Antioxidant |
| Sheep and lambs | Bone, cartilage, carcass trimmings and meat [37] | Not given | I | Not given | 5 < MW < 100 KDa | Good use of by-product |
| Bees | Royal Jelly [38] | Drosophila | Not given | Gly, Glu, asp, Arg, Pro, Leu, Ala and Val | <1 KDa | Anti-oxidative, anti-aging |
2.1.2. Porcine
2.1.3. Marine
2.1.4. Chicken
2.1.5. Others Recent Sources
2.2. Species as Collagen Hydrolysate Subjects
3. Collagen Type
3.1. Type I
3.1.1. Aimed Biomedical Function for Type I
3.1.2. Novel Method for Type I
3.1.3. Novel Source for Type I
3.1.4. Biosynthesis and Culturing for Type I
| Source | Category | Method | Days | Purpose | Results |
|---|---|---|---|---|---|
| Chicken-foot collagen hydrolysate [53] | Novel Source and Biomedical function | Proteases (enzymatic hydrolysis) | To explain the mechanism for absorbance of low molecular weight hydrolysate | Osteoporosis is decreased and the following are increased, Organic substance, tri-peptides in plasma | |
| Emu skin [57] | Novel source | Enzymatic hydrolysis (pepsin, trypsin, papain, chymotrypsin, pronase E, thermolysine) | To make Type collagen hydrolysate with Emu skin | Better antioxidant and anti-allergic properties | |
| Gutted silver carp [56] | Novel Method and source | pH shift for extracting pure sediments only, then sequential pepsin trypsin hydrolysis | Exploring the potential of sediments from pH shift for collagen/collagen hydrolysate | High molecular weight collagen with better gel strength | |
| Bovine bone [58] | Novel source and partially Biosynthesis | Novel Ginger proteases (papain family) | MC3T3-E1 pre-osteoblasts culturing for 7 days | Explore the potential of novel proteases for osteoporosis | Novel X-Hype-Gly, 1.9-fold more MC3T3-E1 mineralization |
| Deer sinew [59] | Novel Source | Sequential Enzyme hydrolysis (pepsin then trypsin) | A novel dermal collagen hydrolysate | Oxidation decreased, Cell proliferation of NIH3T3 fibroblasts increases | |
| Patent (unknown) [55] | Biomedical function | Not given | 4 weeks | Effect of collagen hydrolysate in skins under sun-exposed areas | Elasticity in skin was enhanced |
3.2. Type II
3.2.1. Aimed Biomedical Function for Type II
3.2.2. Novel Method for Type II
| Source | Category | Method | Days | Purpose | Results |
|---|---|---|---|---|---|
| Not given [50] | Biomedical function | Not given | 24 weeks | OA joint pain symptoms after hydrolysate ingestion for 24 weeks | OA athlete’s pain is significantly decreased |
| Articular bovine chondrocytes, collagen hydrolysate source is not given [60] | Bio-synthesis and culturing | Articular cartilage chondrocytes by enzymatic hydrolysis using pronase and collagenase. Collagen hydrolysate source is not given. | 14, 28, 42 days | Culturing of hydrolysate in chondrocyte and agarose gel to enhance Type II collagen in engineered | Compressive properties, Type II collagen are improved. |
| Whale shark (WS) cartilage [71] | Novel Method and Source | Enzymatic (pepsin) and acid hydrolysis. | Whale shark is not extracted before with acid and pepsin hydrolysis. | High denaturation temperature, hydroxyproline, and protein content of pepsin soluble method as compared to acid hydrolysis. | |
| Chicken sternum cartilage extract [72] | Novel Source | Not given | 14 weeks | Using collagen II to reduce skin problems. | Reduction of wrinkles, inhibiting cell damage due to UV rays. |
| Chicken sternum cartilage [68] | Biomedical function | Not given | 180 days | ||
| Type II from bovine cartilage [70] | Novel method | Acidic hydrolysis | A novel technique and material for orthopedic problems. | Better dispersion, mechanical properties. | |
| Porcine [69] | Biomedical function | Not given | 13 days | Low dosage effects of Type II on rat OA. | Less CTX-II, Better postural balance, Better motor activity. |
3.2.3. Novel Source for Type II
3.2.4. Biosynthesis and Culturing for Type II
3.3. Type III
3.3.1. Aimed Biomedical Function for Type III
3.3.2. Novel Sources for Type III
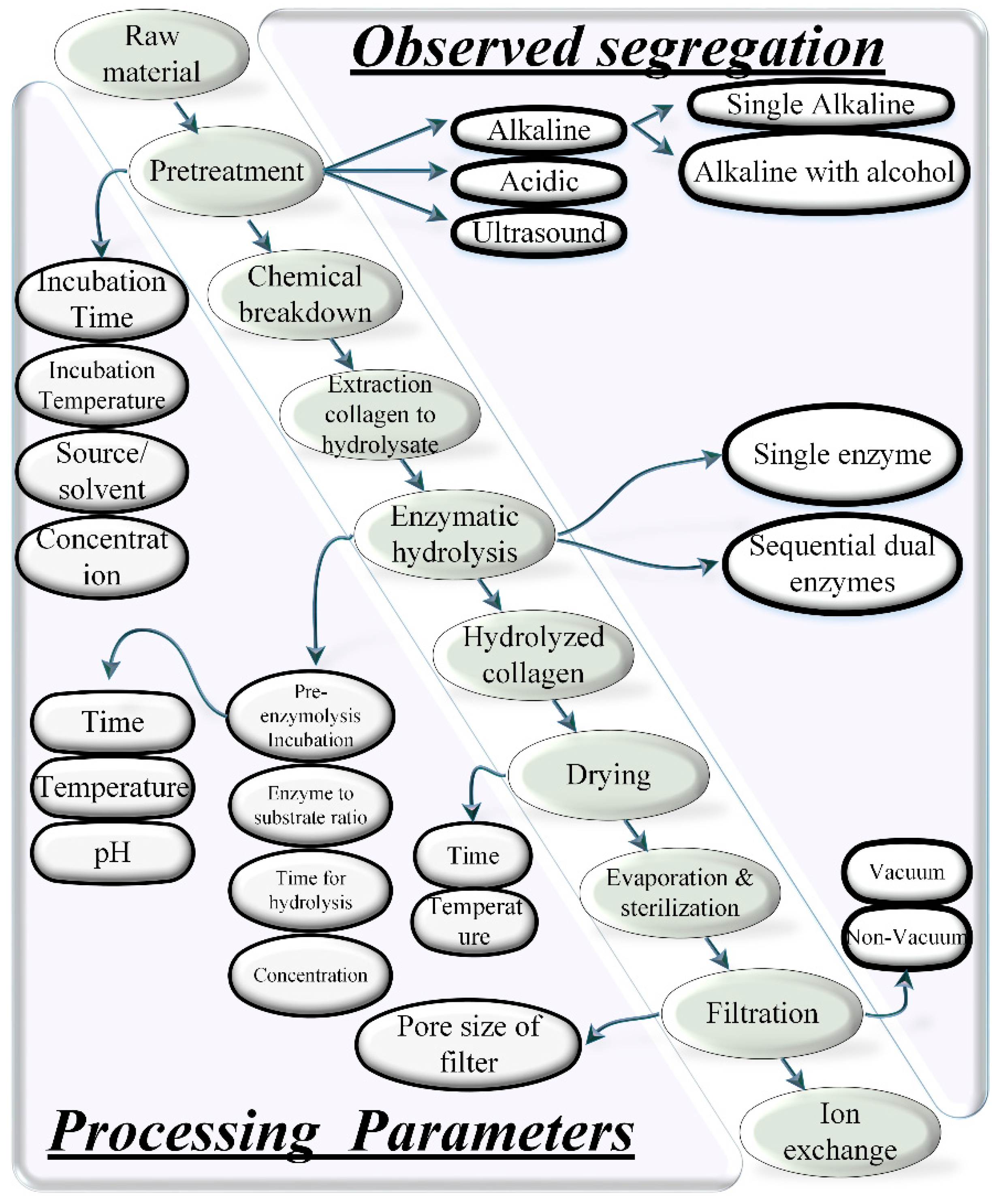
4. Process
4.1. Chemical Hydrolysis
4.2. Enzymatic Hydrolysis
4.3. Pre-Treatments
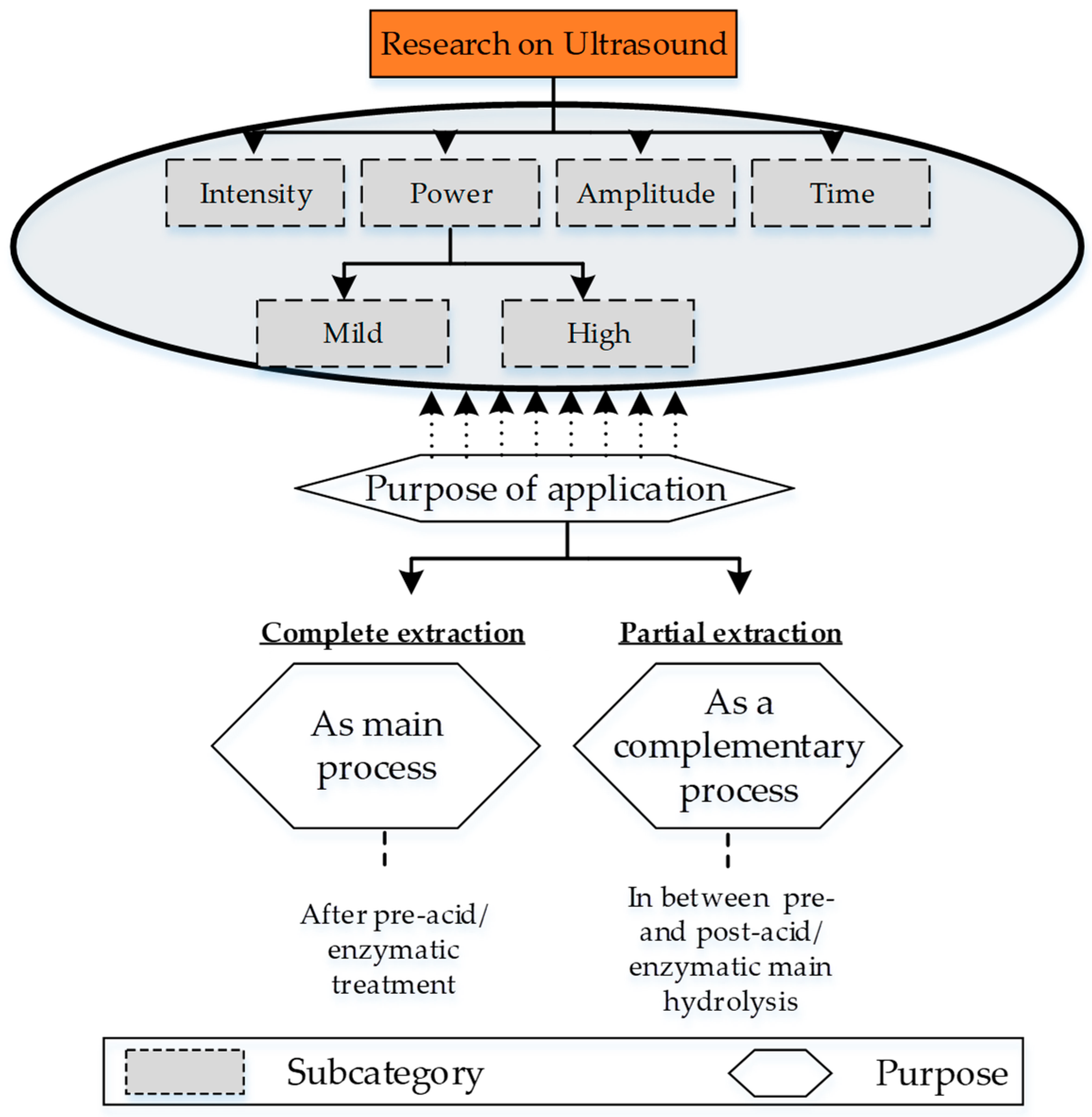
4.4. Ultrasound-Based Extraction
4.5. Super-Critical Carbon Dioxide-Based Extraction
4.6. Hydrostatic Extraction
4.6.1. High Pressure and Temperature with Acid or Alkaline Pre-Treatment
4.6.2. Water Extraction
5. Disease
6. Receptors-Based Collagen
| Collagen Source | Receptor/Proteins/Complex | Activated Pathway | Associated Receptors with Activation of Pathway | Resulting Factors (Anti-Bacterial, Antimicrobial, Anti-Oxidation) | Biomedical Effect |
|---|---|---|---|---|---|
| Prolyl-hydroxyproline (Pro-Hyp) [160] | p75NTR | - | - | - | Wound healing |
| Porcine collagen [19] | PTEN, CDK-2, CDK-4 | PI3K/AKT | Cell proliferation, anti-apoptosis | ||
| Salmo salar skin collagen peptides (Ss-SCPs) and Tilapia nilotica skin collagen peptides [25] | pattern recognition receptor (NOD2) | NF-κB signal pathway | Toll-like receptors (TLRs) | BD14 (modulation through upregulation/down regulation/null) | Wound healing |
| Chum Salmon skin hydrolysate [172] | TGF-β | Smad | TGF-β type II receptors (TβRII) | Type I and III mRNA, TGF-βRII | Skin cellular homeostasis |
| Porcine bone collagen peptide [20] | Runx2 | MAPK | - | - | Osteoblast |
| β-Catenin | Wnt/β-catenin | - | - | ||
| Fish collagen hydrolysate [173] | IL-1β | ERK and p38 | - | MMP13 | Osteoarthritis |
7. Future Perspective and Challenges
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kehinde, B.A.; Sharma, P. Recently isolated antidiabetic hydrolysates and peptides from multiple food sources: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 322–340. [Google Scholar] [CrossRef]
- Kang, N.J.; Jin, H.-S.; Lee, S.-E.; Kim, H.J.; Koh, H.; Lee, D.-W. New approaches towards the discovery and evaluation of bioactive peptides from natural resources. Crit. Rev. Environ. Sci. Technol. 2020, 50, 72–103. [Google Scholar] [CrossRef]
- León-López, A.; Morales-Peñaloza, A.; Martínez-Juárez, V.M.; Vargas-Torres, A.; Zeugolis, D.I.; Aguirre-Álvarez, G. Hydrolyzed Collagen—Sources and Applications. Molecules 2019, 24, 4031. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Nikoo, M.; Boran, G.; Zhou, P.; Regenstein, J.M. Collagen and gelatin. Annu. Rev. Food Sci. Technol. 2015, 6, 527–557. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M. Processing of collagen based biomaterials and the resulting materials properties. Biomed. Eng. Online 2019, 18, 24. [Google Scholar] [CrossRef]
- Sionkowska, A.; Skrzyński, S.; Śmiechowski, K.; Kołodziejczak, A. The review of versatile application of collagen. Polym. Adv. Technol. 2017, 28, 4–9. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, G.; Shi, B. Physicochemical properties of collagen, gelatin and collagen hydrolysate derived from bovine limed split wastes. J. Soc. Leather Technol. Chem. 2006, 90, 23. [Google Scholar]
- Liu, X.; Machado, G.C.; Eyles, J.P.; Ravi, V.; Hunter, D.J. Dietary supplements for treating osteoarthritis: A systematic review and meta-analysis. Br. J. Sports Med. 2018, 52, 167–175. [Google Scholar] [CrossRef] [PubMed]
- El-Sayyad, A.R.I.; Elghareeb, T.I.; Khashaba, M.M.; Zayed, M.A. Evaluation of the Effect of Solcoseryl on Promotion of Bone Regeneration in Calvarial Bony Defect–An Experimental Pilot study. J. Adv. Dent. Res. 2020, 2, 12–23. [Google Scholar] [CrossRef]
- Liu, J.; Wang, J.; Guo, Y. Effect of Collagen Peptide, Alone and in Combination with Calcium Citrate, on Bone Loss in Tail-Suspended Rats. Molecules 2020, 25, 782. [Google Scholar] [CrossRef]
- Ju, H.; Liu, X.; Zhang, G.; Liu, D.; Yang, Y. Comparison of the Structural Characteristics of Native Collagen Fibrils Derived from Bovine Tendons using Two Different Methods: Modified Acid-Solubilized and Pepsin-Aided Extraction. Materials 2020, 13, 358. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zhang, C.; Jia, W.; Shen, Q.; Qin, X.; Zhang, H.; Zhu, L. Metabolomics strategy reveals the osteogenic mechanism of yak (Bos grunniens) bone collagen peptides on ovariectomy-induced osteoporosis in rats. Food Funct. 2020, 11, 1498–1512. [Google Scholar] [CrossRef]
- Ye, M.; Jia, W.; Zhang, C.; Shen, Q.; Zhu, L.; Wang, L. Preparation, identification and molecular docking study of novel osteoblast proliferation-promoting peptides from yak (Bos grunniens) bones. RSC Adv. 2019, 9, 14627–14637. [Google Scholar] [CrossRef]
- Zhang, Y.; Olsen, K.; Grossi, A.; Otte, J. Effect of pretreatment on enzymatic hydrolysis of bovine collagen and formation of ACE-inhibitory peptides. Food Chem. 2013, 141, 2343–2354. [Google Scholar] [CrossRef]
- Fu, Y.; Young, J.F.; Løkke, M.M.; Lametsch, R.; Aluko, R.E.; Therkildsen, M. Revalorisation of bovine collagen as a potential precursor of angiotensin I-converting enzyme (ACE) inhibitory peptides based on in silico and in vitro protein digestions. J. Funct. Foods 2016, 24, 196–206. [Google Scholar] [CrossRef]
- Lima, C.A.; Campos, J.F.; Lima Filho, J.L.; Converti, A.; da Cunha, M.G.C.; Porto, A.L. Antimicrobial and radical scavenging properties of bovine collagen hydrolysates produced by Penicillium aurantiogriseum URM 4622 collagenase. Int. J. Food Sci. Technol. 2015, 52, 4459–4466. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.M.; Lafarga, T.; Hayes, M.; O’Brien, N.M. Bioactivity of bovine lung hydrolysates prepared using papain, pepsin, and Alcalase. J. Food Biochem. 2017, 41, e12406. [Google Scholar] [CrossRef]
- Wang, C.-H.; Hsieh, D.-J.; Periasamy, S.; Chuang, C.-T.; Tseng, F.-W.; Kuo, J.-C.; Tarng, Y.-W. Regenerative porcine dermal collagen matrix developed by supercritical carbon dioxide extraction technology: Role in accelerated wound healing. Acta Mater. 2020, 9, 100576. [Google Scholar] [CrossRef]
- Zhu, L.; Xie, Y.; Wen, B.; Ye, M.; Liu, Y.; Imam, K.M.S.U.; Cai, H.; Zhang, C.; Wang, F.; Xin, F. Porcine bone collagen peptides promote osteoblast proliferation and differentiation by activating the PI3K/Akt signaling pathway. J. Funct. Foods 2020, 64, 103697. [Google Scholar] [CrossRef]
- Wu, W.; He, L.; Li, C.; Zhao, S.; Liang, Y.; Yang, F.; Zhang, M.; Jin, G.; Ma, M. Phosphorylation of porcine bone collagen peptide to improve its calcium chelating capacity and its effect on promoting the proliferation, differentiation and mineralization of osteoblastic MC3T3-E1 cells. J. Funct. Foods 2020, 64, 103701. [Google Scholar] [CrossRef]
- Kim, D.; Kim, Y.H.B.; Ham, J.-S.; Lee, S.K.; Jang, A. Pig Skin Gelatin Hydrolysates Attenuate Acetylcholine Esterase Activity and Scopolamine-induced Impairment of Memory and Learning Ability of Mice. Korean J. Food Sci. 2020, 40, 183. [Google Scholar] [CrossRef]
- Koizumi, S.; Inoue, N.; Sugihara, F.; Igase, M. Effects of Collagen Hydrolysates on Human Brain Structure and Cognitive Function: A Pilot Clinical Study. Nutrients 2020, 12, 50. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Park, K.; Ha, G.; Jung, J.R.; Chang, O.; Ham, J.-S.; Jeong, S.-G.; Park, B.-Y.; Song, J.; Jang, A. Anti-oxidative and neuroprotective activities of pig skin gelatin hydrolysates. Korean J. Food Sci. 2013, 33, 258–267. [Google Scholar] [CrossRef]
- Bang, J.S.; Jin, Y.J.; Choung, S.-Y. Low molecular polypeptide from oyster hydrolysate recovers photoaging in SKH-1 hairless mice. Toxicol. Appl. Pharm. 2020, 386, 114844. [Google Scholar] [CrossRef]
- Mei, F.; Liu, J.; Wu, J.; Duan, Z.; Chen, M.; Meng, K.; Chen, S.; Shen, X.; Xia, G.; Zhao, M. Collagen peptides isolated from salmo salar and tilapia nilotica skin accelerate wound healing by altering cutaneous microbiome colonization via upregulated NOD2 and BD14. J. Agric. Food Chem. 2020, 68, 1621–1633. [Google Scholar] [CrossRef]
- Wang, W.-Y.; Zhao, Y.-Q.; Zhao, G.-X.; Chi, C.-F.; Wang, B. Antioxidant Peptides from Collagen Hydrolysate of Redlip Croaker (Pseudosciaena polyactis) Scales: Preparation, Characterization, and Cytoprotective Effects on H2O2-Damaged HepG2 Cells. Mar. Drugs 2020, 18, 156. [Google Scholar] [CrossRef]
- Wang, S.; Chen, G.; Chen, K.; Kan, J. Bioavailability and prebiotic potential of Carapax Trionycis, a waste from soft-shelled turtle processing. J. Sci. Food Agric. 2020, 100, 2554–2567. [Google Scholar] [CrossRef]
- Sousa, R.O.; Alves, A.L.; Carvalho, D.N.; Martins, E.; Oliveira, C.; Silva, T.H.; Reis, R.L. Acid and enzymatic extraction of collagen from Atlantic cod (Gadus Morhua) swim bladders envisaging health-related applications. J. Biomater. Sci. Polym. Ed. 2020, 31, 20–37. [Google Scholar] [CrossRef]
- Korczek, K.R.; Tkaczewska, J.; Duda, I.; Migdał, W. Effect of Heat Treatment on the Antioxidant and Antihypertensive Activity as Well as in vitro Digestion Stability of Mackerel (Scomber scombrus) Protein Hydrolysates. J. Aquat. Food Prod. Technol. 2020, 29, 73–89. [Google Scholar] [CrossRef]
- Halim, N.; Sarbon, N. Characterization of Asian swamp eel (Monopterus sp.) protein hydrolysate functional properties prepared using Alcalase® enzyme. Food Res. 2020, 4, 207–215. [Google Scholar] [CrossRef]
- Xu, X.H.; Lv, P.F.; Wang, T.X.; Wang, B.X.; Shi, Y.; Wang, B.X.; Meng, Z.R.; Chen, Q.X.; Zhuang, J.X.; Wang, Y.Y. Bone-strengthening effects and safety of compound peptides from skin of Chiloscyllium plagiosum and Mustelus griseus. Food Sci. Nutr. 2020, 13, 1522–1533. [Google Scholar] [CrossRef] [PubMed]
- Akram, A.N.; Zhang, C. Effect of ultrasonication on the yield, functional and physicochemical characteristics of collagen-II from chicken sternal cartilage. Food Chem. 2020, 307, 125544. [Google Scholar] [CrossRef]
- Gál, R.; Mokrejš, P.; Mrázek, P.; Pavlačková, J.; Janáčová, D.; Orsavová, J. Chicken Heads as a Promising By-Product for Preparation of Food Gelatins. Molecules 2020, 25, 494. [Google Scholar] [CrossRef]
- Bichukale, A.; Koli, J.; Sonavane, A.; Vishwasrao, V.; Pujari, K.; Shingare, P. Functional Properties of Gelatin Extracted from Poultry Skin and Bone Waste. IJPAB 2018, 6, 87–101. [Google Scholar]
- Zhao, Y.; Wang, Z.; Zhang, J.; Su, T. Extraction and characterization of collagen hydrolysates from the skin of Rana chensinensis. 3 Biotech. 2018, 8, 181. [Google Scholar] [CrossRef] [PubMed]
- Sonklin, C.; Alashi, M.A.; Laohakunjit, N.; Kerdchoechuen, O.; Aluko, R.E. Identification of antihypertensive peptides from mung bean protein hydrolysate and their effects in spontaneously hypertensive rats. J. Funct. Foods 2020, 64, 103635. [Google Scholar] [CrossRef]
- Vidal, A.R.; Duarte, L.P.; Schmidt, M.M.; Cansian, R.L.; Fernandes, I.A.; de Oliveira Mello, R.; Demiate, I.M.; Dornelles, R.C.P. Extraction and characterization of collagen from sheep slaughter by-products. J. Waste Manag. 2020, 102, 838–846. [Google Scholar] [CrossRef]
- Qiu, W.; Chen, X.; Tian, Y.; Wu, D.; Du, M.; Wang, S. Protection against oxidative stress and anti-aging effect in Drosophila of royal jelly-collagen peptide. Food Chem. Toxicol. 2020, 135, 110881. [Google Scholar] [CrossRef]
- Bechaux, J.; Ferraro, V.; Sayd, T.; Chambon, C.; Le Page, J.F.; Drillet, Y.; Gatellier, P.; Santé-Lhoutellier, V. Workflow towards the generation of bioactive hydrolysates from porcine products by combining in silico and in vitro approaches. Food Res. Int. 2020, 132, 109123. [Google Scholar] [CrossRef]
- Alves, F.E.D.S.B.; Carpiné, D.; Teixeira, G.L.; Goedert, A.C.; de Paula Scheer, A.; Ribani, R.H. Valorization of an Abundant Slaughterhouse By-product as a Source of Highly Technofunctional and Antioxidant Protein Hydrolysates. Waste Biomass Valori. 2020, 1–17. [Google Scholar]
- Nogimura, D.; Mizushige, T.; Taga, Y.; Nagai, A.; Shoji, S.; Azuma, N.; Kusubata, M.; Adachi, S.I.; Yoshizawa, F.; Kabuyama, Y. Prolyl-hydroxyproline, a collagen-derived dipeptide, enhances hippocampal cell proliferation, which leads to antidepressant-like effects in mice. FASEB J. 2020, 34, 5715–5723. [Google Scholar] [CrossRef]
- Tanaka, M.; Koyama, Y.-I.; Nomura, Y. Effects of collagen peptide ingestion on UV-B-induced skin damage. Biosci. Biotech. Biochem. 2009, 73, 930–932. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.G.; Lindhe, J. Ridge preservation with the use of Bio-Oss® collagen: A 6-month study in the dog. Clin. Oral Implant. Res. 2009, 20, 433–440. [Google Scholar] [CrossRef]
- Bornstein, M.M.; Bosshardt, D.; Buser, D. Effect of two different bioabsorbable collagen membranes on guided bone regeneration: A comparative histomorphometric study in the dog mandible. J. Periodontol. 2007, 78, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Minami, S.; Hata, M.; Tamai, Y.; Hashida, M.; Takayama, T.; Yamamoto, S.; Okada, M.; Funatsu, T.; Tsuka, T.; Imagawa, T. Clinical application of D-glucosamine and scale collagen peptide on canine and feline orthopedic diseases and spondylitis deformans. Carbohydr. Polym. 2011, 84, 831–834. [Google Scholar] [CrossRef]
- Tal, H.; Kozlovsky, A.; Artzi, Z.; Nemcovsky, C.E.; Moses, O. Cross-linked and non-cross-linked collagen barrier membranes disintegrate following surgical exposure to the oral environment: A histological study in the cat. Clin. Oral Implant. Res. 2008, 19, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Zentek, J.; Schulz, A. Urinary composition of cats is affected by the source of dietary protein. Nutr. J. 2004, 134, 2162S–2165S. [Google Scholar] [CrossRef]
- Kimura, M.; Nito, T.; Imagawa, H.; Tayama, N.; Chan, R.W. Collagen injection as a supplement to arytenoid adduction for vocal fold paralysis. Ann. Otol. Rhinol. Laryngol. 2008, 117, 430–436. [Google Scholar] [CrossRef]
- Hays, N.P.; Kim, H.; Wells, A.M.; Kajkenova, O.; Evans, W.J. Effects of whey and fortified collagen hydrolysate protein supplements on nitrogen balance and body composition in older women. J. Am. Diet. Assoc. 2009, 109, 1082–1087. [Google Scholar] [CrossRef]
- Clark, K.L.; Sebastianelli, W.; Flechsenhar, K.R.; Aukermann, D.F.; Meza, F.; Millard, R.L.; Deitch, J.R.; Sherbondy, P.S.; Albert, A. 24-Week study on the use of collagen hydrolysate as a dietary supplement in athletes with activity-related joint pain. Curr. Med. Res. Opin. 2008, 24, 1485–1496. [Google Scholar] [CrossRef]
- Bello, A.E.; Oesser, S. Collagen hydrolysate for the treatment of osteoarthritis and other joint disorders: A review of the literature. Curr. Med. Res. Opin. 2006, 22, 2221–2232. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Cruz, G.; León-López, A.; Cruz-Gómez, V.; Jiménez-Alvarado, R.; Aguirre-Álvarez, G. Collagen Hydrolysates for Skin Protection: Oral Administration and Topical Formulation. Antioxidants 2020, 9, 181. [Google Scholar] [CrossRef]
- Crowley, D.C.; Lau, F.C.; Sharma, P.; Evans, M.; Guthrie, N.; Bagchi, M.; Bagchi, D.; Dey, D.K.; Raychaudhuri, S.P. Safety and efficacy of undenatured type II collagen in the treatment of osteoarthritis of the knee: A clinical trial. Int. J. Med. Sci. 2009, 6, 312. [Google Scholar] [CrossRef] [PubMed]
- Watanabe-Kamiyama, M.; Shimizu, M.; Kamiyama, S.; Taguchi, Y.; Sone, H.; Morimatsu, F.; Shirakawa, H.; Furukawa, Y.; Komai, M. Absorption and effectiveness of orally administered low molecular weight collagen hydrolysate in rats. J. Agric. Food Chem. 2010, 58, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Sangsuwan, W.; Asawanonda, P. Four-weeks daily intake of oral collagen hydrolysate results in improved skin elasticity, especially in sun-exposed areas: A randomized, double-blind, placebo-controlled trial. J. Dermatol. Treat. 2020, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (Hypophthalmichthys molitrix), a biorefinery approach. Food Chem. 2018, 242, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T. Anti-Oxidative and Anti-Allergic Activities of Type I Collagen Hydrolysates from Emu Skins as By-Product. SF Food Dairy Tech. J. 2018, 2. Available online: https://www.semanticscholar.org/paper/Anti-Oxidative-and-Anti-Allergic-Activities-of-Type-Nagai-Tanoue/40fbd764c99d0f6c7eddc1f73e1ea79461f612cb (accessed on 10 May 2021).
- Taga, Y.; Kusubata, M.; Ogawa-Goto, K.; Hattori, S.; Funato, N. Collagen-derived X-Hyp-Gly-type tripeptides promote differentiation of MC3T3-E1 pre-osteoblasts. J. Funct. Foods 2018, 46, 456–462. [Google Scholar] [CrossRef]
- Zhang, H.; Pan, D.; Dong, Y.; Su, W.; Su, H.; Wei, X.; Yang, C.; Jing, L.; Tang, X.; Li, X. Transdermal permeation effect of collagen hydrolysates of deer sinew on mouse skin, ex vitro, and antioxidant activity, increased type I collagen secretion of percutaneous proteins in NIH/3T3 cells. J. Cosmet. Dermatol. 2020, 19, 519–528. [Google Scholar] [CrossRef]
- Ng, K.W.; Saliman, J.D.; Lin, E.Y.; Statman, L.Y.; Kugler, L.E.; Lo, S.B.; Ateshian, G.A.; Hung, C.T. Culture duration modulates collagen hydrolysate-induced tissue remodeling in chondrocyte-seeded agarose hydrogels. Ann. Biomed. Eng. 2007, 35, 1914–1923. [Google Scholar] [CrossRef]
- Meng, D.; Li, W.; Ura, K.; Takagi, Y. Effects of phosphate ion concentration on in-vitro fibrillogenesis of sturgeon type I collagen. Int. J. Biol. Macromol. 2020, 148, 182–191. [Google Scholar] [CrossRef]
- Liu, X.; Long, X.; Gao, Y.; Liu, W.; Hayashi, T.; Mizuno, K.; Hattori, S.; Fujisaki, H.; Ogura, T.; Onodera, S. Type I collagen inhibits adipogenic differentiation via YAP activation in vitro. J. Cell. Physiol. 2020, 235, 1821–1837. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, Y.; Long, X.; Hayashi, T.; Mizuno, K.; Hattori, S.; Fujisaki, H.; Ogura, T.; Wang, D.O.; Ikejima, T. Type I collagen promotes the migration and myogenic differentiation of C2C12 myoblasts via the release of interleukin-6 mediated by FAK/NF-κB p65 activation. Food Funct. 2020, 11, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Oesser, S.; Adam, M.; Babel, W.; Seifert, J. Oral administration of 14C labeled gelatin hydrolysate leads to an accumulation of radioactivity in cartilage of mice (C57/BL). Nutr. J. 1999, 129, 1891–1895. [Google Scholar] [CrossRef] [PubMed]
- Oesser, S.; Seifert, J. Stimulation of type II collagen biosynthesis and secretion in bovine chondrocytes cultured with degraded collagen. Cell Tissue Res. 2003, 311, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Oesser, S.; Seifert, J. Impact of Collagen Fragments on the Synthesis and Degradation of the Extracellular Matrix of Cartilage Tissue. 2005. Available online: http://flexovitalis.com/wp-content/uploads/2015/05/S-Oesser-2005_eng.pdf (accessed on 21 May 2021).
- Ng, K.; Saliman, J.; Tan, K.; Statman, L. Collagen Hydrolysate Increases the Mechanical Properties and Type II Collagen Synthesis of Tissue Engineered Articular Cartilage. In Proceedings of the 51st Annual Meeting of the ORS, Washington, DC, USA, 20–23 February 2005; Orthopaedic Research Society: Washington, DC, USA, 2015. Available online: https://www.ors.org/Transactions/51/0259.pdf (accessed on 21 May 2021).
- Lugo, J.P.; Saiyed, Z.M.; Lane, N.E. Efficacy and tolerability of an undenatured type II collagen supplement in modulating knee osteoarthritis symptoms: A multicenter randomized, double-blind, placebo-controlled study. Nutr. J. 2015, 15, 14. [Google Scholar] [CrossRef]
- Mannelli, L.D.C.; Micheli, L.; Zanardelli, M.; Ghelardini, C. Low dose native type II collagen prevents pain in a rat osteoarthritis model. BMC Musculoskelet. Disord. 2013, 14, 228. [Google Scholar]
- Senra, M.R.; Marques, M.d.F.V. Thermal and mechanical behavior of ultra-high molecular weight polyethylene/collagen blends. J. Mech. Behav. Biomed. 2020, 103, 103577. [Google Scholar] [CrossRef]
- Jeevithan, E.; Jingyi, Z.; Wang, N.; He, L.; Bao, B.; Wu, W. Physico-chemical, antioxidant and intestinal absorption properties of whale shark type-II collagen based on its solubility with acid and pepsin. Process. Biochem. 2015, 50, 463–472. [Google Scholar] [CrossRef]
- Phipps, K.R.; Lee, H.Y.; Kim, H.; Jeon, B. Oral administration of a novel hydrolyzed chicken sternal cartilage extract (BioCell Collagen®) reduces UVB-induced photoaging in mice. J. Funct. Foods 2020, 68, 103870. [Google Scholar] [CrossRef]
- Xu, D.; Li, D.; Zhao, Z.; Wu, J.; Zhao, M. Regulation by walnut protein hydrolysate on the components and structural degradation of photoaged skin in SD rats. Food Funct 2019, 10, 6792–6802. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Q.; Wang, L.; Xu, W.; He, Y.; Li, Y.; He, S.; Ma, H. Improvement of skin condition by oral administration of collagen hydrolysates in chronologically aged mice. J. Sci. Food Agric. 2017, 97, 2721–2726. [Google Scholar] [CrossRef]
- Song, H.; Zhang, S.; Zhang, L.; Li, B. Effect of orally administered collagen peptides from bovine bone on skin aging in chronologically aged mice. Nutrients 2017, 9, 1209. [Google Scholar] [CrossRef] [PubMed]
- Kashiuchi, S.; Miyazawa, R.; Nagata, H.; Shirai, M.; Shimizu, M.; Sone, H.; Kamiyama, S. Effects of administration of glucosamine and chicken cartilage hydrolysate on rheumatoid arthritis in SKG mice. Food Funct. 2019, 10, 5008–5017. [Google Scholar] [CrossRef] [PubMed]
- Bolke, L.; Schlippe, G.; Gerß, J.; Voss, W. A collagen supplement improves skin hydration, elasticity, roughness, and density: Results of a randomized, placebo-controlled, blind study. Nutrients 2019, 11, 2494. [Google Scholar] [CrossRef]
- Fan, J.; Zhuang, Y.; Li, B. Effects of collagen and collagen hydrolysate from jellyfish umbrella on histological and immunity changes of mice photoaging. Nutrients 2013, 5, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, X.; Bai, F.; Fang, Y.; Wang, J.; Gao, R. The anti-skin-aging effect of oral administration of gelatin from the swim bladder of Amur sturgeon (Acipenser schrenckii). Food Funct. 2019, 10, 3890–3897. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Dornelles, R.; Mello, R.; Kubota, E.; Mazutti, M.; Kempka, A.; Demiate, I. Collagen extraction process. Int. Food Res. J. 2016, 23, 913–922. [Google Scholar]
- Sun, L.; Zhang, S.; Yuan, L.; Yang, Y.; Lin, S. Glutamine and methionine targeted pulsed electric field treatment for enhanced immune activity in pine nut Gln-Trp-Phe-Met peptides. Int. J. Food Sci. Technol. 2020, 55, 2954–2961. [Google Scholar] [CrossRef]
- Mohammad, A.W.; Suhimi, N.M.; Aziz, A.G.K.A.; Jahim, J.M. Resources: Potential for Further Development. Res. J. Appl. Sci. 2014, 14, 1319–1323. [Google Scholar]
- Nalinanon, S.; Benjakul, S.; Kishimura, H. Collagens from the skin of arabesque greenling (Pleurogrammus azonus) solubilized with the aid of acetic acid and pepsin from albacore tuna (Thunnus alalunga) stomach. J. Sci. Food Agric. 2010, 90, 1492–1500. [Google Scholar] [CrossRef] [PubMed]
- Sadowska, M.; Kołodziejska, I.; Niecikowska, C. Isolation of collagen from the skins of Baltic cod (Gadus morhua). Food Chem. 2003, 81, 257–262. [Google Scholar] [CrossRef]
- Nagai, T.; Suzuki, N. Preparation and partial characterization of collagen from paper nautilus (Argonauta argo, Linnaeus) outer skin. Food Chem. 2002, 76, 149–153. [Google Scholar] [CrossRef]
- Nagai, T.; Araki, Y.; Suzuki, N. Collagen of the skin of ocellate puffer fish (Takifugu rubripes). Food Chem. 2002, 78, 173–177. [Google Scholar] [CrossRef]
- Nagai, T.; Suzuki, N. Isolation of collagen from fish waste material—skin, bone and fins. Food Chem. 2000, 68, 277–281. [Google Scholar] [CrossRef]
- Sivakumar, P.; Chandrakasan, G. Occurrence of a novel collagen with three distinct chains in the cranial cartilage of the squid Sepia officinalis: Comparison with shark cartilage collagen. Biochim. Biophys. Acta 1998, 1381, 161–169. [Google Scholar] [CrossRef]
- Ciarlo, A.S.; Paredi, M.E.; Fraga, A.N. Isolation of soluble collagen from hake skin (Merluccius hubbsi). J. Aquat. Food Prod. Technol. 1997, 6, 65–77. [Google Scholar] [CrossRef]
- Nomura, Y.; Sakai, H.; Ishii, Y.; Shirai, K. Preparation and some properties of type I collagen from fish scales. Biosci. Biotech. Biochem. 1996, 60, 2092–2094. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Miyauchi, Y.; Uchida, N. Scale and bone type I collagens of carp (Cyprinus carpio). Comp. Biochem. Phys. B 1991, 99, 473–476. [Google Scholar] [CrossRef]
- Alemán, A.; Giménez, B.; Gómez-Guillén, M.C.; Montero, P. Enzymatic hydrolysis of fish gelatin under high pressure treatment. Int. J. Food Sci. Technol. 2011, 46, 1129–1136. [Google Scholar] [CrossRef]
- Huang, Y.-Q.; Guan, R.; Huang, M.-Z. Study on hydrolysis of macromolecular gelatin with enzymes in combination mode. Chin. J. Polym. Sci. 2004, 22, 599–602. [Google Scholar]
- Jia, J.; Zhou, Y.; Lu, J.; Chen, A.; Li, Y.; Zheng, G. Enzymatic hydrolysis of Alaska pollack (Theragra chalcogramma) skin and antioxidant activity of the resulting hydrolysate. J. Sci. Food Agric. 2010, 90, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Li, B.F. Radical scavenging properties of protein hydrolysates from Jumbo flying squid (Dosidicus eschrichitii Steenstrup) skin gelatin. J. Sci. Food Agric. 2006, 86, 2290–2295. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Byun, H.-G.; Kim, S.-K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-I.; Ho, H.-Y.; Chu, Y.-J.; Chow, C.-J. Characteristic and antioxidant activity of retorted gelatin hydrolysates from cobia (Rachycentron canadum) skin. Food Chem. 2008, 110, 128–136. [Google Scholar] [CrossRef]
- Wang, L.; Yang, B.; Du, X.; Yang, Y.; Liu, J. Optimization of conditions for extraction of acid-soluble collagen from grass carp (Ctenopharyngodon idella) by response surface methodology. Innov. Food Sci. Emerg. 2008, 9, 604–607. [Google Scholar] [CrossRef]
- Liu, D.; Wei, G.; Li, T.; Hu, J.; Lu, N.; Regenstein, J.M.; Zhou, P. Effects of alkaline pretreatments and acid extraction conditions on the acid-soluble collagen from grass carp (Ctenopharyngodon idella) skin. Food Chem. 2015, 172, 836–843. [Google Scholar] [CrossRef]
- Regenstein, J.; Zhou, P. Collagen and gelatin from marine by-products. In Maximising the Value of Marine By-Products; Elsevier: Amsterdam, The Netherlands, 2007; pp. 279–303. [Google Scholar]
- Muyonga, J.; Cole, C.; Duodu, K. Characterisation of acid soluble collagen from skins of young and adult Nile perch (Lates niloticus). Food Chem. 2004, 85, 81–89. [Google Scholar] [CrossRef]
- Nagai, T.; Suzuki, N.; Nagashima, T. Collagen from common minke whale (Balaenoptera acutorostrata) unesu. Food Chem. 2008, 111, 296–301. [Google Scholar] [CrossRef]
- Sato, K.; Yoshinaka, R.; Sato, M.; Shimizu, Y. Isolation of native acid-soluble collagen from fish muscle. JSFS 1987, 53, 1431–1436. [Google Scholar] [CrossRef][Green Version]
- Yang, H.; Wang, Y.; Jiang, M.; Oh, J.H.; Herring, J.; Zhou, P. 2-Step optimization of the extraction and subsequent physical properties of channel catfish (Ictalurus punctatus) skin gelatin. J. Food Sci. 2007, 72, C188–C195. [Google Scholar] [CrossRef]
- Zhou, P.; Regenstein, J.M. Effects of alkaline and acid pretreatments on Alaska pollock skin gelatin extraction. J. Food Sci. 2005, 70, c392–c396. [Google Scholar] [CrossRef]
- Hou, P.; Regenstein, J. Optimization of extraction conditions for pollock skin gelatin. J. Food Sci. 2004, 69, C393–C398. [Google Scholar] [CrossRef]
- Li, D.; Mu, C.; Cai, S.; Lin, W. Ultrasonic irradiation in the enzymatic extraction of collagen. Ultrason. Sonochem. 2009, 16, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Ovsianko, S.; Chernyavsky, E.; Minchenya, V.; Adzerikho, I.; Shkumatov, V. Effect of ultrasound on activation of serine proteases precursors. Ultrason. Sonochem. 2005, 12, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.M.; Wan, M.X.; Wang, S.P.; Kang, J.Q. Effects of ultrasound and additives on the function and structure of trypsin. Ultrason. Sonochem. 2004, 11, 399–404. [Google Scholar] [CrossRef]
- Amiri, A.; Sharifian, P.; Soltanizadeh, N. Application of ultrasound treatment for improving the physicochemical, functional and rheological properties of myofibrillar proteins. Int. J. Biol. Macromol. 2018, 111, 139–147. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, Y.H.; Kim, Y.J.; Park, H.J.; Lee, N.H. Effects of ultrasonic treatment on collagen extraction from skins of the sea bass Lateolabrax japonicus. Fish. Res. 2012, 78, 485–490. [Google Scholar] [CrossRef]
- Ran, X.G.; Wang, L.Y. Use of ultrasonic and pepsin treatment in tandem for collagen extraction from meat industry by-products. J. Sci. Food Agric. 2014, 94, 585–590. [Google Scholar] [CrossRef]
- Barros, A.A.; Aroso, I.M.; Silva, T.H.; Mano, J.F.; Duarte, A.R.C.; Reis, R.L. Water and carbon dioxide: Green solvents for the extraction of collagen/gelatin from marine sponges. ACS Sustain. Chem. Eng. 2015, 3, 254–260. [Google Scholar] [CrossRef]
- Silva, J.C.; Barros, A.A.; Aroso, I.M.; Fassini, D.; Silva, T.H.; Reis, R.L.; Duarte, A.R.C. Extraction of collagen/gelatin from the marine demosponge Chondrosia reniformis (Nardo, 1847) using water acidified with carbon dioxide–process optimization. Ind. Eng. Chem. Res. 2016, 55, 6922–6930. [Google Scholar] [CrossRef]
- Walkenström, P.; Hermansson, A.-M. High-pressure treated mixed gels of gelatin and whey proteins. Food Hydrocoll. 1997, 11, 195–208. [Google Scholar] [CrossRef]
- Montero, P.; Fernández-Díaz, M.; Gómez-Guillén, M. Characterization of gelatin gels induced by high pressure. Food Hydrocoll. 2002, 16, 197–205. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.; Giménez, B.; Montero, P. Extraction of gelatin from fish skins by high pressure treatment. Food Hydrocoll. 2005, 19, 923–928. [Google Scholar] [CrossRef]
- Uriarte-Montoya, M.H.; Santacruz-Ortega, H.; Cinco-Moroyoqui, F.J.; Rouzaud-Sández, O.; Plascencia-Jatomea, M.; Ezquerra-Brauer, J.M. Giant squid skin gelatin: Chemical composition and biophysical characterization. Food Res. Int. 2011, 44, 3243–3249. [Google Scholar] [CrossRef]
- Kołodziejska, I.; Skierka, E.; Sadowska, M.; Kołodziejski, W.; Niecikowska, C. Effect of extracting time and temperature on yield of gelatin from different fish offal. Food Chem. 2008, 107, 700–706. [Google Scholar] [CrossRef]
- Liu, H.Y.; Li, D.; Guo, S.D. Extraction and properties of gelatin from channel catfish (Ietalurus punetaus) skin. LWT Food Sci. Technol. 2008, 41, 414–419. [Google Scholar] [CrossRef]
- Lee, S.K.; Posthauer, M.E.; Dorner, B.; Redovian, V.; Maloney, M.J. Pressure ulcer healing with a concentrated, fortified, collagen protein hydrolysate supplement: A randomized controlled trial. Adv. Ski. Wound Care 2006, 19, 92–96. [Google Scholar] [CrossRef]
- Pinotti, E.; Montuori, M.; Borrelli, V.; Giuffrè, M.; Angrisani, L. Sarcopenia: What a Surgeon Should Know. Obes. Surg. 2020, 1–6. [Google Scholar] [CrossRef]
- Kemmler, W.; Weineck, M.; Kohl, M.; Stengel, S.V.; Giessing, J.; Fröhlich, M.; Schoene, D. High Intensity Resistance Exercise Training to Improve Body Composition and Strength in Older Men with Osteosarcopenia. Results of the Randomized Controlled Franconian Osteopenia and Sarcopenia Trial (FrOST). Front. Sports Act. Living 2020, 2, 4. [Google Scholar] [CrossRef]
- Zhu, C.-F.; Li, G.-Z.; Peng, H.-B.; Zhang, F.; Chen, Y.; Li, Y. Treatment with marine collagen peptides modulates glucose and lipid metabolism in Chinese patients with type 2 diabetes mellitus. Appl. Physiol. Nutr. Metab. 2010, 35, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-Y.; Hsieh, C.-H.; Hung, C.-C.; Jao, C.-L.; Chen, M.-C.; Hsu, K.-C. Fish skin gelatin hydrolysates as dipeptidyl peptidase IV inhibitors and glucagon-like peptide-1 stimulators improve glycaemic control in diabetic rats: A comparison between warm-and cold-water fish. J. Funct. Foods 2015, 19, 330–340. [Google Scholar] [CrossRef]
- Nasri, R.; Abdelhedi, O.; Jemil, I.; Daoued, I.; Hamden, K.; Kallel, C.; Elfeki, A.; Lamri-Senhadji, M.; Boualga, A.; Nasri, M. Ameliorating effects of goby fish protein hydrolysates on high-fat-high-fructose diet-induced hyperglycemia, oxidative stress and deterioration of kidney function in rats. Chem. Biol. Interact. 2015, 242, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Li-Chan, E.C.; Hunag, S.-L.; Jao, C.-L.; Ho, K.-P.; Hsu, K.-C. Peptides derived from Atlantic salmon skin gelatin as dipeptidyl-peptidase IV inhibitors. J. Agric. Food Chem. 2012, 60, 973–978. [Google Scholar] [CrossRef]
- Huang, S.-L.; Jao, C.-L.; Ho, K.-P.; Hsu, K.-C. Dipeptidyl-peptidase IV inhibitory activity of peptides derived from tuna cooking juice hydrolysates. Peptides 2012, 35, 114–121. [Google Scholar] [CrossRef]
- Hsu, K.-C.; Tung, Y.-S.; Huang, S.-L.; Jao, C.-L. Dipeptidyl Peptidase-IV Inhibitory Activity of Peptides in Porcine Skin Gelatin Hydrolysates, Bioactive Food Peptides in Health and Disease; Intech Open: London, UK, 2013; pp. 205–218. [Google Scholar]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of novel DPP–IV inhibitory peptides from Atlantic salmon (Salmo salar) skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.-X.; Wang, B.-K.; Wu, Y.-C.; Li, Q.-Y.; Qin, B.-W.; Li, H.-J. Release of antidiabetic peptides from Stichopus japonicas by simulated gastrointestinal digestion. Food Chem 2020, 315, 126273. [Google Scholar] [CrossRef]
- Iandolo, D.; Sheard, J.; Levy, G.K.; Pitsalidis, C.; Tan, E.; Dennis, A.; Kim, J.-S.; Markaki, A.E.; Widera, D.; Owens, R.M. Biomimetic and electroactive 3D scaffolds for human neural crest-derived stem cell expansion and osteogenic differentiation. MRS Commun. 2020, 10, 179–187. [Google Scholar] [CrossRef]
- Mathiesen, I.H.; Hitz, M.F.; Katzenstein, T.L.; Oturai, P.; Skov, M.; Jørgensen, N.R.; Jensen, P.O.; Mikkelsen, C.R.; Krogh-Madsen, R.; Pressler, T. Markers of bone turnover are reduced in patients with CF related diabetes; the role of glucose. J. Cyst Fibros. 2019, 18, 436–441. [Google Scholar] [CrossRef]
- Zhang, R.; Gong, H.; Zhu, D.; Gao, J.; Fang, J.; Fan, Y. Seven day insertion rest in whole body vibration improves multi-level bone quality in tail suspension rats. PLoS ONE 2014, 9, e92312. [Google Scholar] [CrossRef]
- Xing, L.-Z.; Ni, H.-J.; Wang, Y.-L. Quercitrin attenuates osteoporosis in ovariectomized rats by regulating mitogen-activated protein kinase (MAPK) signaling pathways. Biomed. Pharm. 2017, 89, 1136–1141. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Chen, A.; Han, Z.; Wang, Y.; Lu, X.; Yang, Y. 1H NMR-based metabonomic study on the effects of Epimedium on glucocorticoid-induced osteoporosis. J. Chromatogr. B Biomed. Appl. 2016, 1038, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Hooshmand, S.; Elam, M.; Browne, J.; Campbell, S.; Payton, M. Evidence for bone reversal properties of a calcium-collagen chelate, a novel dietary supplement. J. Food Nutr. Disor. 2013, 1, 2. [Google Scholar]
- Yang, T.-H.; Hsu, P.-Y.; Meng, M.; Su, C.-C. Supplement of 5-hydroxytryptophan before induction suppresses inflammation and collagen-induced arthritis. Arthritis Res. 2015, 17, 364. [Google Scholar] [CrossRef]
- Ichimura, T.; Yamanaka, A.; Otsuka, T.; Yamashita, E.; Maruyama, S. Antihypertensive effect of enzymatic hydrolysate of collagen and Gly-Pro in spontaneously hypertensive rats. Biosci. Biotech. Biochem. 2009, 73, 2317–2319. [Google Scholar] [CrossRef] [PubMed]
- Saiga-Egusa, A.; Iwai, K.; Hayakawa, T.; Takahata, Y.; Morimatsu, F. Antihypertensive effects and endothelial progenitor cell activation by intake of chicken collagen hydrolysate in pre-and mild-hypertension. Biosci. Biotech. Biochem. 2009, 73, 422–424. [Google Scholar] [CrossRef]
- Lin, H.-C.; Alashi, A.M.; Aluko, R.E.; Sun Pan, B.; Chang, Y.-W. Antihypertensive properties of tilapia (Oreochromis spp.) frame and skin enzymatic protein hydrolysates. Food Nutr. Res. 2017, 61, 1391666. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Chen, G.-W.; Yeh, C.H.; Song, H.; Tsai, J.-S. Purification and identification of angiotensin I-converting enzyme inhibitory peptides and the antihypertensive effect of Chlorella sorokiniana protein hydrolysates. Nutrients 2018, 10, 1397. [Google Scholar] [CrossRef]
- Cao, S.; Wang, Y.; Hao, Y.; Zhang, W.; Zhou, G. Antihypertensive effects in vitro and vivo of novel angiotensin-converting enzyme inhibitory peptides from bovine bone gelatin hydrolysate. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef]
- Fu, W.; Wang, P.; Wu, H.; Zhang, Z.; Zeng, H.; Zhang, Y.; Zheng, B.; Hu, J. Antihypertensive effects of Trichiurus lepturus myosin hydrolysate in spontaneously hypertensive rats. Food Funct. 2020, 11, 3645–3656. [Google Scholar] [CrossRef]
- Zhao, Y.-Q.; Zhang, L.; Tao, J.; Chi, C.-F.; Wang, B. Eight antihypertensive peptides from the protein hydrolysate of Antarctic krill (Euphausia superba): Isolation, identification, and activity evaluation on human umbilical vein endothelial cells (HUVECs). Food Res. Int. 2019, 121, 197–204. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Chen, C.-A.; Tsai, J.-S.; Chen, G.-W. Preparation and Identification of Novel Antihypertensive Peptides from the In Vitro Gastrointestinal Digestion of Marine Cobia Skin Hydrolysates. Nutrients 2019, 11, 1351. [Google Scholar] [CrossRef] [PubMed]
- Nasri, M. Bioactive Peptides from Fish Collagen Byproducts: A Review; Wiley: Hoboken, NJ, USA, 2019; pp. 309–333. [Google Scholar]
- Alvarado, Y.; Muro, C.; Illescas, J.; del Carmen Díaz, M.; Riera, F. Encapsulation of Antihypertensive Peptides from Whey Proteins and Their Releasing in Gastrointestinal Conditions. Biomolecules 2019, 9, 164. [Google Scholar] [CrossRef] [PubMed]
- Mizushige, T.; Nogimura, D.; Nagai, A.; Mitsuhashi, H.; Taga, Y.; Kusubata, M.; Hattori, S.; Kabuyama, Y. Ginger-Degraded Collagen Hydrolysate Exhibits Antidepressant Activity in Mice. J. Nutr. Sci. Vitaminol. 2019, 65, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Su, G.; Zhao, T.; Sun, B.; Zheng, L.; Zhao, M. Neuroprotection of round scad (Decapterus maruadsi) hydrolysate in glutamate-damaged PC12 cells: Possible involved signaling pathways and potential bioactive peptides. J. Funct. Foods 2020, 64, 103690. [Google Scholar] [CrossRef]
- Chan, C.-J.; Tseng, J.-K.; Wang, S.-Y.; Lin, Y.-L.; Wu, Y.-H.S.; Chen, J.-W.; Chen, Y.-C. Ameliorative effects of functional chalaza hydrolysates prepared from protease-A digestion on cognitive dysfunction and brain oxidative damages. Poult. Sci. J. 2020, 99, 2819–2832. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Yu, H.; Huang, C.; Zhong, Q.; Chen, Y.; Xie, J.; Zhou, Z.; Xu, J.; Wang, H. Inhibition of phosphodiesterase 4 by FCPR16 protects SH-SY5Y cells against MPP+-induced decline of mitochondrial membrane potential and oxidative stress. Redox Biol. 2018, 16, 47–58. [Google Scholar] [CrossRef]
- Lee, J.; Song, K.; Huh, E.; Oh, M.S.; Kim, Y.S. Neuroprotection against 6-OHDA toxicity in PC12 cells and mice through the Nrf2 pathway by a sesquiterpenoid from Tussilago farfara. Redox Biol. 2018, 18, 6–15. [Google Scholar] [CrossRef]
- Pincelli, C. p75 neurotrophin receptor in the skin: Beyond its neurotrophic function. Front. Med. 2017, 4, 22. [Google Scholar] [CrossRef]
- Wang, E.X.; Jiang, X. Stem cells from trabecular meshwork cells can secrete extracellular matrix. Biochem. Biophys. Res. Commun. 2020, 523, 522–526. [Google Scholar] [CrossRef]
- Fabricant, R.N.; De Larco, J.E.; Todaro, G.J. Nerve growth factor receptors on human melanoma cells in culture. Proc. Natl. Acad. Sci. USA 1977, 74, 565–569. [Google Scholar] [CrossRef]
- Meeker, R.; Williams, K. Dynamic nature of the p75 neurotrophin receptor in response to injury and disease. J. Neuroimmune Pharm. 2014, 9, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Hasebe, Y.; Hasegawa, S.; Hashimoto, N.; Toyoda, M.; Matsumoto, K.; Umezawa, A.; Yagami, A.; Matsunaga, K.; Mizutani, H.; Nakata, S. Analysis of cell characterization using cell surface markers in the dermis. J. Dermatol. Sci. 2011, 62, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Akamatsu, H.; Hasegawa, S.; Yamamoto, N.; Yoshimura, T.; Hasebe, Y.; Inoue, Y.; Mizutani, H.; Uzawa, T.; Matsunaga, K. Age-related changes of p75 neurotrophin receptor-positive adipose-derived stem cells. J. Dermatol. Sci. 2010, 58, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.T.; Yoshikawa, K.; Sawada, K.; Fukamizu, K.; Koyama, Y.-I.; Shigemura, Y.; Jimi, S.; Sato, K. Mouse skin fibroblasts with mesenchymal stem cell marker p75 neurotrophin receptor proliferate in response to prolyl-hydroxyproline. J. Funct. Foods 2020, 66, 103792. [Google Scholar] [CrossRef]
- Fan, Y.-S.; Li, Q.; Hamdan, N.; Bian, Y.-F.; Zhuang, S.; Fan, K.; Liu, Z.-J. Tetrahydroxystilbene Glucoside Regulates Proliferation, Differentiation, and OPG/RANKL/M-CSF Expression in MC3T3-E1 Cells via the PI3K/Akt Pathway. Molecules 2018, 23, 2306. [Google Scholar] [CrossRef]
- Ding, G.; Zhao, J.; Jiang, D. Allicin inhibits oxidative stress-induced mitochondrial dysfunction and apoptosis by promoting PI3K/AKT and CREB/ERK signaling in osteoblast cells. Exp. Med. 2016, 11, 2553–2560. [Google Scholar] [CrossRef]
- Fujita, T.; Azuma, Y.; Fukuyama, R.; Hattori, Y.; Yoshida, C.; Koida, M.; Ogita, K.; Komori, T. Runx2 induces osteoblast and chondrocyte differentiation and enhances their migration by coupling with PI3K-Akt signaling. J. Cell Biol. 2004, 166, 85–95. [Google Scholar] [CrossRef]
- Levy, A.; Stedman, A.; Deutsch, E.; Donnadieu, F.; Virgin, H.W.; Sansonetti, P.J.; Nigro, G. Innate immune receptor NOD2 mediates LGR5+ intestinal stem cell protection against ROS cytotoxicity via mitophagy stimulation. Proc. Natl. Acad. Sci. USA 2020, 117, 1994–2003. [Google Scholar] [CrossRef]
- Wang, G.; Fu, Y.; Ma, K.; Liu, J.; Liu, X. NOD2 regulates microglial inflammation through the TAK1-NF-κB pathway and autophagy activation in murine pneumococcal meningitis. Brain Res. Bull. 2020, 158, 20–30. [Google Scholar] [CrossRef]
- Williams, H.; Campbell, L.; Crompton, R.A.; Singh, G.; McHugh, B.J.; Davidson, D.J.; McBain, A.J.; Cruickshank, S.M.; Hardman, M.J. Microbial host interactions and impaired wound healing in mice and humans: Defining a role for BD14 and NOD2. J. Investig. Dermatol. 2018, 138, 2264–2274. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, T.; Hovingh, E.S.; Foerster, E.G.; Abdel-Nour, M.; Philpott, D.J.; Girardin, S.E. NOD1 and NOD2 in inflammation, immunity and disease. Arch. Biochem. Biophys. 2019, 670, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, X.; Liang, J.; Wu, J.; Tan, D.; Hu, W. Lefty-1 inhibits renal epithelial–mesenchymal transition by antagonizing the TGF-β/Smad signaling pathway. J. Mol. Histol. 2020, 51, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiong, L.; Tang, J.; Zhu, G.; Dai, R.; Li, L. Mouse skin-derived precursors alleviates ultraviolet B irradiation damage via early activation of TGF-β/Smad pathway by thrombospondin1. Cell Cycle 2020, 19, 492–503. [Google Scholar] [CrossRef]
- Olsen, O.E.; Hella, H.; Elsaadi, S.; Jacobi, C.; Martinez-Hackert, E.; Holien, T. Activins as Dual Specificity TGF-β Family Molecules: SMAD-Activation via Activin-and BMP-Type 1 Receptors. Biomolecules 2020, 10, 519. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Transforming growth factor–β in tissue fibrosis. J. Exp. Med. 2020, 21. [Google Scholar] [CrossRef]
- Liang, J.; Pei, X.; Zhang, Z.; Wang, N.; Wang, J.; Li, Y. The protective effects of long-term oral administration of marine collagen hydrolysate from chum salmon on collagen matrix homeostasis in the chronological aged skin of sprague-dawley male rats. J. Food Sci. 2010, 75, H230–H238. [Google Scholar] [CrossRef]
- Boonmaleerat, K.; Wanachewin, O.; Phitak, T.; Pothacharoen, P.; Kongtawelert, P. Fish collagen hydrolysates modulate cartilage metabolism. Cell Biochem. Biophys. 2018, 76, 279–292. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Matsumoto, M.; Kato, R.; Nimura, A.; Yoshii, T.; Okawa, A.; Fujita, K. Activation of cancer-related and mitogen-activated protein kinase signaling pathways in human mature osteoblasts isolated from patients with type 2 diabetes. Bone Rep. 2019, 10, 100199. [Google Scholar] [CrossRef]
- Kim, J.-M.; Yang, Y.-S.; Park, K.H.; Oh, H.; Greenblatt, M.B.; Shim, J.-H. The ERK MAPK pathway is essential for skeletal development and homeostasis. Int. J. Mol. Sci. 2019, 20, 1803. [Google Scholar] [CrossRef]
- Wysokinski, D.; Pawlowska, E.; Blasiak, J. RUNX2: A master bone growth regulator that may be involved in the DNA damage response. DNA Cell Biol. 2015, 34, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Gomathi, K.; Akshaya, N.; Srinaath, N.; Moorthi, A.; Selvamurugan, N. Regulation of Runx2 by post-translational modifications in osteoblast differentiation. Life Sci. 2020, 117389. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Zhao, Q.; Wang, J.; He, X. MicroRNA-10a influences osteoblast differentiation and angiogenesis by regulating β-catenin expression. Cell Physiol. Biochem. 2015, 37, 2194–2208. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wu, F.; Zhang, Y.; Zhang, Y.; Wang, F.; Jiang, M.; Wang, Z.; Zhang, M.; Li, S.; Yang, L. Irisin promotes human umbilical vein endothelial cell proliferation through the ERK signaling pathway and partly suppresses high glucose-induced apoptosis. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Bagi, C.M.; Berryman, E.; Moalli, M.R. Comparative bone anatomy of commonly used laboratory animals: Implications for drug discovery. Comp. Med. 2011, 61, 76–85. [Google Scholar] [PubMed]







| Collagen Source | Acid Solvent | Incubation Time (Hours) | Concentration (M) | Incubation Temperature (°C) | Ratio of Source to Solvent | Yield (%) |
|---|---|---|---|---|---|---|
| Octopus, outer skin [85] | Acetic acid | 2 times 24 | 0.5 | 4 | 5 | |
| Acetic acid with pepsin | 48 | 0.5 | 4 | 50 | ||
| Cod, skin (minced) [84] | Acetic acid | 24 | 0.1 | 4 | 1:6 | 52 |
| Acetic acid | 24 | 0.25 | 4 | 1:6 | 54 | |
| Acetic acid | 24 | 0.5 | 4 | 1:6 | 59 | |
| Cod, skin (whole) [84] | Acetic acid | 24 | 0.5 | 4 | 1:6 | 20 |
| Citric acid | 24 | 0.5 | 4 | 1:4 to 1:20 | 10 to 25 | |
| Citric acid | 3 times 24 | 0.5 | 4 | 1:4 to 1:20 | 70 to 90 | |
| Freshwater Carp, bones [91] | Acetic acid | 24 | 0.5 | 4 | 20 | |
| Freshwater Carp, scales [91] | Acetic acid | 24 | 0.5 | 4 | 7 | |
| Japanese sea bass, bone [87] | Acetic acid | 72 + 48 | 0.5 | 4 | 41 | |
| Japanese sea bass, skin [87] | Acetic acid | 72 + 48 | 0.5 | 4 | 51 | |
| Freshwater Nile percha (adult), skin [101] | Acetic acid | 16 | 0.5 | 15 | 59 | |
| Freshwater Nile percha (young), skin [101] | Acetic acid | 16 | 0.5 | 15 | 63 | |
| Ocellate puffer fish, skin [86] | Acetic acid | 72 | 0.5 | 4 | 11 | |
| Acetic acid with pepsin | 48 | 0.5 | 4 | 45 | ||
| Sardine, scales [90] | Acetic acid | - | 0.5 | 4 | 5 | |
| Acetic acid with pepsin | 96 | 0.5 | 4 | 14 | ||
| Acetic acid with pepsin | 96 | 0.5 | 15 | 71 | ||
| Shark, cartilage [88] | Acetic acid | - | 0.5 | 4 | 20 | |
| Acetic acid with pepsin | - | 0.5 | 4 | 54 |
| Collagen Source | Solvent | Pre-Treatment | Optimal Pre-Treatment Values | Relation of Yield | ||||
|---|---|---|---|---|---|---|---|---|
| Time t (Hours) | Concentration C (Moles) | Temperature T (°C) | Time (Hours) | Concentration (Moles) | Temperature (°C) | |||
| Grass carp skin [99] | NaOH | 1, 2, 4, 8 or 12 | 0.05, 0.1, 0.2 or 0.5 | 4, 10, 15 and 20 | 12 h | 0.05 and 0.1 M Increased loss of Hydroxyproline Above 0.1 M | 4 and 20 | α t, α C, α T |
| Acetic acid | 72 | 0.1, 0.2, 0.5 or 1.0 | 4, 10, 15 or 20 | 72 | 0.5 | Similar in range 4–20 °C at 0.5 M | α C (<0.5 M), α T (<0.5 M) | |
| Fish myocommata [103] | NaOH | 96 (4 nights) | 0.01, 0.05, 0.1, 0.5, 1 | 5 | 96 | 0.1 | 5 | α C |
| Alaska pollock cold water fish skin [106] | Ca(OH)2 | 0.75 (45 min) and 1.5 | 0.01 and 0.1 mol/L of OH in alkali solvent | 2 and 20 | Not clear | 0.1 | 2 °C was noted for significant loss of collagen | α C |
| Alaska Pollock Skin [105] | NaOH Ca(OH)2 | 1 | 0.01, 0.1, 0.2, and 0.5 mol/L of OH in alkali solvent | 2 to 4 | 1 | 0.01 and 0.1 | 2 to 4 | α C (<0.1) |
| acetic acid | 1 | 0.05 mol/L | 2 to 4 | 1 | 0.05 | 2 to 4 | Not investigated | |
| Arabesque greenling [83] | NaOH, butyl alcohol | 6 (NaOH) 18 (butyl alcohol) 6 | 0.1 M/lit | 4 | Same | Same | Same | Not investigated |
| Salmo salar skin [25] | NaOH Acetic acid Enzyme | 0.16 (10 min) | 0.1 M NaOH 0.005 M Acetic acid | Not given | Not investigated | Not investigated | Not investigated | Not investigated |
| Source | Pre-/Post Ultrasonic Processing/s | Ultrasonic Processing | Yield (%) | Optimal Parameters/Results | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Solvents | Concentration | Temperature (°C) | Time | Ratio of Source to Acid Solution | Power and Amplitude | Intensity | Time | Temperature | Pulse Interval | |||
| Longissimus dorsi muscle from Holstein bulls [110] | Pre-processing KCl, K2HPO4, KH2PO4, EDTA | 100 mM (KCl), 25 mM (K2HPO4) 25 mm (KH2PO4) 2 mM (EDTA) | 4 | Not given | 100, 300 | 20 kHz | 10, 20, 30 (Minutes) | Not given | On-time 2 s Off time 4 s | Not given | 300 Wat and 30 min high pH, Good gel strength, Good emulsification | |
| Cattle tandem [112] | Pre-process Acetic acid Post- process Pepsin with 0.5 M acetic acid | 0.5 M/liter (Acetic acid) 20–70 U mg−1 of tendon (Pepsin + acetic acid) | 4 | 0 to 24 h | 1:15 (w/v) | Low power (amplitude Not given) | 20 kHz | 0 to 24 h | 4 | On-time 20 s Off time 20 s | Based on dry weight 6.2% (24 h of US & 24 h of enzyme acidic) | 5.7% is the Optimal for 12 h of treatment |
| Sea bass skins [111] | Pre-process Acetic acid | 0.5 M/L | 4 | 1:2 (w/v) | Amplitude 20, 40, 60, and 80% | 20 kHz | 0 to 24 h | 4 | On-time 20 s Off time 20 s | 93% | Combined acidic and ultrasonic has better result | |
| Chicken sternal cartilage [32] | Pre-process NaCl, Tris HCl, Na2CO3, EDTA, Isopropyl Alcohol, Acetic acid | 0.05 mol/L (Tris HCl) 0.5 M/L (Na2CO3) 0.3 M/L (EDTA) 0.5 M/L (Acetic acid) | 4 | Overnight (NaCl) 24 h (Tris HCl) 24 h (EDTA) | 1:10 w/v (NaCl) 10% v/v (isopropyl alcohol) 1:6 w/v (Acetic acid) | 950 Amplitude of φ10 | 20–25 kHz | 6, 12, 24, 36 (Minutes) | 4 | On-time 2 s Off time 3 s | Protein contents excluding other ~84% | Higher the treatment time, higher the yield 36 min is the highest |
| Source | Category of Hydrostatic Extraction | Hydrostatic Extraction Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Alkaline and Acidic Followed by Water with Temperature | Water + Temperature Only | Water (Temperature) + Enzyme | Acid Base Gelatin then Hydrostatic | Pre-Acid Treatment Followed by Water Extraction | Gelatin with Only High Pressure | Pressure | Time | Temperature | Yield | |
| Mixed Bovine gelatine and Whey protein (Post) [115] | √ | 6 kbar | 20 min | -- | ||||||
| Fish skin from Cod, fish skin from megrim (post) [116] | √ | 200, 300, 400 (MPa) | 20 °C, 78 °C | -- | ||||||
| Dover sole (Solea vulgaris) skin [117] | √ 0.05 M acetic acid (3 h) | 16–18 h | 45 °C | -- | ||||||
| √ Mild 0.05 M acetic acid (3 h) | 250 MPa | 10, 20 min | 45 °C | 22.8% | ||||||
| Giant squid (Dosidicus gigas) skin [118] | √Skin to water ratio is 1:4 | 12 h | 65 °C | 7.50% | ||||||
| Skins of fresh and cold-smoked salmon, Skins of salted and marinated herrings [119] | √ | 15–120 min | minced skins 45 °C, 70 °C, 100 °C | -- | ||||||
| Backbones and heads of cod [119] | √ | 1st stage 45 min 2nd stage 45 min 3rd stage 45 min | 1st stage 45 °C 2nd stage 60 °C 3rd stage 70 °C | -- | ||||||
| Channel fish skin [120] | √ Calcium hydroxide solution for 68–76 | 5.34–6 | 40 to 46 °C | -- | ||||||
| Pig Skin [21] | √ | 1st stage 7 h 2nd stage 2 h 3rd stage 3 h | 100 °C | -- | ||||||
| √ | same as before then enzymatic extraction for 12 h | 50 °C | -- | |||||||
| Category | Novel Areas to Explore for Future |
|---|---|
| Process | The ultrasound-based extraction at high parameters (intensity, power, amplitude, and time) for various unexplored sources (bovine, porcine). |
| Water extraction along with enzymatic hydrolysis for different species is not yet explored. | |
| Water extraction with combined pre-chemical and post-enzymatic hydrolysis for different species is not yet explored. | |
| Supercritical carbon dioxide is the least researched technique among all extraction processes in terms of combinations with pre- and post-treatments, species, process parameters (pressure, temperature). | |
| Type | Sediment method is only used to extract Type I collagen hydrolysate from a marine source (gutted fish). It is not yet explored for other types of collagen (Type II and III) and various other species (bovine, chicken, porcine). |
| Glucosamine negative effects along with collagen hydrolysate are confusing and need dedicated research for clarifications. | |
| Species | The effects of collagen hydrolysate are not thoroughly investigated for rabbits, cats, and dogs. |
| Collagen hydrolysate extracted from Amur sturgeon (swim bladders) does not accumulate in the kidney and may lead to controlled urinary secretions in cats. This needs a separate area of research to discover the potential of novel sources of collagen hydrolysate in cats. | |
| Different chicken organs (cartilage, heads, skin) as a collagen hydrolysate species are not yet investigated for wound healing, cognitive repair, and antioxidation. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris, M.; Potgieter, J.; Ishfaq, K.; Shahzad, M. Developments for Collagen Hydrolysate in Biological, Biochemical, and Biomedical Domains: A Comprehensive Review. Materials 2021, 14, 2806. https://doi.org/10.3390/ma14112806
Harris M, Potgieter J, Ishfaq K, Shahzad M. Developments for Collagen Hydrolysate in Biological, Biochemical, and Biomedical Domains: A Comprehensive Review. Materials. 2021; 14(11):2806. https://doi.org/10.3390/ma14112806
Chicago/Turabian StyleHarris, Muhammad, Johan Potgieter, Kashif Ishfaq, and Muhammad Shahzad. 2021. "Developments for Collagen Hydrolysate in Biological, Biochemical, and Biomedical Domains: A Comprehensive Review" Materials 14, no. 11: 2806. https://doi.org/10.3390/ma14112806
APA StyleHarris, M., Potgieter, J., Ishfaq, K., & Shahzad, M. (2021). Developments for Collagen Hydrolysate in Biological, Biochemical, and Biomedical Domains: A Comprehensive Review. Materials, 14(11), 2806. https://doi.org/10.3390/ma14112806