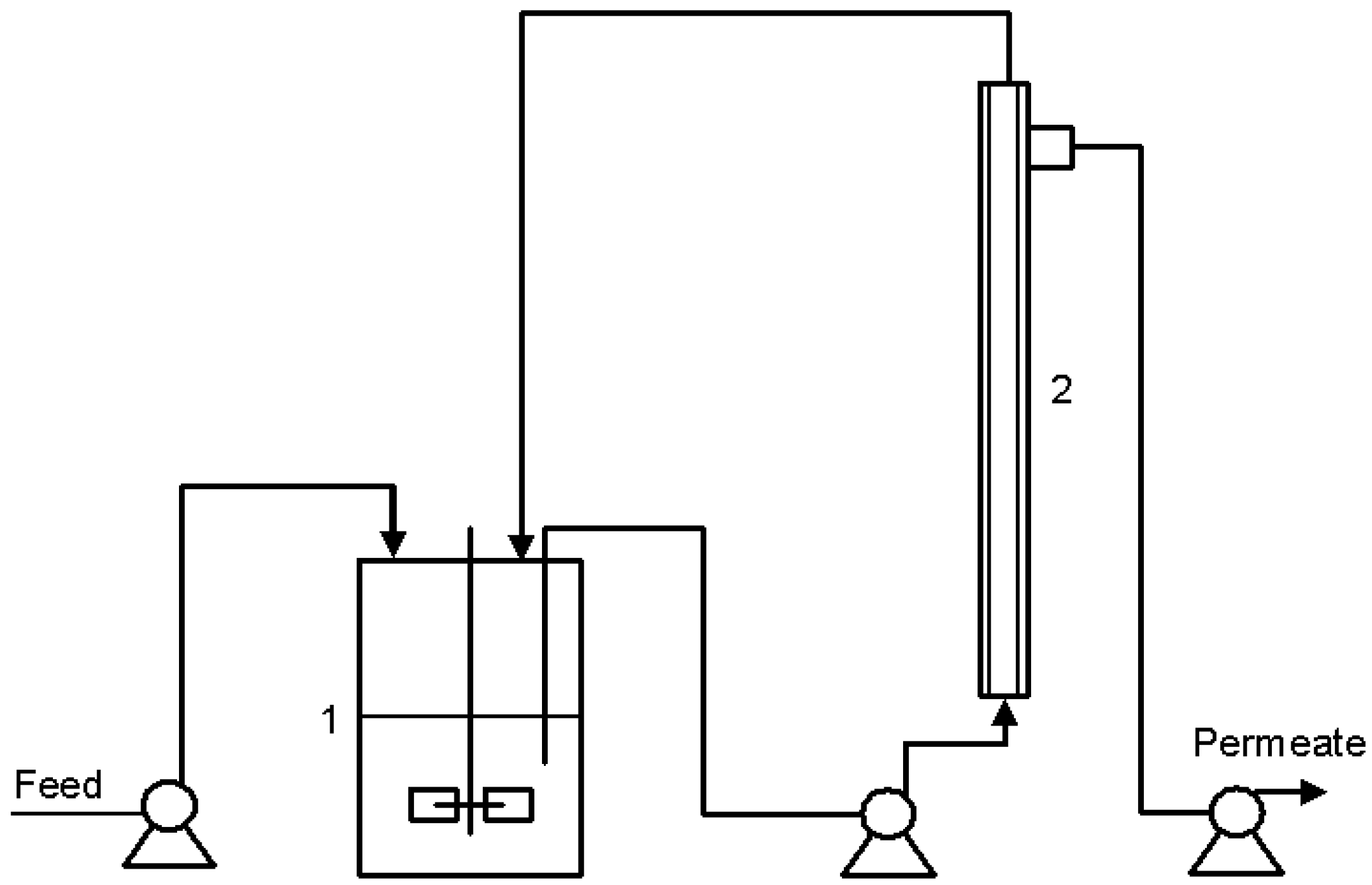
3.1. The Cross Flow Membrane
Cross-flow filtration is ideal for industrial-scale separation, purification, concentration and clarification processes in the biological, medical, food, dairy, beverage and pharmaceutical industries. Membrane fouling is the main difficulty, which can be dealt with by applying high cross-flow velocities and membrane cleaning [
20,
21]. The cross flow membrane used during this study was run with no fouling problems during one entire week without any chemical cleaning. Physical cleaning by relaxing the membrane was employed in order to avoid fouling. During the entire study, the permeate from the cross-flow unit was entirely transparent and free of suspended yeast.
One observation made during the cultivations was that the tubular cross-flow membrane was affected by the furfural concentration in the broth. As the furfural concentration was increased to 10 g L−1 and above, the tubular membranes started to elongate and twist inside the module. However, no tubular membranes broke during any of the cultivations and the permeate from the filter was always transparent and essentially free from yeast. As medium containing less furfural was supplied to the system, the tubular membrane retained its original shape. Because of the problem with the elongation of the membranes and the risk of blocking the 0.9 mm diameter tubular holes, it was decided not to increase the furfural to more than 20.6 g L−1.
The membranes used in the module applied performed ultrafiltration and had a pore size of about 0.02 µm. An ultrafiltration module was applied since some studies have shown that ultrafiltration modules have a lower fouling rate than microfiltration modules [
22,
23].
3.2. Anaerobic Pulse Addition Experiments
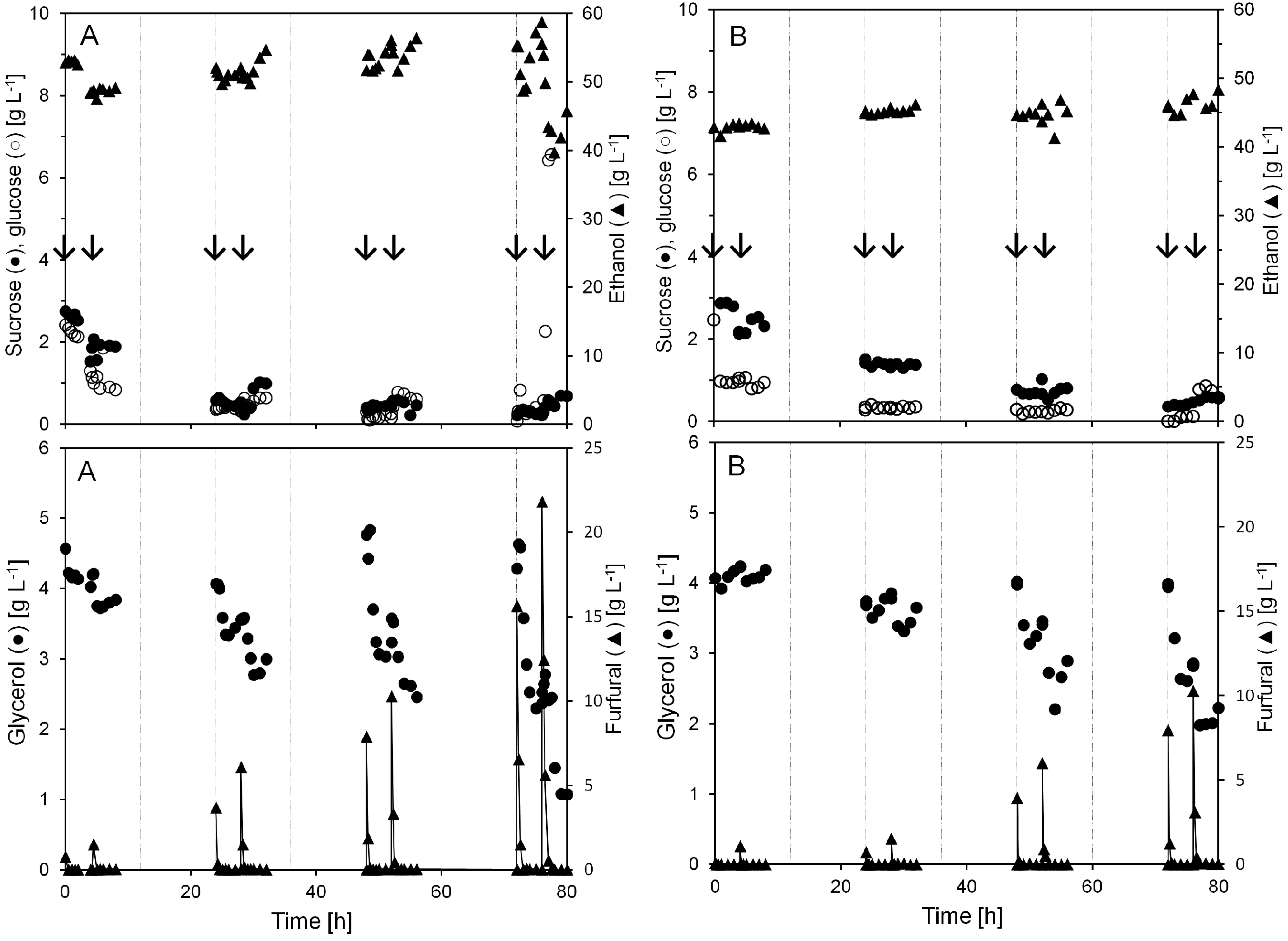
The anaerobic conversion of furfural and its effect on ethanol production was studied by pulse injection of furfural to a continuous cultivation of yeast at high biomass concentration. By injecting furfural directly inside the bioreactor, the furfural concentration could be raised very quickly in the broth, and the effect on the cultivation could be evaluated. Each day, the cultivation was exposed to two pulse injections of different amounts of furfural, once in the morning and once four hours after the first injection. Two separate cultivations were performed and the results for both cultivations are summarized in
Figure 2. The product yields and carbon balance for both cultivations are presented in
Table 1. During pulse injections of furfural the ethanol yield on sucrose (Y
p/s) was stable, and had an average value of 0.49 ± 0.02. Hence, the production of ethanol was not influenced to any larger extent by the injection of furfural.
Table 1.
Product yields and carbon balance during anaerobic continuous cultivations with either pulse injection or continuous furfural addition. The indicated furfural concentration is either the amount of furfural pulse injected to the cultivation or the level of furfural in the feed medium.
Table 1.
Product yields and carbon balance during anaerobic continuous cultivations with either pulse injection or continuous furfural addition. The indicated furfural concentration is either the amount of furfural pulse injected to the cultivation or the level of furfural in the feed medium.
| Furfural pulse injections | | Furfural in feed medium |
|---|
| Furfural [g L−1] | Yp/s c | Ygly/s c | Carbon balance e | Furfural [g L−1] | Yp/s d | Ygly/s d | Carbon balance e |
| 0.1 b | 0.45 | 0.041 | 0.93 | 4.1 b | 0.52 | 0.018 | 0.99 ± 0.02 |
| 0.5 b | 0.45 | 0.042 | 0.96 | 5.7 b | 0.53 | 0.013 | 0.99 ± 0.02 |
| 0.8 ± 0.1 a,b | 0.48 ± 0.04 | 0.039 ± 0.004 | 1.01 ±0.05 | 6.7 a | 0.51 | 0.015 | 1.03 ± 0.06 |
| 1.5 ± 0.0 a,b | 0.48 ± 0.04 | 0.037 ± 0.003 | 0.98 ± 0.00 | 7.8 b | 0.50 | 0.013 | 0.93 ± 0.01 |
| 3.9 ± 0.2 a,b | 0.49 ± 0.04 | 0.039 ± 0.004 | 0.99 ± 0.02 | 9.0 b | 0.53 | 0.011 | 1.00 ± 0.01 |
| 6.1 ± 0.1 a,b | 0.49 ± 0.03 | 0.031 ± 0.003 | 0.98 ± 0.02 | 9.6 a | 0.53 | 0.009 | 1.01 ± 0.05 |
| 7.8 ± 0.1 a,b | 0.50 ± 0.04 | 0.036 ± 0.002 | 1.00 ± 0.00 | 14.4 b | 0.50 | 0.013 | 0.90 ± 0.04 |
| 10.3 ± 0.0 a,b | 0.51 ± 0.03 | 0.026 ± 0.007 | 1.01 ± 0.01 | 17.0 a | 0.53 | 0.010 | 1.02 ± 0.03 |
| 15.6 a | 0.52 | 0.036 | 0.95 | 18.6 b | 0.30 | 0.021 | 0.63 ± 0.23 |
| 21.8 a | 0.49 | 0.026 | 0.90 | 20.6 a | 0.19 | 0.017 | 0.79 ± 0.21 |
At the lower furfural additions of 0.8 and 1.5 g L−1, ethanol and glycerol concentrations in the broth decreased less than 10%. However, as larger amounts of furfural were injected to the cultivation, ethanol and glycerol concentration decreased up to around 20% and 60%. Ethanol and glycerol production rates recovered after the furfural had been converted or eluted from the cultivation broth. A faster recovery occurred in the ethanol production rate compared to the glycerol production rate.
Furfural conversion has earlier been shown to compensate for the production of glycerol [
8]. Under anaerobic conditions,
S. cerevisiae produces glycerol in order to re-oxidize the surplus cytosolic NADH in order to maintain the redox balance in the cell. As the reduction of furfural consumes NADH, it prevents the formation of the same molar amount of glycerol. This will therefore lower the concentration of glycerol while the furfural is converted. The reduced glycerol production was clearly observed, especially when large amounts of furfural were added (
Figure 2). However, after furfural was depleted from the cultivation broth, the glycerol production slowly increased again.
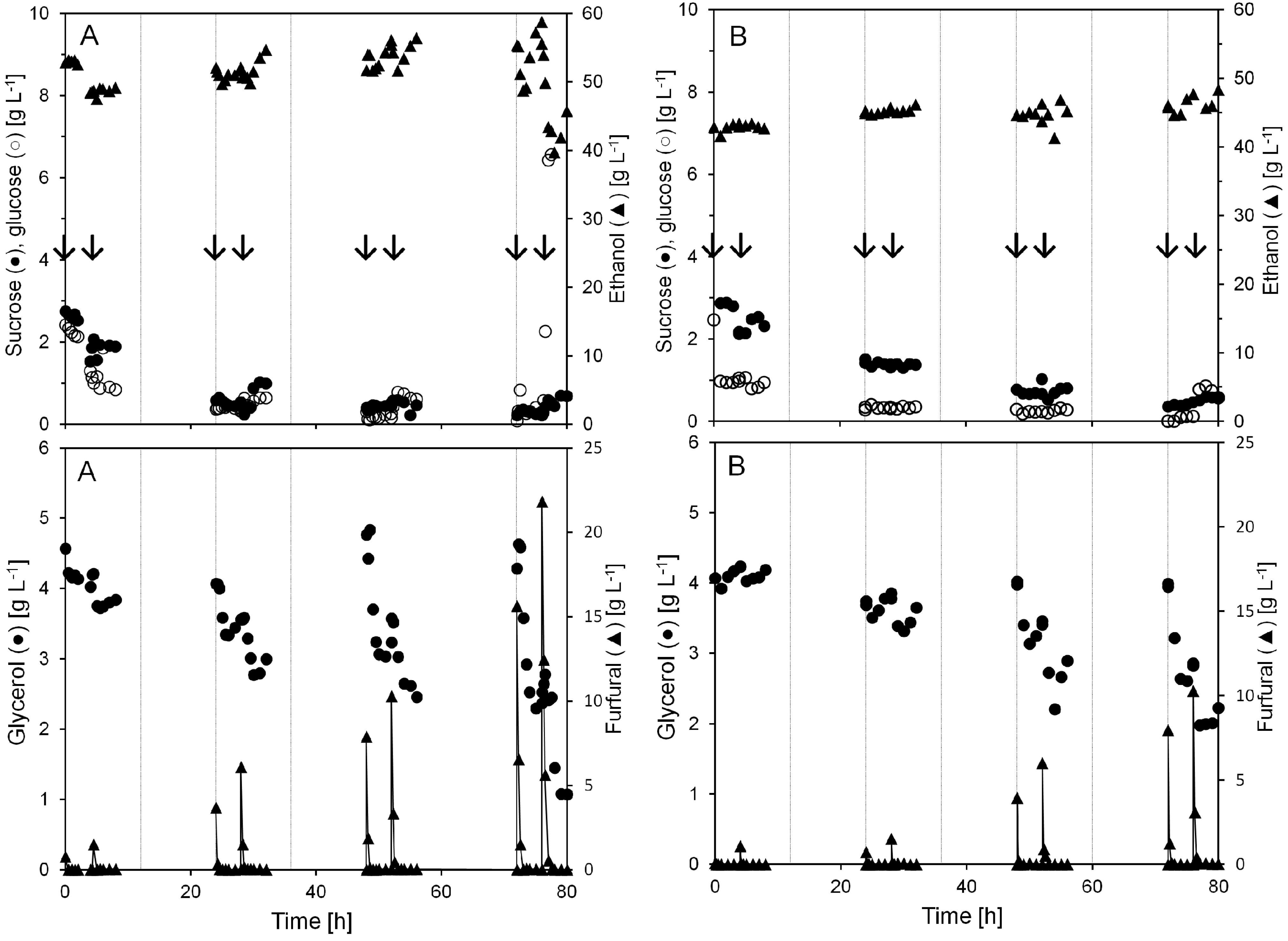
Figure 2.
Concentrations of sucrose, glucose, ethanol, glycerol, and furfural during two anaerobic continuous cultivations with pulse injection of furfural (A and B). Each pulse injection of furfural is indicated by arrows. The gridlines denote each 12 h period in the continuous cultivation.
Figure 2.
Concentrations of sucrose, glucose, ethanol, glycerol, and furfural during two anaerobic continuous cultivations with pulse injection of furfural (A and B). Each pulse injection of furfural is indicated by arrows. The gridlines denote each 12 h period in the continuous cultivation.
Pulse injection of 15.6 and 21.8 g L−1 furfural after 72 h in cultivation A caused the ethanol concentration to decline from 55 to below 40 g L−1 and the glycerol from 4.6 g L−1 to 1 g L−1. At the same time the glucose level rose from almost 0 to above 10 g L−1. Nevertheless, furfural conversion started immediately after the injection and at all the different pulse additions the furfural was gone from the cultivation broth within 1 h after the addition.
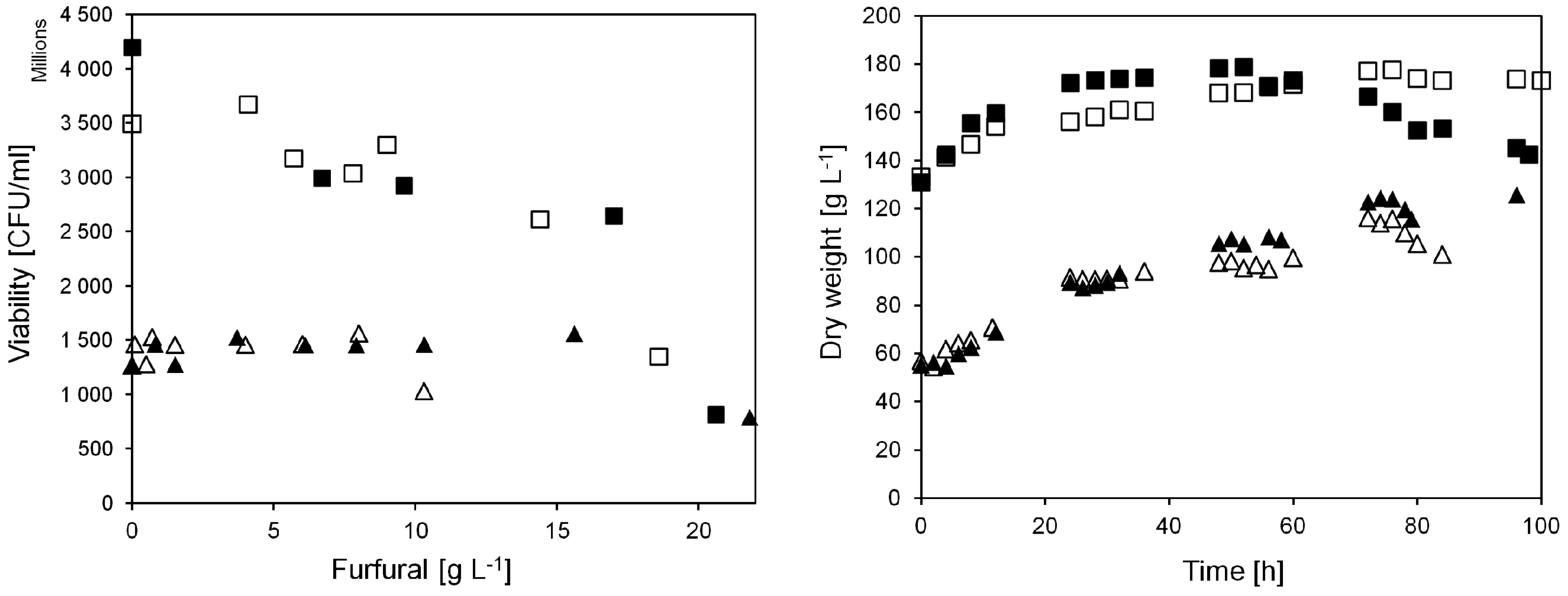
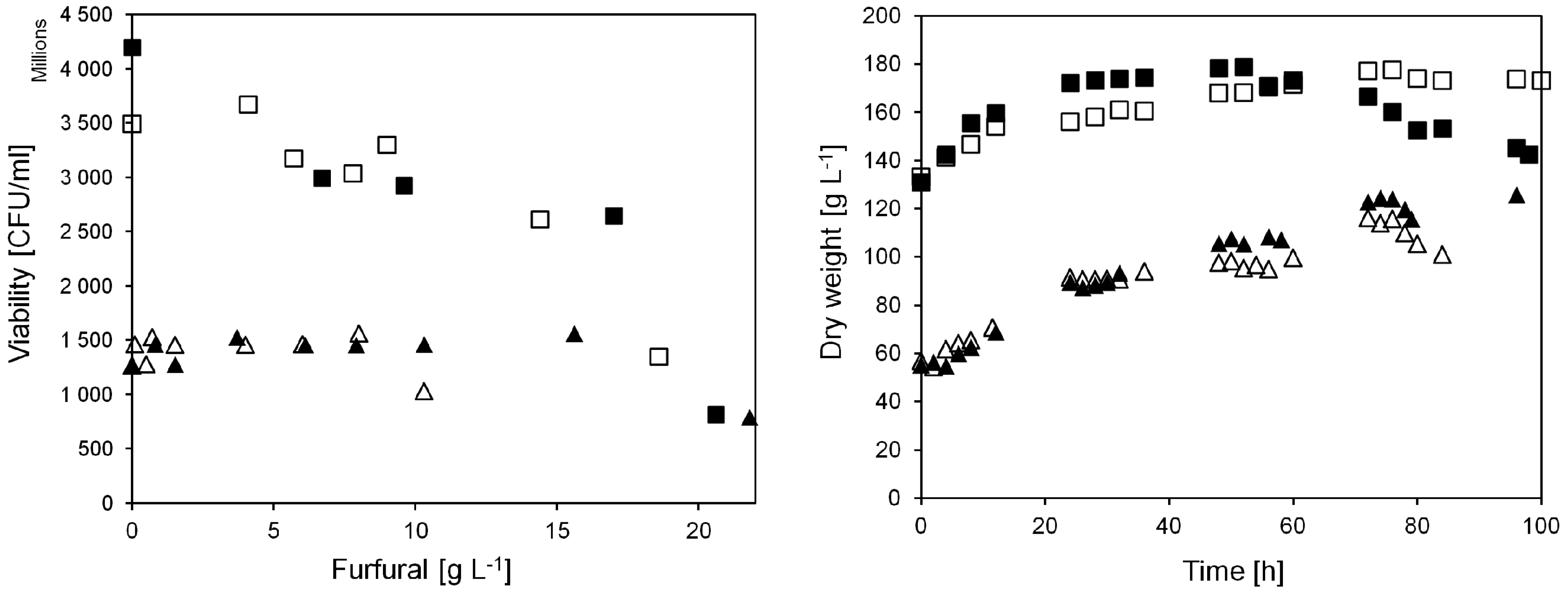
When furfural was injected to the culture in the current work, the yeast dry weight in the bioreactor was about 60 g L
−1. After 72 h of cultivation it had doubled to approximately 120 g L
−1 (
Figure 3). In addition, the viability of the yeast was stable at around 1.5 × 10
9 CFU mL
−1. This shows that the pulse injection of furfural did not severely affect the yeast viability in the cultivation until 72 h. After 76 h when furfural addition was increased to 21.8 g L
−1 (A), a decrease in yeast viability was observed, down to 0.8 × 10
9 CFU mL
−1. A decreasing yeast biomass concentration was also observed. This was caused by a lack of cell growth, and because of sampling, since biomass was collected several times during the cultivation.
Even at a yeast biomass concentration of 60 g L
−1 during the first 24 h there were residual sugars left in the permeate. The level of residual sugars decreased to almost zero, as the yeast biomass concentration increased along the cultivation. However, when 21.8 g L
−1 (A) furfural was injected the residual sugar concentration drastically increased to 11 g L
−1 (
Figure 2).
Figure 3.
Yeast viability and biomass dry weight at different furfural additions in anaerobic continuous cultivations on sucrose. Triangular symbols denote pulse injection experiments, viability measured 4 h after furfural injection. Square symbols denote experiments with continuous furfural addition, viability measured after 24 h continuous addition of furfural at the indicated concentrations. Closed and open symbols denote cultivation A and B, respectively.
Figure 3.
Yeast viability and biomass dry weight at different furfural additions in anaerobic continuous cultivations on sucrose. Triangular symbols denote pulse injection experiments, viability measured 4 h after furfural injection. Square symbols denote experiments with continuous furfural addition, viability measured after 24 h continuous addition of furfural at the indicated concentrations. Closed and open symbols denote cultivation A and B, respectively.
3.3. Anaerobic Continuous Cultivations with Furfural Containing Inlet Medium
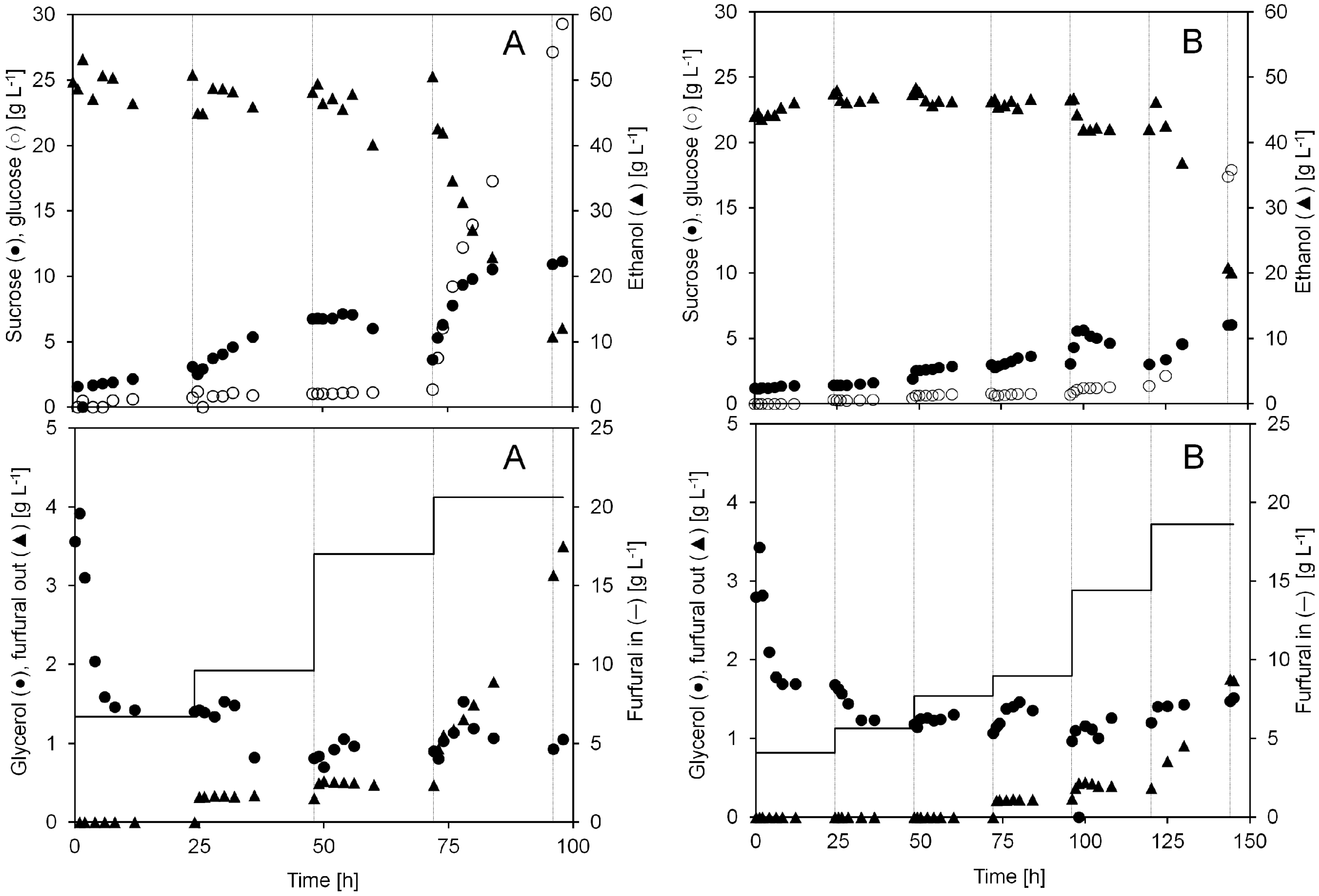
The result from the continuous cultivation with furfural addition to the medium is summarized in
Figure 4 and
Table 1. In this experiment, more residual sugars were left unfermented in the spent liquid in contrast to when furfural pulse injections were performed. The level of sucrose increased during the entire cultivation (A) from 2 to 11 g L
−1. The glucose concentration stayed below 2 g L
−1 during the first 72 h, but increased to 29 g L
−1 after adding 20.6 g L
−1 furfural to the cultivation for 24 h. The same trends were seen in cultivation B.
The cell viability remained above 2.5 × 10
9 CFU mL
−1, except when the furfural concentration was increased to either 18.6 (B) or 20.6 (A) g L
−1 in the feed (
Figure 3). After feeding 18.6 or 20.6 g L
−1 furfural for 24 h, the cell viability had decreased by 52% or 68% to only 1.35 × 10
9 or 0.8 × 10
9 CFU mL
−1. During this time, an increasing amount of furfural in the spent liquid was observed, indicating that the inflow of furfural was higher than the yeasts´
in situ detoxification capacity for furfural. Throughout most of the cultivation, the furfural level in the spent medium was less than 0.5 g L
−1. When the concentration of furfural was increased to 18.6 (B) or 20.6 (A) g L
−1, the outflow of the bioreactor contained a rising concentration of furfural. Apparently, the yeast could convert almost all furfural as long as the amount of furfural did not exceed a certain level.
At a constant furfural addition of 6.7, 9.6 and 17 g L−1 (A) or 4.1, 5.7, 7.8, and 9.0 g L−1 (B) the ethanol concentration remained stable. When adding 18.6 (B) or 20.6 (A) g L−1 furfural the conversion capacity of the yeast was not high enough and the furfural concentration reached a critical level which made the yeast unable to maintain a stable ethanol production. During the following hours from 72 to 98 h (A) the ethanol concentration in the bioreactor decreased from 50 to 11 g L−1.
Figure 4.
Concentrations of sucrose, glucose, ethanol, glycerol and furfural during two anaerobic continuous cultivations with continuous furfural addition (A and B). The furfural concentration (g L−1) in the inlet medium and the permeate during each cultivation day is shown in the lower graphs. After each 24 h period, denoted by the gray gridlines in the graphs, the furfural concentration in the inlet medium was increased to the indicated concentration.
Figure 4.
Concentrations of sucrose, glucose, ethanol, glycerol and furfural during two anaerobic continuous cultivations with continuous furfural addition (A and B). The furfural concentration (g L−1) in the inlet medium and the permeate during each cultivation day is shown in the lower graphs. After each 24 h period, denoted by the gray gridlines in the graphs, the furfural concentration in the inlet medium was increased to the indicated concentration.
The maximal cell concentration reached was 180 g L
−1, although, after 72 h when adding 17.0 g L
−1 (A) furfural into the cultivation this value started to decline in cultivation A. The biomass amount inside the reactor decreased not only due to biomass sampling and lack of cell growth but also because of reduced cell size and/or cell lysis. Interestingly, the concentration of glycerol in the broth was below 2 g L
−1 except during the first 4 h. This is probably due to the same effect discussed above, since furfural conversion prevents the formation of glycerol. However, glycerol production never stopped entirely, glycerol was always present at low levels in the broth. At an inlet furfural concentrations of 18.6 g/L or higher, the furfural concentrations in the reactor became inhibitory. The increased stress under such conditions may very well lead to increased glycerol yield as part of the general stress response. Hypothetically, the increased sugar concentrations and the very high cell density may have triggered responses to osmotic stress, something which is well known to induce glycerol production [
24].
3.4. Cell Retention and Furfural Conversion
When furfural is added to high broth concentrations, the yeast growth is strongly inhibited and this may lead to washout in a traditional continuous cultivation without cell retention [
7]. However, when a membrane filtration unit is used for total cell retention, washout will not occur even if the cells are not growing, since all cells are kept inside the bioreactor by the membranes even at a dilution rate of 0.5 h
−1. This gives good advantages since the cells do not have to grow to be retained inside the bioreactor. The combination of eliminated washout and high cell concentrations makes cell retention methods especially suitable for fermentation of sugars at high dilution rates and higher productivity [
17]. Immobilizing the yeast has also been proven as a successful method for fermenting toxic media, e.g., lignocellulosic hydrolyzate, at higher dilution rates of 0.3–0.5 h
−1 [
25,
26]. However, several practical problems with immobilization exist, which make immobilization complicated to use at industrial scale.
It has been confirmed [
7,
27,
28] that the NADH-dependent enzyme alcohol dehydrogenase, which catalyzes the reduction of acetaldehyde to ethanol, additionally converts furfural to furfuryl alcohol under anaerobic conditions. This metabolic biotransformation is referred to as
in situ detoxification since a less toxic product is created in the conversion. Since the same enzyme is used both for acetaldehyde as for furfural conversion, the production rate of ethanol and ATP would be lowered if the enzyme capacity becomes limiting. Reduced energy supply in the cells can cause, e.g., reduced biomass production. This would explain why batch cultivations with added furfural result in reduced biomass yield, lowered specific growth rate and ethanol productivity [
6,
8,
29].
A prerequisite to maintain the cells’ ability to perform
in situ detoxification is to select a suitable operation mode for the cultivation. During continuous and fed-batch cultivations,
S. cerevisiae is more tolerant to furfural, because high concentrations of inhibitors can be avoided in the cultivation broth if the feeding rate is appropriate. In a continuous cultivation without yeast retention no negative effects on either ethanol or biomass yield appeared at a dilution rate of 0.315 h
−1 with 2.0 g L
−1 furfural in the inlet medium, while 4.3 g L
−1 furfural resulted in washout [
7]. However, in this study, the yeast successfully produced ~47 g L
−1 ethanol at an inlet medium level of 17.0 g L
−1 furfural at a dilution rate of 0.5 h
−1.
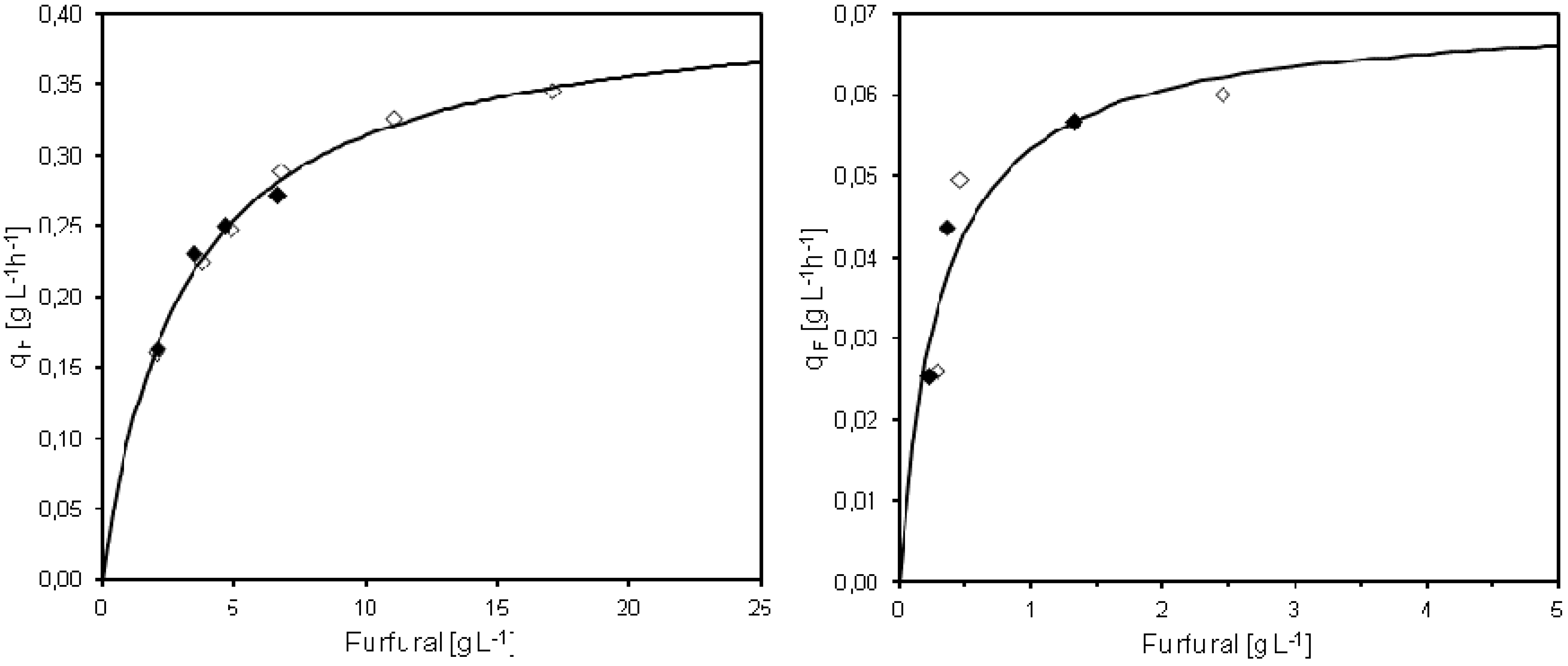
3.5. Kinetics of Furfural Conversion
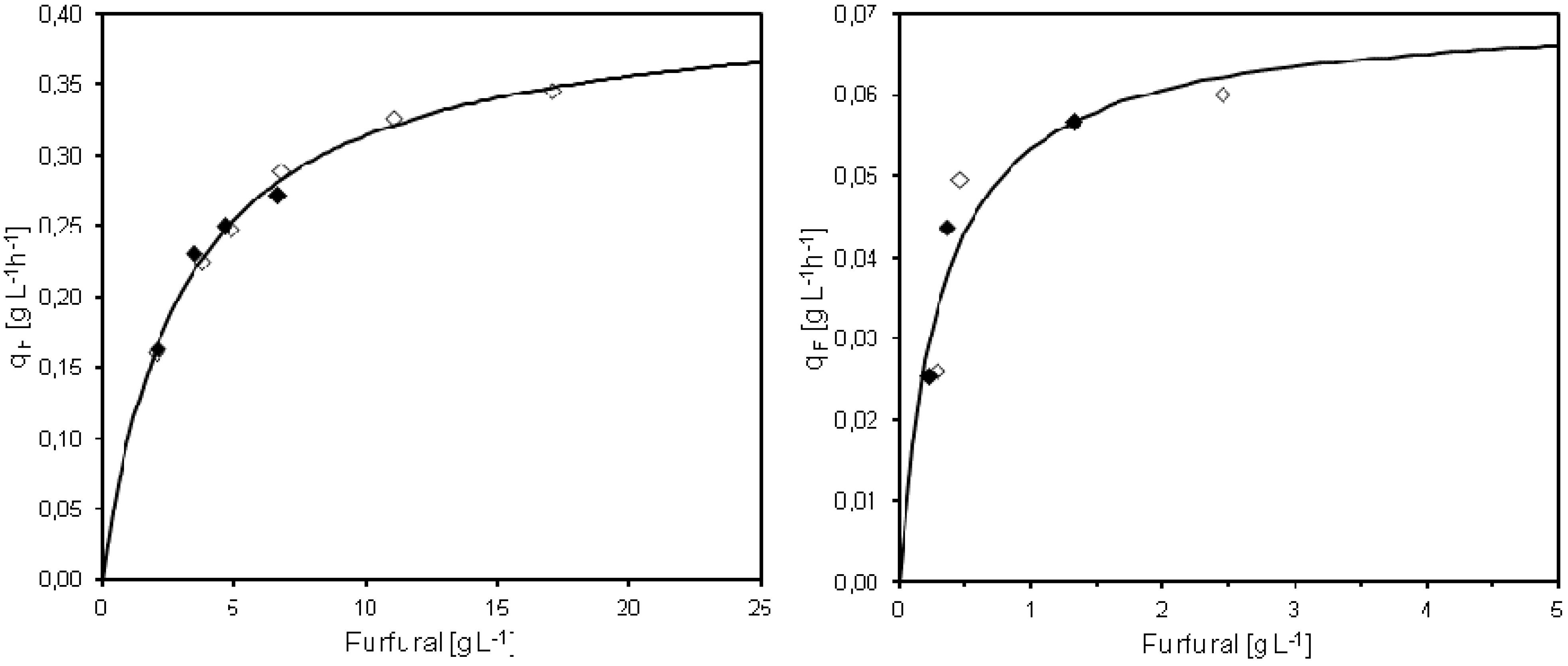
Michaelis-Menten saturation kinetics was used in order to describe the specific furfural conversion rate based on results from the pulse injection or continuous furfural addition experiments in MATLAB
®. The maximum specific conversion rate of furfural (q
max) and the Michaelis-Menten constant (K
m) for furfural pulse injections was determined as q
max = 0.41 g L
−1 h
−1 and K
m = 3.1 g L
−1 (
Figure 5), in the same way as earlier described by Sárvári Horváth
et al. [
7]. The estimated specific furfural conversion rate was 0.35 g g
−1 h
−1 within 15 min after pulse injection of 21.8 g L
−1 furfural to the cultivation. When less than 3.7 g L
−1 furfural was added, the conversion rate could not be calculated as all furfural had been converted in less than 15 min.
The calculated q
max of 0.41 g L
−1 h
−1 during furfural pulse injections was lower than previously reported maximum rates. However, unlike previous investigations the yeast in the present study was cultivated at high cell density, the yeast grew slowly and behaved more like stationary phase cells. In previous studies, furfural was pulsed either to exponentially growing yeast in batch cultivations [
8] or into ordinary chemostat cultures at steady-state, without cell retention [
7]. Sárvári Horváth
et al. [
7] presented a maximum specific furfural conversion rate of around 0.6 g g
−1 h
−1, determined during pulse addition of furfural for yeast growing in anaerobic conditions at a dilution rate of 0.10 h
−1.
Michaelis-Menten saturation kinetics was fitted also to the observed specific furfural conversion rates and steady state furfural concentrations obtained at different feed concentrations of furfural. In this case, the fitted q
max was 0.07 g L
−1 and K
m was 0.25 g L
−1 h
−1 (
Figure 5). The results of the furfural conversion during continuous furfural addition thereby showed completely different saturation kinetics compared to the pulse injections experiments. Without cell retention, Sárvári Horváth
et al. [
7] showed that the specific conversion rate of furfural increases with increasing dilution rate until washout with a maximum value of 0.15 g g
−1 h
−1. Thus, growing yeast appears to convert furfural at faster specific rates compared to the slow growing cells in the membrane bioreactor. However, unlike in the Sárvári Horváth
et al. study, washout could not occur in our experiment and a much faster total volumetric furfural conversion rate could be reached due to the high cell density.
Figure 5.
Summary of the specific conversion rate of furfural (qF) in cultivations with pulse injection (left) or continuous (right) furfural addition. The qF values for two separate cultivations are presented together with the regression curve calculated based on Michaelis-Menten kinetics. Open/closed symbols denote separate experiments.
Figure 5.
Summary of the specific conversion rate of furfural (qF) in cultivations with pulse injection (left) or continuous (right) furfural addition. The qF values for two separate cultivations are presented together with the regression curve calculated based on Michaelis-Menten kinetics. Open/closed symbols denote separate experiments.
Interestingly, furfural seems to not have a direct toxic effect on the cells, since the cells recovered even when over 15 g L
−1 was pulsed into the cultivation. The cells could also survive several days with constant addition of furfural, although the cell growth was affected negatively at a furfural concentration over 10 g L
−1. A previous study on the proteomic response of
S. cerevisiae to furfural addition revealed that the expressions of most proteins did not change 20 min after furfural pulse injection [
30]. However, after 2 h, 50 proteins demonstrated up or downregulation. On the other hand, pulse addition of furfural has been shown to induce a large transcriptome change in
S. cerevisiae within 20 minutes [
31]. For example, genes controlling transcription and translation are downregulated which will affect cell growth. Several stress response genes, encoding mitochondrial proteins, are also upregulated [
31]. These observations indicate, that when furfural is continuously added to the cultures, there is probably a transcriptional response which affects the furfural-converting enzymes, while during the pulse injections, no major changes occur before the added furfural is consumed. This can explain why two different furfural conversion kinetics were observed in our study depending on if furfural was pulse injected or fed continuously to the cultivation.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}