
Exploring the Pivotal Significance of Microalgae-Derived Sustainable Lipid Production: A Critical Review of Green Bioenergy Development

 , , , and
, , , and 
Abstract
1. Introduction
1.1. Microalgae vs. Plants
- i.
- Under photoautotrophic conditions, microalgae maintain a simple unicellular structural configuration that allows the entire biomass to be photosynthetically reactive without the use of heterotrophic organelles. They are naturally occurring and exist across a variety of habitats from fresh water to marine without regard for a seasonal life cycle.
- ii.
- The efficient metabolic activity of microalgal cells is influenced by the availability of several trophic modes, namely photoautotrophic, mixotrophic, and heterotrophic, which are dependent on physicochemical balance and environmental adaptation. Over intrinsic photoautotrophy, the organic carbon and light-interceded mixotrophic cultivation conditions provide a supplementary benefit to the entire microalgal structure for increasing biomass and accumulation of essential by-products. As a result, microalgae have a quicker doubling time and a flexible metabolism.
- iii.
- Microalgae are not involved in the food vs. fuel debate because they can grow in both non-fertile land and wastewater. Accordingly, the use of microalgae would evolve as an environmentally sustainable option to plant-based crops.
- iv.
- Different microalgal strains can be designated for specific growth conditions, which are suitable and easily adaptable to terrestrial climatic behaviour, which is portrayed as more difficult with conventional crops.
1.2. Microalgae Biorefinery
2. Overview of Research Status of Bioenergy Generation
2.1. International Status
2.2. National Status
3. Photosynthetic Carbon Sequestration Efficiency in Microalgae
3.1. Microalgae Based Composition
3.2. Factors Influencing Microalgae Growth
3.2.1. Nutrients
3.2.2. Culture Conditions
- (a)
- Light and Temperature
- (b)
- Magnetic fields
- (c)
- pH and salinity
3.2.3. Operational Mode
3.2.4. Reactor Types
4. Microalgae Lipids—Production and Relevance
4.1. Technologies in Lipid Extraction
4.1.1. Traditional Methods: Folch and Bligh-Dyer
- (a)
- Folch Method
- (b)
- Bligh and Dyer
4.1.2. Accelerated Solvent Extraction Procedure (ASE)
- Organic solvents become much less viscous at high temperatures, increasing solvent diffusion [96];
- As the temperature rises from 20 to 150 °C, the diffusion rate increases [95];
- The high pressure of the entire ASE-related lipid-extraction system emphasises the organic solvents’ deep penetration into the cell membrane, causing cell disruption;
- A smaller volume of organic solvent and a shorter extraction time are both highly advantageous for the ASE technique.
4.1.3. Supercritical Fluids
- Easy and fast penetration into solid matrices to release the lipid biomolecules;
- Extracted or released lipids will be recovered rapidly by the evaporation of SC-CO2 at the gaseous stage via depressurization, yielding a solvent-free lipid.
4.1.4. Soxhlet Extraction of Lipids
4.2. Lipid Processing and Conversion to Fuel
4.2.1. Thermochemical Conversion
- (a)
- Combustion process
- (b)
- Pyrolysis
- (c)
- Gasification
4.2.2. Biochemical Conversion
- (a)
- Transesterification
- (b)
- Anaerobic digestion (AD)
5. Biochemical Conversion for Bioenergy Production
5.1. Biodiesel
5.2. Bioethanol
5.3. Biohydrogen
5.4. Bio Oil
5.5. Bio Jet Fuel
6. Energy Kinetics—TGA
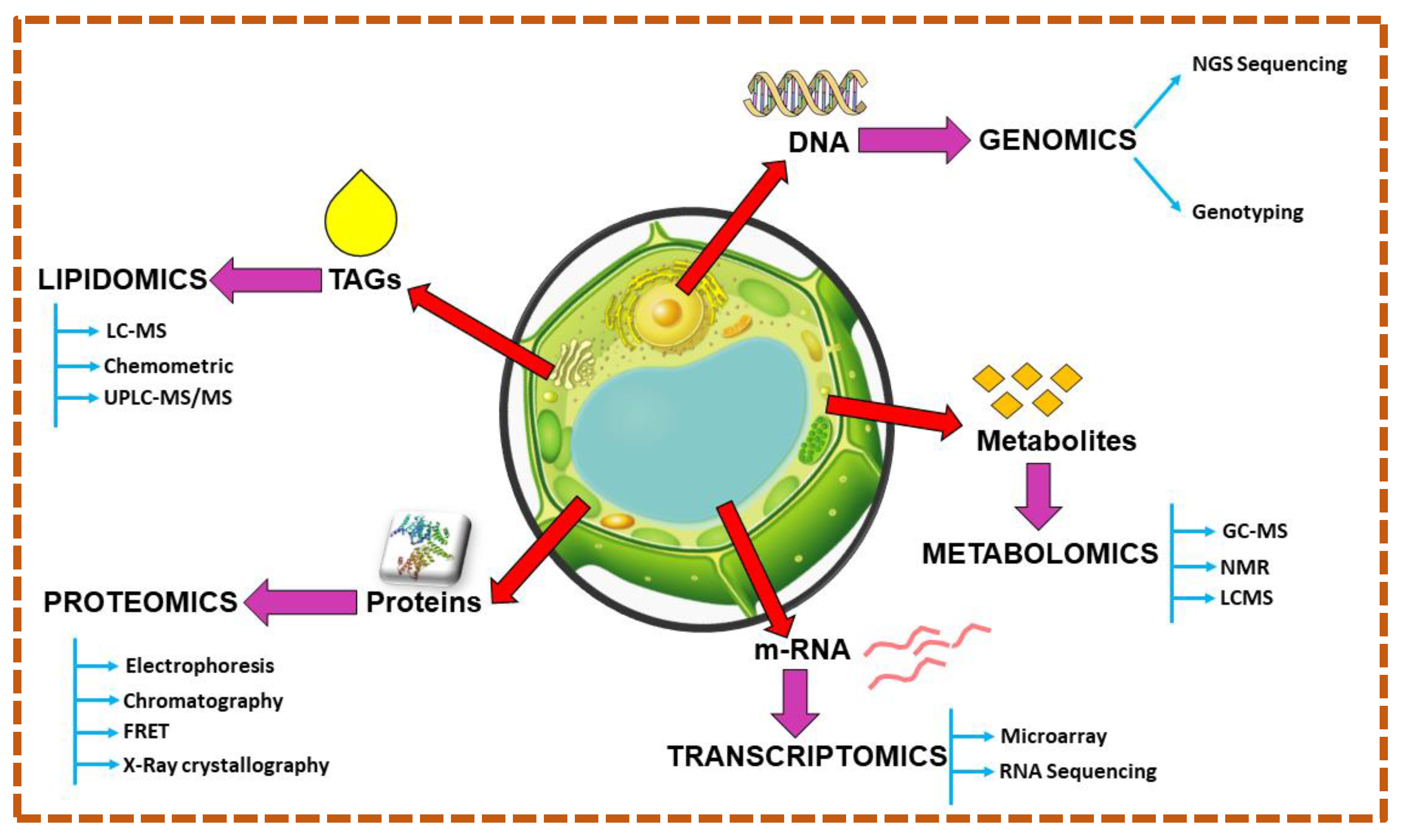
7. Algomics Approach for Microalgae-Based Bioenergy Synthesis
7.1. Genomics
7.2. Proteomics
7.3. Transcriptomics
7.4. Metabolomics
7.5. In Silico Metabolic Approach
8. Economic Aspect of Microalgae-Derived Biofuels
8.1. Life Cycle Assessment (LCA)
8.2. Economics of Microalgae Based Biofuels
9. Research Needs and Future Implications
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khanra, A.; Vasistha, S.; Rai, M.P.; Cheah, W.Y.; Khoo, K.S.; Chew, K.W.; Chuah, L.F.; Show, P.L. Green bioprocessing and applications of microalgae-derived biopolymers as a renewable feedstock: Circular bioeconomy approach. Environ. Technol. Innov. 2022, 28, 102872. [Google Scholar] [CrossRef]
- Vasistha, S.; Khanra, A.; Clifford, M.; Rai, M.P. Current advances in microalgae harvesting and lipid extraction processes for improved biodiesel production: A review. Renew. Sust. Energ. Rev. 2021, 137, 110498. [Google Scholar] [CrossRef]
- Sharma, G.K.; Khan, S.A.; Shrivastava, M.; Gupta, N.; Kumar, S.; Malav, L.C.; Nogiya, M.; Dubey, S.K. Bioremediation of sewage wastewater through microalgae (Chlorella minutissima). Indian J. Agric. Sci. 2020, 90, 2024–2028. [Google Scholar] [CrossRef]
- Rai, M.P.; Gupta, S. Effect of media composition and light supply on biomass, lipid content and FAME profile for quality biofuel production from Scenedesmus abundans. Energy Convers. Manag. 2017, 141, 85–92. [Google Scholar] [CrossRef]
- Yadav, K.K.; Krishnan, S.; Gupta, N.; Prasad, S.; Amin, M.A.; Cabral-Pinto, M.M.S.; Sharma, G.K.; Marzouki, R.; Jeon, B.-H.; Kumar, S.; et al. Review on Evaluation of Renewable Bioenergy Potential for Sustainable Development: Bright Future in Energy Practice in India. ACS Sustain. Chem. Eng. 2021, 9, 16007–16030. [Google Scholar] [CrossRef]
- Khan, M.; Salman, M.; Bashir, U.; Malik, M.S.; Ikram, A. Joint external evaluation of IHR core capacities of the Islamic Republic of Pakistan, 2016. Int. J. Infect. Dis. 2018, 73, 36–37. [Google Scholar] [CrossRef]
- Yukesh Kannah, R.; Kavitha, S.; Parthiba Karthikeyan, O.; Rene, E.R.; Kumar, G.; Rajesh Banu, J. A review on anaerobic digestion of energy and cost effective microalgae pretreatment for biogas production. Bioresour. Technol. 2021, 332, 125055. [Google Scholar] [CrossRef]
- Khan, S.A.; Sharma, G.K.; Malla, F.A.; Kumar, A.; Gupta, N. Microalgae based biofertilizers: A biorefinery approach to phycoremediate wastewater and harvest biodiesel and manure. J. Clean. Prod. 2019, 211, 1412–1419. [Google Scholar] [CrossRef]
- Baldos, U.L.C.; Fuglie, K.O.; Hertel, T.W. The research cost of adapting agriculture to climate change: A global analysis to 2050. Agric. Econ. 2020, 51, 207–220. [Google Scholar] [CrossRef]
- Malla, F.A.; Khan, S.A.; Rashmi; Sharma, G.K.; Gupta, N.; Abraham, G. Phycoremediation potential of Chlorella minutissima on primary and tertiary treated wastewater for nutrient removal and biodiesel production. Ecol. Eng. 2015, 75, 343–349. [Google Scholar] [CrossRef]
- Khanra, A.; Vasistha, S.; Kumar, S.; Rai, M.P. Cultivation of microalgae on unhydrolysed waste molasses syrup using mass cultivation strategy for improved biodiesel. 3 Biotech 2021, 11, 287. [Google Scholar] [CrossRef] [PubMed]
- Pierre, G.; Delattre, C.; Dubessay, P.; Jubeau, S.; Vialleix, C.; Cadoret, J.-P.; Probert, I.; Michaud, P. What Is in Store for EPS Microalgae in the Next Decade? Molecules 2019, 24, 4296. [Google Scholar] [CrossRef] [PubMed]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.; Vecchi, V.; Barera, S.; Dall’Osto, L. Biomass from microalgae: The potential of domestication towards sustainable biofactories. Microb. Cell Factories 2018, 17, 173. [Google Scholar] [CrossRef] [PubMed]
- Raheem, A.; Prinsen, P.; Vuppaladadiyam, A.K.; Zhao, M.; Luque, R. A review on sustainable microalgae based biofuel and bioenergy production: Recent developments. J. Clean. Prod. 2018, 181, 42–59. [Google Scholar] [CrossRef]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.-J.; Chang, J.-S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef]
- Hemalatha, M.; Sravan, J.S.; Min, B.; Venkata Mohan, S. Microalgae-biorefinery with cascading resource recovery design associated to dairy wastewater treatment. Bioresour. Technol. 2019, 284, 424–429. [Google Scholar] [CrossRef]
- Singh, J.; Dhar, D.W. Overview of Carbon Capture Technology: Microalgal Biorefinery Concept and State-of-the-Art. Front. Mar. Sci. 2019, 6, 29. [Google Scholar] [CrossRef]
- Erbland, P.; Caron, S.; Peterson, M.; Alyokhin, A. Design and performance of a low-cost, automated, large-scale photobioreactor for microalgae production. Aquac. Eng. 2020, 90, 102103. [Google Scholar] [CrossRef]
- ElFar, O.A.; Chang, C.-K.; Leong, H.Y.; Peter, A.P.; Chew, K.W.; Show, P.L. Prospects of Industry 5.0 in algae: Customization of production and new advance technology for clean bioenergy generation. Energy Convers. Manag. X 2021, 10, 100048. [Google Scholar] [CrossRef]
- Young, P.; Phasey, J.; Wallis, I.; Vandamme, D.; Fallowfield, H. Autoflocculation of microalgae, via magnesium hydroxide precipitation, in a high rate algal pond treating municipal wastewater in the South Australian Riverland. Algal Res. 2021, 59, 102418. [Google Scholar] [CrossRef]
- Pugazhendhi, A.; Shobana, S.; Bakonyi, P.; Nemestóthy, N.; Xia, A.; Banu, J.R.; Kumar, G. A review on chemical mechanism of microalgae flocculation via polymers. Biotechnol. Rep. 2019, 21, e00302. [Google Scholar] [CrossRef] [PubMed]
- Duarah, P.; Haldar, D.; Patel, A.K.; Dong, C.-D.; Singhania, R.R.; Purkait, M.K. A review on global perspectives of sustainable development in bioenergy generation. Bioresour. Technol. 2022, 348, 126791. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Ke, J.H.; Zhou, S.; Xie, G.H. Estimation of the quantity and availability of forestry residue for bioenergy production in China. Resour. Conserv. Recycl. 2020, 162, 104993. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, P. Focus on bioenergy industry development and energy security in China. Renew. Sust. Energ. Rev. 2014, 32, 302–312. [Google Scholar] [CrossRef]
- Peidong, Z.; Yanli, Y.; Yongsheng, T.; Xutong, Y.; Yongkai, Z.; Yonghong, Z.; Lisheng, W. Bioenergy industries development in China: Dilemma and solution. Renew. Sust. Energ. Rev. 2009, 13, 2571–2579. [Google Scholar] [CrossRef]
- Namsaraev, Z.B.; Gotovtsev, P.M.; Komova, A.V.; Vasilov, R.G. Current status and potential of bioenergy in the Russian Federation. Renew. Sust. Energ. Rev. 2018, 81, 625–634. [Google Scholar] [CrossRef]
- Oecd FAO. OECD-FAO Agricultural Outlook 2022–2031; OECD: Paris, France, 2022. [Google Scholar]
- Energy, E.T. US Energy Information Administration (EIA) Monthly Energy Review; U.S. Department of Energy: Washington, DC, USA, 2021. [Google Scholar]
- Wahab, A.G. Biofuels Annual; MY2020-0013; Office of Agricultural Affairs: Wilayah Persekutuan Kuala Lumpur, Malaysia, 2020. [Google Scholar]
- Hiloidhari, M.; Das, D.; Baruah, D.C. Bioenergy potential from crop residue biomass in India. Renew. Sust. Energ. Rev. 2014, 32, 504–512. [Google Scholar] [CrossRef]
- Lee, X.J.; Ong, H.C.; Gan, Y.Y.; Chen, W.-H.; Mahlia, T.M.I. State of art review on conventional and advanced pyrolysis of macroalgae and microalgae for biochar, bio-oil and bio-syngas production. Energy Convers. Manag. 2020, 210, 112707. [Google Scholar] [CrossRef]
- Hildebrand, M.; Abbriano, R.M.; Polle, J.E.W.; Traller, J.C.; Trentacoste, E.M.; Smith, S.R.; Davis, A.K. Metabolic and cellular organization in evolutionarily diverse microalgae as related to biofuels production. Curr. Opin. Chem. Biol. 2013, 17, 506–514. [Google Scholar] [CrossRef]
- Lim, J.H.K.; Gan, Y.Y.; Ong, H.C.; Lau, B.F.; Chen, W.-H.; Chong, C.T.; Ling, T.C.; Klemeš, J.J. Utilization of microalgae for bio-jet fuel production in the aviation sector: Challenges and perspective. Renew. Sust. Energ. Rev. 2021, 149, 111396. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Zhao, X.-Q.; Yen, H.-W.; Ho, S.-H.; Cheng, C.-L.; Lee, D.-J.; Bai, F.-W.; Chang, J.-S. Microalgae-based carbohydrates for biofuel production. Biochem. Eng. J. 2013, 78, 1–10. [Google Scholar] [CrossRef]
- Yamada, T.; Sakaguchi, K. Comparative studies onChlorella cell walls: Induction of protoplast formation. Arch. Microbiol. 1982, 132, 10–13. [Google Scholar] [CrossRef]
- Muñoz, R.; Gonzalez-Fernandez, C. Microalgae-Based Biofuels and Bioproducts: From Feedstock Cultivation to End-Products; Woodhead Publishing: Sawston, UK, 2017. [Google Scholar]
- de Carvalho Silvello, M.A.; Severo Gonçalves, I.; Patrícia Held Azambuja, S.; Silva Costa, S.; Garcia Pereira Silva, P.; Oliveira Santos, L.; Goldbeck, R. Microalgae-based carbohydrates: A green innovative source of bioenergy. Bioresour. Technol. 2022, 344, 126304. [Google Scholar] [CrossRef] [PubMed]
- Freitas, B.C.B.; Cassuriaga, A.P.A.; Morais, M.G.; Costa, J.A.V. Pentoses and light intensity increase the growth and carbohydrate production and alter the protein profile of Chlorella minutissima. Bioresour. Technol. 2017, 238, 248–253. [Google Scholar] [CrossRef]
- Solís-Salinas, C.E.; Patlán-Juárez, G.; Okoye, P.U.; Guillén-Garcés, A.; Sebastian, P.J.; Arias, D.M. Long-term semi-continuous production of carbohydrate-enriched microalgae biomass cultivated in low-loaded domestic wastewater. Sci. Total Environ. 2021, 798, 149227. [Google Scholar] [CrossRef]
- Khan, S.A.; Malla, F.A.; Malav, L.C.; Gupta, N.; Kumar, A. Potential of wastewater treating Chlorella minutissima for methane enrichment and CO2 sequestration of biogas and producing lipids. Energy 2018, 150, 153–163. [Google Scholar] [CrossRef]
- Sonkar, S.; Mallick, N. An alternative strategy for enhancing lipid accumulation in chlorophycean microalgae for biodiesel production. J. Appl. Phycol. 2018, 30, 2179–2192. [Google Scholar] [CrossRef]
- Khoo, K.S.; Chew, K.W.; Yew, G.Y.; Leong, W.H.; Chai, Y.H.; Show, P.L.; Chen, W.-H. Recent advances in downstream processing of microalgae lipid recovery for biofuel production. Bioresour. Technol. 2020, 304, 122996. [Google Scholar] [CrossRef]
- Suparmaniam, U.; Lam, M.K.; Uemura, Y.; Lim, J.W.; Lee, K.T.; Shuit, S.H. Insights into the microalgae cultivation technology and harvesting process for biofuel production: A review. Renew. Sust. Energ. Rev. 2019, 115, 109361. [Google Scholar] [CrossRef]
- Mandotra, S.K.; Kumar, P.; Suseela, M.R.; Nayaka, S.; Ramteke, P.W. Evaluation of fatty acid profile and biodiesel properties of microalga Scenedesmus abundans under the influence of phosphorus, pH and light intensities. Bioresour. Technol. 2016, 201, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Dunker, S.; Wilhelm, C. Cell Wall Structure of Coccoid Green Algae as an Important Trade-Off Between Biotic Interference Mechanisms and Multidimensional Cell Growth. Front. Microbiol. 2018, 9, 719. [Google Scholar] [CrossRef] [PubMed]
- Amini Khoeyi, Z.; Seyfabadi, J.; Ramezanpour, Z. Effect of light intensity and photoperiod on biomass and fatty acid composition of the microalgae, Chlorella vulgaris. Aquac. Int. 2012, 20, 41–49. [Google Scholar] [CrossRef]
- Gonçalves, A.L.; Pires, J.C.M.; Simões, M. The effects of light and temperature on microalgal growth and nutrient removal: An experimental and mathematical approach. RSC Adv. 2016, 6, 22896–22907. [Google Scholar] [CrossRef]
- Wang, L.; Addy, M.; Lu, Q.; Cobb, K.; Chen, P.; Chen, X.; Liu, Y.; Wang, H.; Ruan, R. Cultivation of Chlorella vulgaris in sludge extracts: Nutrient removal and algal utilization. Bioresour. Technol. 2019, 280, 505–510. [Google Scholar] [CrossRef]
- Huang, J.; Hankamer, B.; Yarnold, J. Design scenarios of outdoor arrayed cylindrical photobioreactors for microalgae cultivation considering solar radiation and temperature. Algal Res. 2019, 41, 101515. [Google Scholar] [CrossRef]
- Suthar, S.; Verma, R. Production of Chlorella vulgaris under varying nutrient and abiotic conditions: A potential microalga for bioenergy feedstock. Process Saf. Environ. Prot. 2018, 113, 141–148. [Google Scholar] [CrossRef]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Santos, L.O.; Deamici, K.M.; Menestrino, B.C.; Garda-Buffon, J.; Costa, J.A.V. Magnetic treatment of microalgae for enhanced product formation. World J. Microbiol. Biotechnol. 2017, 33, 169. [Google Scholar] [CrossRef]
- Huo, S.; Chen, X.; Zhu, F.; Zhang, W.; Chen, D.; Jin, N.; Cobb, K.; Cheng, Y.; Wang, L.; Ruan, R. Magnetic field intervention on growth of the filamentous microalgae Tribonema sp. in starch wastewater for algal biomass production and nutrients removal: Influence of ambient temperature and operational strategy. Bioresour. Technol. 2020, 303, 122884. [Google Scholar] [CrossRef]
- Menestrino, B.d.C.; Pintos, T.H.C.; Sala, L.; Costa, J.A.V.; Santos, L.O. Application of Static Magnetic Fields on the Mixotrophic Culture of Chlorella minutissima for Carbohydrate Production. Appl. Biochem. Biotechnol. 2020, 192, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Qiu, R.; Gao, S.; Lopez, P.A.; Ogden, K.L. Effects of pH on cell growth, lipid production and CO2 addition of microalgae Chlorella sorokiniana. Algal Res. 2017, 28, 192–199. [Google Scholar] [CrossRef]
- Ren, T. Primary Factors Affecting Growth of Microalgae Optimal Light Exposure Duration and Frequency. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2014. [Google Scholar]
- Maizatul, A.Y.; Radin Mohamed, R.M.S.; Al-Gheethi, A.A.; Hashim, M.K.A. An overview of the utilisation of microalgae biomass derived from nutrient recycling of wet market wastewater and slaughterhouse wastewater. Int. Aquat. Res. 2017, 9, 177–193. [Google Scholar] [CrossRef]
- Daliry, S.; Hallajisani, A.; Mohammadi Roshandeh, J.; Nouri, H.; Golzary, A. Investigation of optimal condition for Chlorella vulgaris microalgae growth. Glob. J. Environ. Sci. 2017, 3, 217–230. [Google Scholar]
- Juneja, A.; Ceballos, R.M.; Murthy, G.S. Effects of Environmental Factors and Nutrient Availability on the Biochemical Composition of Algae for Biofuels Production: A Review. Energies 2013, 6, 4607–4638. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sust. Energ. Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Ho, S.-H.; Kondo, A.; Hasunuma, T.; Chang, J.-S. Engineering strategies for improving the CO2 fixation and carbohydrate productivity of Scenedesmus obliquus CNW-N used for bioethanol fermentation. Bioresour. Technol. 2013, 143, 163–171. [Google Scholar] [CrossRef]
- Rosa, G.M.d.; Moraes, L.; Cardias, B.B.; Souza, M.d.R.A.Z.d.; Costa, J.A.V. Chemical absorption and CO2 biofixation via the cultivation of Spirulina in semicontinuous mode with nutrient recycle. Bioresour. Technol. 2015, 192, 321–327. [Google Scholar] [CrossRef]
- Qu, W.; Loke Show, P.; Hasunuma, T.; Ho, S.-H. Optimizing real swine wastewater treatment efficiency and carbohydrate productivity of newly microalga Chlamydomonas sp. QWY37 used for cell-displayed bioethanol production. Bioresour. Technol. 2020, 305, 123072. [Google Scholar] [CrossRef]
- Zhou, W.; Lu, Q.; Han, P.; Li, J. Chapter 3—Microalgae Cultivation and Photobioreactor Design. In Microalgae Cultivation for Biofuels Production; Yousuf, A., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 31–50. [Google Scholar] [CrossRef]
- Li, X.; Li, W.; Zhai, J.; Wei, H. Effect of nitrogen limitation on biochemical composition and photosynthetic performance for fed-batch mixotrophic cultivation of microalga Spirulina platensis. Bioresour. Technol. 2018, 263, 555–561. [Google Scholar] [CrossRef]
- Verma, R.; Kumari, K.V.L.K.; Srivastava, A.; Kumar, A. Photoautotrophic, mixotrophic, and heterotrophic culture media optimization for enhanced microalgae production. J. Environ. Chem. Eng. 2020, 8, 104149. [Google Scholar] [CrossRef]
- Li, T.; Cheng, J.; Zhang, X.; Liu, J.; Huang, R.; Zhou, J. Jet range hydrocarbons converted from microalgal biodiesel over mesoporous zeolite-based catalysts. Int. J. Hydrog. Energy 2018, 43, 9988–9993. [Google Scholar] [CrossRef]
- Gonçalves, C.F.; Menegol, T.; Rech, R. Biochemical composition of green microalgae Pseudoneochloris marina grown under different temperature and light conditions. Biocatal. Agric. Biotechnol. 2019, 18, 101032. [Google Scholar] [CrossRef]
- Lee, S.Y.; Khoiroh, I.; Vo, D.-V.N.; Senthil Kumar, P.; Show, P.L. Techniques of lipid extraction from microalgae for biofuel production: A review. Environ. Chem. Lett. 2021, 19, 231–251. [Google Scholar] [CrossRef]
- Yap, J.K.; Sankaran, R.; Chew, K.W.; Halimatul Munawaroh, H.S.; Ho, S.-H.; Rajesh Banu, J.; Show, P.L. Advancement of green technologies: A comprehensive review on the potential application of microalgae biomass. Chemosphere 2021, 281, 130886. [Google Scholar] [CrossRef]
- Mishra, S.; Maiti, A. The efficiency of Eichhornia crassipes in the removal of organic and inorganic pollutants from wastewater: A review. Environ. Sci. Pollut. Res. 2017, 24, 7921–7937. [Google Scholar] [CrossRef]
- Ranjith Kumar, R.; Hanumantha Rao, P.; Arumugam, M. Lipid Extraction Methods from Microalgae: A Comprehensive Review. Front. Energy Res. 2015, 2, 61. [Google Scholar] [CrossRef]
- Patil, P.D.; Reddy, H.; Muppaneni, T.; Schaub, T.; Holguin, F.O.; Cooke, P.; Lammers, P.; Nirmalakhandan, N.; Li, Y.; Lu, X.; et al. In situ ethyl ester production from wet algal biomass under microwave-mediated supercritical ethanol conditions. Bioresour. Technol. 2013, 139, 308–315. [Google Scholar] [CrossRef]
- Amaro, H.M.; Fernandes, F.; Valentão, P.; Andrade, P.B.; Sousa-Pinto, I.; Malcata, F.X.; Guedes, A.C. Effect of Solvent System on Extractability of Lipidic Components of Scenedesmus obliquus (M2-1) and Gloeothece sp. on Antioxidant Scavenging Capacity Thereof. Marine Drugs 2015, 13, 6453–6471. [Google Scholar] [CrossRef]
- Qiu, C.; He, Y.; Huang, Z.; Li, S.; Huang, J.; Wang, M.; Chen, B. Lipid extraction from wet Nannochloropsis biomass via enzyme-assisted three phase partitioning. Bioresour. Technol. 2019, 284, 381–390. [Google Scholar] [CrossRef]
- Karemore, A.; Sen, R. Downstream processing of microalgal feedstock for lipid and carbohydrate in a biorefinery concept: A holistic approach for biofuel applications. RSC Adv. 2016, 6, 29486–29496. [Google Scholar] [CrossRef]
- Jeevan Kumar, S.P.; Vijay Kumar, G.; Dash, A.; Scholz, P.; Banerjee, R. Sustainable green solvents and techniques for lipid extraction from microalgae: A review. Algal Res. 2017, 21, 138–147. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Du, T.-B.; Pi, H.-C.; Jang, S.-M.; Lin, Y.-H.; Lee, H.-T. Comparative study of lipid extraction from microalgae by organic solvent and supercritical CO2. Bioresour. Technol. 2011, 102, 10151–10153. [Google Scholar] [CrossRef] [PubMed]
- Sati, H.; Mitra, M.; Mishra, S.; Baredar, P. Microalgal lipid extraction strategies for biodiesel production: A review. Algal Res. 2019, 38, 101413. [Google Scholar] [CrossRef]
- Deshmukh, S.; Kumar, R.; Bala, K. Microalgae biodiesel: A review on oil extraction, fatty acid composition, properties and effect on engine performance and emissions. Fuel Process. Technol. 2019, 191, 232–247. [Google Scholar] [CrossRef]
- Lorenzen, J.; Igl, N.; Tippelt, M.; Stege, A.; Qoura, F.; Sohling, U.; Brück, T. Extraction of microalgae derived lipids with supercritical carbon dioxide in an industrial relevant pilot plant. Bioprocess Biosyst Eng. 2017, 40, 911–918. [Google Scholar] [CrossRef]
- McKennedy, J.; Önenç, S.; Pala, M.; Maguire, J. Supercritical carbon dioxide treatment of the microalgae Nannochloropsis oculata for the production of fatty acid methyl esters. J. Supercrit. Fluids 2016, 116, 264–270. [Google Scholar] [CrossRef]
- Obeid, S.; Beaufils, N.; Camy, S.; Takache, H.; Ismail, A.; Pontalier, P.-Y. Supercritical carbon dioxide extraction and fractionation of lipids from freeze-dried microalgae Nannochloropsis oculata and Chlorella vulgaris. Algal Res. 2018, 34, 49–56. [Google Scholar] [CrossRef]
- Patel, A.; Matsakas, L.; Sartaj, K.; Chandra, R. Chapter 2—Extraction of lipids from algae using supercritical carbon dioxide. In Green Sustainable Process for Chemical and Environmental Engineering and Science; Inamuddin Asiri, A.M., Isloor, A.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 17–39. [Google Scholar] [CrossRef]
- Patil, P.D.; Dandamudi, K.P.R.; Wang, J.; Deng, Q.; Deng, S. Extraction of bio-oils from algae with supercritical carbon dioxide and co-solvents. J. Supercrit. Fluids 2018, 135, 60–68. [Google Scholar] [CrossRef]
- Cicci, A.; Sed, G.; Bravi, M. Potential of choline chloride–based natural deep eutectic solvents (NaDES) in the extraction of microalgal metabolites. Chem. Eng. Trans. 2017, 57, 61–66. [Google Scholar]
- Lu, W.; Alam, M.A.; Pan, Y.; Wu, J.; Wang, Z.; Yuan, Z. A new approach of microalgal biomass pretreatment using deep eutectic solvents for enhanced lipid recovery for biodiesel production. Bioresour. Technol. 2016, 218, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Alam, M.A.; Wang, Z.; Huang, D.; Hu, K.; Chen, H.; Yuan, Z. One-step production of biodiesel from wet and unbroken microalgae biomass using deep eutectic solvent. Bioresour. Technol. 2017, 238, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Sed, G.; Cicci, A.; Jessop, P.G.; Bravi, M. A novel switchable-hydrophilicity, natural deep eutectic solvent (NaDES)-based system for bio-safe biorefinery. RSC Adv. 2018, 8, 37092–37097. [Google Scholar] [CrossRef]
- Tommasi, E.; Cravotto, G.; Galletti, P.; Grillo, G.; Mazzotti, M.; Sacchetti, G.; Samorì, C.; Tabasso, S.; Tacchini, M.; Tagliavini, E. Enhanced and Selective Lipid Extraction from the Microalga P. tricornutum by Dimethyl Carbonate and Supercritical CO2 Using Deep Eutectic Solvents and Microwaves as Pretreatment. ACS Sustain. Chem. Eng. 2017, 5, 8316–8322. [Google Scholar] [CrossRef]
- Słupek, E.; Makoś, P.; Gębicki, J. Theoretical and Economic Evaluation of Low-Cost Deep Eutectic Solvents for Effective Biogas Upgrading to Bio-Methane. Energies 2020, 13, 3379. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Nagle, N.; Lemke, P. Production of methyl ester fuel from microalgae. Appl. Biochem. Biotechnol. 1990, 24, 355–361. [Google Scholar] [CrossRef]
- Richter, B.E.; Jones, B.A.; Ezzell, J.L.; Porter, N.L.; Avdalovic, N.; Pohl, C. Accelerated Solvent Extraction: A Technique for Sample Preparation. Anal. Chem. 1996, 68, 1033–1039. [Google Scholar] [CrossRef]
- Bu, X.; Pang, M.; Wang, B.; Zhang, Y.; Xie, K.; Zhao, X.; Wang, Y.; Guo, Y.; Liu, C.; Wang, R.; et al. Determination of Piperazine in Eggs Using Accelerated Solvent Extraction (ASE) and Solid Phase Extraction (SPE) with High-Performance Liquid Chromatography—Fluorescence Detection (HPLC-FLD) and Pre-Column Derivatization with Dansyl Chloride. Anal. Lett. 2020, 53, 53–71. [Google Scholar] [CrossRef]
- Mulbry, W.; Kondrad, S.; Buyer, J.; Luthria, D.L. Optimization of an Oil Extraction Process for Algae from the Treatment of Manure Effluent. J. Am. Oil Chem. Soc. 2009, 86, 909–915. [Google Scholar] [CrossRef]
- Choi, K.J.; Nakhost, Z.; Krukonis, V.J.; Karel, M. Supercritical fluid extraction and characterization of lipids from algae scenedesmus obliquus. Food Biotechnol. 1987, 1, 263–281. [Google Scholar] [CrossRef]
- Li, Y.; Ghasemi Naghdi, F.; Garg, S.; Adarme-Vega, T.C.; Thurecht, K.J.; Ghafor, W.A.; Tannock, S.; Schenk, P.M. A comparative study: The impact of different lipid extraction methods on current microalgal lipid research. Microb. Cell Factories 2014, 13, 14. [Google Scholar] [CrossRef]
- Laurens, L.M.L.; Quinn, M.; Van Wychen, S.; Templeton, D.W.; Wolfrum, E.J. Accurate and reliable quantification of total microalgal fuel potential as fatty acid methyl esters by in situ transesterification. Anal. Bioanal. Chem. 2012, 403, 167–178. [Google Scholar] [CrossRef]
- Ríos, S.D.; Castañeda, J.; Torras, C.; Farriol, X.; Salvadó, J. Lipid extraction methods from microalgal biomass harvested by two different paths: Screening studies toward biodiesel production. Bioresour. Technol. 2013, 133, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Patnaik, R.; Singh, A.K.; Mallick, N. Comparative assessment of various lipid extraction protocols and optimization of transesterification process for microalgal biodiesel production. Environ. Technol. 2013, 34, 2009–2018. [Google Scholar] [CrossRef]
- Hajinajaf, N.; Rabbani, Y.; Mehrabadi, A.; Tavakoli, O. Experimental and modeling assessment of large-scale cultivation of microalgae Nannochloropsis sp. PTCC 6016 to reach high efficiency lipid extraction. Int. J. Environ. Sci. Technol. 2022, 19, 5511–5528. [Google Scholar] [CrossRef]
- Saini, R.K.; Prasad, P.; Shang, X.; Keum, Y.-S. Advances in Lipid Extraction Methods—A Review. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.C. Thermochemical Processing of Biomass: Conversion into Fuels, Chemicals and Power; John Wiley & Sons: New York, NY, USA, 2019. [Google Scholar]
- Boumanchar, I.; Chhiti, Y.; M’Hamdi Alaoui, F.E.; Elkhouakhi, M.; Sahibed-dine, A.; Bentiss, F.; Jama, C.; Bensitel, M. Investigation of (co)-combustion kinetics of biomass, coal and municipal solid wastes. Waste Manag. 2019, 97, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Bhuiya, M.M.K.; Rasul, M.; Khan, M.; Ashwath, N.; Mofijur, M. Comparison of oil extraction between screw press and solvent (n-hexane) extraction technique from beauty leaf (Calophyllum inophyllum L.) feedstock. Ind. Crops Prod. 2020, 144, 112024. [Google Scholar] [CrossRef]
- Sajid, Z.; Khan, F.; Zhang, Y. Process simulation and life cycle analysis of biodiesel production. Renew. Energy 2016, 85, 945–952. [Google Scholar] [CrossRef]
- Hidalgo, P.; Toro, C.; Ciudad, G.; Navia, R. Advances in direct transesterification of microalgal biomass for biodiesel production. Rev. Environ. Sci. Biotechnol. 2013, 12, 179–199. [Google Scholar] [CrossRef]
- Siddiki, S.Y.A.; Mofijur, M.; Kumar, P.S.; Ahmed, S.F.; Inayat, A.; Kusumo, F.; Badruddin, I.A.; Khan, T.M.Y.; Nghiem, L.D.; Ong, H.C.; et al. Microalgae biomass as a sustainable source for biofuel, biochemical and biobased value-added products: An integrated biorefinery concept. Fuel 2022, 307, 121782. [Google Scholar] [CrossRef]
- Teo, S.H.; Islam, A.; Ng, F.L.; Taufiq-Yap, Y.H. Biodiesel synthesis from photoautotrophic cultivated oleoginous microalgae using a sand dollar catalyst. RSC Adv. 2015, 5, 47140–47152. [Google Scholar] [CrossRef]
- Umdu, E.S.; Tuncer, M.; Seker, E. Transesterification of Nannochloropsis oculata microalga’s lipid to biodiesel on Al
2O
3 supported CaO and MgO catalysts. Bioresour. Technol. 2009, 100, 2828–2831. [Google Scholar] [CrossRef] [PubMed] - Catone, C.M.; Ripa, M.; Geremia, E.; Ulgiati, S. Bio-products from algae-based biorefinery on wastewater: A review. J. Environ. Manag. 2021, 293, 112792. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, C.; Song, L.; Sommerfeld, M.; Hu, Q. A high throughput Nile red method for quantitative measurement of neutral lipids in microalgae. J. Microbiol. Methods 2009, 77, 41–47. [Google Scholar] [CrossRef]
- Ungureanu, N.; Vladut, V.; Biris, S.-S. Capitalization of wastewater-grown algae in bioethanol production. In Proceedings of the 19th International Scientific Conference, Jelgava, Latvia, 20–22 May 2020; Engineering for Rural Development, Latvia University of Life Sciences and Technologies: Jelgava, Latvia, 2020; pp. 1859–1864. [Google Scholar]
- Kim, S.W.; Koo, B.S.; Lee, D.H. A comparative study of bio-oils from pyrolysis of microalgae and oil seed waste in a fluidized bed. Bioresour. Technol. 2014, 162, 96–102. [Google Scholar] [CrossRef]
- Velazquez-Lucio, J.; Rodríguez-Jasso, R.M.; Colla, L.M.; Sáenz-Galindo, A.; Cervantes-Cisneros, D.E.; Aguilar, C.N.; Fernandes, B.D.; Ruiz, H.A. Microalgal biomass pretreatment for bioethanol production: A review. Biofuel Res. J. 2018, 5, 780–791. [Google Scholar] [CrossRef]
- Ramos Tercero, E.A.; Domenicali, G.; Bertucco, A. Autotrophic production of biodiesel from microalgae: An updated process and economic analysis. Energy 2014, 76, 807–815. [Google Scholar] [CrossRef]
- Patel, S.K.S.; Kumar, P.; Mehariya, S.; Purohit, H.J.; Lee, J.-K.; Kalia, V.C. Enhancement in hydrogen production by co-cultures of Bacillus and Enterobacter. Int. J. Hydrog. Energy 2014, 39, 14663–14668. [Google Scholar] [CrossRef]
- Rashid, N.; Rehman, M.S.U.; Memon, S.; Ur Rahman, Z.; Lee, K.; Han, J.-I. Current status, barriers and developments in biohydrogen production by microalgae. Renew. Sust. Energ. Rev. 2013, 22, 571–579. [Google Scholar] [CrossRef]
- Batista, A.P.; Ambrosano, L.; Graça, S.; Sousa, C.; Marques, P.A.S.S.; Ribeiro, B.; Botrel, E.P.; Castro Neto, P.; Gouveia, L. Combining urban wastewater treatment with biohydrogen production—An integrated microalgae-based approach. Bioresour. Technol. 2015, 184, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Fakhimi, N.; Tavakoli, O. Improving hydrogen production using co-cultivation of bacteria with Chlamydomonas reinhardtii microalga. Mater. Sci. Energy Technol. 2019, 2, 1–7. [Google Scholar] [CrossRef]
- Vargas, S.R.; Santos, P.V.d.; Zaiat, M.; Calijuri, M.d.C. Optimization of biomass and hydrogen production by Anabaena sp. (UTEX 1448) in nitrogen-deprived cultures. Biomass Bioenergy 2018, 111, 70–76. [Google Scholar] [CrossRef]
- Show, K.-Y.; Lee, D.-J.; Chang, J.-S. Bioreactor and process design for biohydrogen production. Bioresour. Technol. 2011, 102, 8524–8533. [Google Scholar] [CrossRef]
- Biller, P.; Ross, A.B. Potential yields and properties of oil from the hydrothermal liquefaction of microalgae with different biochemical content. Bioresour. Technol. 2011, 102, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Harman-Ware, A.E.; Morgan, T.; Wilson, M.; Crocker, M.; Zhang, J.; Liu, K.; Stork, J.; Debolt, S. Microalgae as a renewable fuel source: Fast pyrolysis of Scenedesmus sp. Renew. Energy 2013, 60, 625–632. [Google Scholar] [CrossRef]
- Yang, W.; Wang, H.; Zhang, M.; Zhu, J.; Zhou, J.; Wu, S. Fuel properties and combustion kinetics of hydrochar prepared by hydrothermal carbonization of bamboo. Bioresour. Technol. 2016, 205, 199–204. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, Z.; Zhang, X.; Liu, J.; Zhou, J.; Cen, K. Sulfonated mesoporous Y zeolite with nickel to catalyze hydrocracking of microalgae biodiesel into jet fuel range hydrocarbons. Int. J. Hydrog. Energy 2019, 44, 1650–1658. [Google Scholar] [CrossRef]
- Bwapwa, J.K.; Anandraj, A.; Trois, C. Microalgae processing for jet fuel production. Biofuels Bioprod. Biorefin. 2018, 12, 522–535. [Google Scholar] [CrossRef]
- Li, L.; Coppola, E.; Rine, J.; Miller, J.L.; Walker, D. Catalytic Hydrothermal Conversion of Triglycerides to Non-ester Biofuels. Energy Fuels 2010, 24, 1305–1315. [Google Scholar] [CrossRef]
- McAfee, E.A. The Aemetis Biorefinery: 100% Replacement, Renewable Jet and Diesel Fuels by Conversion of Existing Biofuels Refinery Facilities. In Proceedings of the Advanced Biofuels Marketing Conference, Amsterdam, The Netherlands, 28–29 June 2012. [Google Scholar]
- Corporan, E.; Edwards, T.; Shafer, L.; DeWitt, M.J.; Klingshirn, C.; Zabarnick, S.; West, Z.; Striebich, R.; Graham, J.; Klein, J. Chemical, Thermal Stability, Seal Swell, and Emissions Studies of Alternative Jet Fuels. Energy Fuels 2011, 25, 955–966. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, Q.; Guan, Q.; He, L.; Li, W. Bio-aviation fuel production from hydroprocessing castor oil promoted by the nickel-based bifunctional catalysts. Bioresour. Technol. 2015, 183, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Scheuermann, S.S.; Forster, S.; Eibl, S. In-Depth Interpretation of Mid-Infrared Spectra of Various Synthetic Fuels for the Chemometric Prediction of Aviation Fuel Blend Properties. Energy Fuels 2017, 31, 2934–2943. [Google Scholar] [CrossRef]
- Buffi, M.; Valera-Medina, A.; Marsh, R.; Pugh, D.; Giles, A.; Runyon, J.; Chiaramonti, D. Emissions characterization tests for hydrotreated renewable jet fuel from used cooking oil and its blends. Appl. Energy 2017, 201, 84–93. [Google Scholar] [CrossRef]
- Gutiérrez-Antonio, C.; Gómez-Castro, F.I.; de Lira-Flores, J.A.; Hernández, S. A review on the production processes of renewable jet fuel. Renew. Sust. Energ. Rev. 2017, 79, 709–729. [Google Scholar] [CrossRef]
- Gómez-De la Cruz, A.; Romero-Izquierdo, A.G.; Gutiérrez-Antonio, C.; Gómez-Castro, F.I.; Hernández, S. Modelling of the hydrotreating process to produce renewable aviation fuel from micro-algae oil. In Computer Aided Chemical Engineering; Espuña, A., Graells, M., Puigjaner, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 40, pp. 655–660. [Google Scholar]
- Garcia Alba, L.; Torri, C.; Samorì, C.; van der Spek, J.; Fabbri, D.; Kersten, S.R.A.; Brilman, D.W.F. Hydrothermal Treatment (HTT) of Microalgae: Evaluation of the Process As Conversion Method in an Algae Biorefinery Concept. Energy Fuels 2012, 26, 642–657. [Google Scholar] [CrossRef]
- Samiee-Zafarghandi, R.; Karimi-Sabet, J.; Abdoli, M.A.; Karbassi, A. Supercritical water gasification of microalga Chlorella PTCC 6010 for hydrogen production: Box-Behnken optimization and evaluating catalytic effect of MnO2/SiO2 and NiO/SiO2. Renew. Energy 2018, 126, 189–201. [Google Scholar] [CrossRef]
- Kim, T.-H.; Lee, K.; Oh, B.-R.; Lee, M.-E.; Seo, M.; Li, S.; Kim, J.-K.; Choi, M.; Chang, Y.K. A novel process for the coproduction of biojet fuel and high-value polyunsaturated fatty acid esters from heterotrophic microalgae Schizochytrium sp. ABC101. Renew. Energy 2021, 165, 481–490. [Google Scholar] [CrossRef]
- Ismail, M.M.; Ismail, G.A.; El-Sheekh, M.M. Potential assessment of some micro- and macroalgal species for bioethanol and biodiesel production. Energy Sources Part A: Recovery Util. Environ. Eff. 2020, 1–17. [Google Scholar] [CrossRef]
- Reyimu, Z.; Özçimen, D. Batch cultivation of marine microalgae Nannochloropsis oculata and Tetraselmis suecica in treated municipal wastewater toward bioethanol production. J. Clean. Prod. 2017, 150, 40–46. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, O.K.; Kim, C.H.; Seo, J.-W.; Oh, B.-R.; Lee, E.Y. Lipase-catalyzed in-situ biosynthesis of glycerol-free biodiesel from heterotrophic microalgae, Aurantiochytrium sp. KRS101 biomass. Bioresour. Technol. 2016, 211, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, Y.; Viamajala, S.; Varanasi, S. In situ and Ex situ Catalytic Pyrolysis of Microalgae and Integration With Pyrolytic Fractionation. Front. Chem. 2020, 8, 786. [Google Scholar] [CrossRef]
- Hidalgo, P.; Ciudad, G.; Schober, S.; Mittelbach, M.; Navia, R. Biodiesel synthesis by direct transesterification of microalga Botryococcus braunii with continuous methanol reflux. Bioresour. Technol. 2015, 181, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Im, H.; Kim, B.; Lee, J.W. Concurrent production of biodiesel and chemicals through wet in situ transesterification of microalgae. Bioresour. Technol. 2015, 193, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.B.; Pinnarat, T.; Savage, P.E. Biodiesel Production from Wet Algal Biomass through in Situ Lipid Hydrolysis and Supercritical Transesterification. Energy Fuels 2010, 24, 5235–5243. [Google Scholar] [CrossRef]
- Jadhao, P.R.; Vuppaladadiyam, A.K.; Prakash, A.; Pant, K.K. Co-pyrolysis characteristics and kinetics of electronic waste and macroalgae: A synergy study based on thermogravimetric analysis. Algal Res. 2022, 61, 102601. [Google Scholar] [CrossRef]
- Rasam, S.; Azizi, K.; Moraveji, M.K.; Akbari, A.; Soria-Verdugo, A. Insights into the co–pyrolysis of olive stone, waste polyvinyl chloride and Spirulina microalgae blends through thermogravimetric analysis. Algal Res. 2022, 62, 102635. [Google Scholar] [CrossRef]
- Shuping, Z.; Yulong, W.; Mingde, Y.; Chun, L.; Junmao, T. Pyrolysis characteristics and kinetics of the marine microalgae Dunaliella tertiolecta using thermogravimetric analyzer. Bioresour. Technol. 2010, 101, 359–365. [Google Scholar] [CrossRef]
- Rizzo, A.M.; Prussi, M.; Bettucci, L.; Libelli, I.M.; Chiaramonti, D. Characterization of microalga Chlorella as a fuel and its thermogravimetric behavior. Appl. Energy 2013, 102, 24–31. [Google Scholar] [CrossRef]
- Agrawal, A.; Chakraborty, S. A kinetic study of pyrolysis and combustion of microalgae Chlorella vulgaris using thermo-gravimetric analysis. Bioresour. Technol. 2013, 128, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Wu, Q.; Tu, P. Pyrolytic characteristics of heterotrophic Chlorella protothecoides for renewable bio-fuel production. J. Appl. Phycol. 2001, 13, 5–12. [Google Scholar] [CrossRef]
- Kim, K.H.; Choi, I.S.; Kim, H.M.; Wi, S.G.; Bae, H.-J. Bioethanol production from the nutrient stress-induced microalga Chlorella vulgaris by enzymatic hydrolysis and immobilized yeast fermentation. Bioresour. Technol. 2014, 153, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Silva, L.; López-González, D.; Garcia-Minguillan, A.M.; Valverde, J.L. Pyrolysis, combustion and gasification characteristics of Nannochloropsis gaditana microalgae. Bioresour. Technol. 2013, 130, 321–331. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, X.; Yang, X. Co-pyrolysis characteristics of microalgae Isochrysis and Chlorella: Kinetics, biocrude yield and interaction. Bioresour. Technol. 2015, 198, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Fayyaz, M.; Chew, K.W.; Show, P.L.; Ling, T.C.; Ng, I.S.; Chang, J.-S. Genetic engineering of microalgae for enhanced biorefinery capabilities. Biotechnol. Adv. 2020, 43, 107554. [Google Scholar] [CrossRef]
- Jiang, W.; Brueggeman Andrew, J.; Horken Kempton, M.; Plucinak Thomas, M.; Weeks Donald, P. Successful Transient Expression of Cas9 and Single Guide RNA Genes in Chlamydomonas reinhardtii. Eukaryot. Cell 2014, 13, 1465–1469. [Google Scholar] [CrossRef]
- Baek, K.; Kim, D.H.; Jeong, J.; Sim, S.J.; Melis, A.; Kim, J.-S.; Jin, E.; Bae, S. DNA-free two-gene knockout in Chlamydomonas reinhardtii via CRISPR-Cas9 ribonucleoproteins. Sci. Rep. 2016, 6, 30620. [Google Scholar] [CrossRef]
- Ajjawi, I.; Verruto, J.; Aqui, M.; Soriaga, L.B.; Coppersmith, J.; Kwok, K.; Peach, L.; Orchard, E.; Kalb, R.; Xu, W.; et al. Lipid production in Nannochloropsis gaditana is doubled by decreasing expression of a single transcriptional regulator. Nat. Biotechnol. 2017, 35, 647–652. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; DeBono, A.; Durrett, T.P. Acyl-lipid metabolism. Arab. Book/Am. Soc. Plant Biol. 2013, 11, e0161. [Google Scholar] [CrossRef]
- Deng, X.-D.; Gu, B.; Li, Y.-J.; Hu, X.-W.; Guo, J.-C.; Fei, X.-W. The roles of acyl-CoA: Diacylglycerol acyltransferase 2 genes in the biosynthesis of triacylglycerols by the green algae Chlamydomonas reinhardtii. Mol. Plant 2012, 5, 945–947. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.-F.; Zhang, M.-H.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Bai, W.-B.; Li, H.-Y. Improvement of Neutral Lipid and Polyunsaturated Fatty Acid Biosynthesis by Overexpressing a Type 2 Diacylglycerol Acyltransferase in Marine Diatom Phaeodactylum tricornutum. Marine Drugs 2013, 11, 4558–4569. [Google Scholar] [CrossRef] [PubMed]
- Cases, S.; Smith, S.J.; Zheng, Y.-W.; Myers, H.M.; Lear, S.R.; Sande, E.; Novak, S.; Collins, C.; Welch, C.B.; Lusis, A.J.; et al. Identification of a gene encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 13018–13023. [Google Scholar] [CrossRef] [PubMed]
- Ndimba, B.K.; Ndimba, R.J.; Johnson, T.S.; Waditee-Sirisattha, R.; Baba, M.; Sirisattha, S.; Shiraiwa, Y.; Agrawal, G.K.; Rakwal, R. Biofuels as a sustainable energy source: An update of the applications of proteomics in bioenergy crops and algae. J. Proteom. 2013, 93, 234–244. [Google Scholar] [CrossRef]
- Stauber, E.J.; Hippler, M. Chlamydomonas reinhardtii proteomics. Plant Physiol. Biochem. 2004, 42, 989–1001. [Google Scholar] [CrossRef]
- Pandhal, J.; Wright, P.C.; Biggs, C.A. Proteomics with a pinch of salt: A cyanobacterial perspective. Saline Syst. 2008, 4, 1. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Baudet, M.; Cuiné, S.; Adriano, J.-M.; Barthe, D.; Billon, E.; Bruley, C.; Beisson, F.; Peltier, G.; Ferro, M.; et al. Proteomic profiling of oil bodies isolated from the unicellular green microalga Chlamydomonas reinhardtii: With focus on proteins involved in lipid metabolism. Proteomics 2011, 11, 4266–4273. [Google Scholar] [CrossRef]
- Cao, J.; Li, J.-L.; Li, D.; Tobin, J.F.; Gimeno, R.E. Molecular identification of microsomal acyl-CoA:glycerol-3-phosphate acyltransferase, a key enzyme in de novo triacylglycerol synthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19695–19700. [Google Scholar] [CrossRef]
- Trentacoste, E.M.; Shrestha, R.P.; Smith, S.R.; Glé, C.; Hartmann, A.C.; Hildebrand, M.; Gerwick, W.H. Metabolic engineering of lipid catabolism increases microalgal lipid accumulation without compromising growth. Proc. Natl. Acad. Sci. USA 2013, 110, 19748–19753. [Google Scholar] [CrossRef]
- Jamers, A.; Blust, R.; De Coen, W. Omics in algae: Paving the way for a systems biological understanding of algal stress phenomena? Aquat. Toxicol. 2009, 92, 114–121. [Google Scholar] [CrossRef]
- Banerjee, C.; Dubey, K.K.; Shukla, P. Metabolic Engineering of Microalgal Based Biofuel Production: Prospects and Challenges. Front. Microbiol. 2016, 7, 432. [Google Scholar] [CrossRef] [PubMed]
- Abreu, A.C.; Molina-Miras, A.; Aguilera-Sáez, L.M.; López-Rosales, L.; Cerón-García, M.d.C.; Sánchez-Mirón, A.; Olmo-García, L.; Carrasco-Pancorbo, A.; García-Camacho, F.; Molina-Grima, E.; et al. Production of Amphidinols and Other Bioproducts of Interest by the Marine Microalga Amphidinium carterae Unraveled by Nuclear Magnetic Resonance Metabolomics Approach Coupled to Multivariate Data Analysis. J. Agric. Food Chem. 2019, 67, 9667–9682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, Z.; Ren, Y.; Wang, L.; Zhang, J.; Liang, C.; Tong, S.; Wang, Y.; Xu, D.; Zhang, X.; et al. Integrating Transcriptomics and Metabolomics to Characterize Metabolic Regulation to Elevated CO2 in Chlamydomonas Reinhardtii. Mar. Biotechnol. 2021, 23, 255–275. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Xu, L.; Huang, R.; Wang, Q. Improved biohydrogen production with an expression of codon-optimized hemH and lba genes in the chloroplast of Chlamydomonas reinhardtii. Bioresour. Technol. 2011, 102, 2610–2616. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-W.; Liu, W.-J.; Hu, D.-X.; Wang, X.; Balamurugan, S.; Alimujiang, A.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Identification of a malonyl CoA-acyl carrier protein transacylase and its regulatory role in fatty acid biosynthesis in oleaginous microalga Nannochloropsis oceanica. Biotechnol. Appl. Biochem. 2017, 64, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Yuan, C.; Jin, Y.; Hu, G.-R.; Li, F.-L. Characterization of 3-ketoacyl-coA synthase in a nervonic acid producing oleaginous microalgae Mychonastes afer. Algal Res. 2018, 31, 225–231. [Google Scholar] [CrossRef]
- Deng, X.; Cai, J.; Fei, X. Effect of the expression and knockdown of citrate synthase gene on carbon flux during triacylglycerol biosynthesis by green algae Chlamydomonas reinhardtii. BMC Biochem. 2013, 14, 38. [Google Scholar] [CrossRef]
- Shang, C.; Zhu, S.; Wang, Z.; Qin, L.; Alam, M.A.; Xie, J.; Yuan, Z. Proteome response of Dunaliella parva induced by nitrogen limitation. Algal Res. 2017, 23, 196–202. [Google Scholar] [CrossRef]
- Arora, N.; Kumari, P.; Kumar, A.; Gangwar, R.; Gulati, K.; Pruthi, P.A.; Prasad, R.; Kumar, D.; Pruthi, V.; Poluri, K.M. Delineating the molecular responses of a halotolerant microalga using integrated omics approach to identify genetic engineering targets for enhanced TAG production. Biotechnol. Biofuels 2019, 12, 2. [Google Scholar] [CrossRef]
- Balamurugan, S.; Wang, X.; Wang, H.-L.; An, C.-J.; Li, H.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Occurrence of plastidial triacylglycerol synthesis and the potential regulatory role of AGPAT in the model diatom Phaeodactylum tricornutum. Biotechnol. Biofuels 2017, 10, 97. [Google Scholar] [CrossRef]
- Babu, S.S.; Gondi, R.; Vincent, G.S.; JohnSamuel, G.C.; Jeyakumar, R.B. Microalgae Biomass and Lipids as Feedstock for Biofuels: Sustainable Biotechnology Strategies. Sustainability 2022, 14, 15070. [Google Scholar] [CrossRef]
- Ubando, A.T.; Anderson, S.; Ng, E.; Chen, W.-H.; Culaba, A.B.; Kwon, E.E. Life cycle assessment of microalgal biorefinery: A state-of-the-art review. Bioresour. Technol. 2022, 360, 127615. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, J.; Huang, Z.; Liu, T.; Li, H. Photothermal technique-enabled ambient production of microalgae biodiesel: Mechanism and life cycle assessment. Bioresour. Technol. 2023, 369, 128390. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Priya, A.K.; Dutta, K.; Rajendran, S.; Vasseghian, Y. Engineering strategies and opportunities of next generation biofuel from microalgae: A perspective review on the potential bioenergy feedstock. Fuel 2022, 312, 122827. [Google Scholar] [CrossRef]
- Huang, R.; Li, J.; Tang, Y.; Song, W.; Yu, Y.; Yang, W.; Cheng, J. Comparative life-cycle assessment of microalgal biodiesel production via various emerging wet scenarios: Energy conversion characteristics and environmental impacts. Energy Convers. Manag. 2022, 257, 115427. [Google Scholar] [CrossRef]
- Goswami, R.K.; Agrawal, K.; Upadhyaya, H.M.; Gupta, V.K.; Verma, P. Microalgae conversion to alternative energy, operating environment and economic footprint: An influential approach towards energy conversion, and management. Energy Convers. Manag. 2022, 269, 116118. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Solvent | Example | Lipid Extraction Efficacy | Efficiency Rating | Cost Involved | Energy Needs | References |
|---|---|---|---|---|---|---|
| Organic solvents | Methanol, chloroform, hexane, ether | Depends on microalgae strain, Volume of extractor Reaction time, Lipid types present in microalgae cell, Solvent ratios | Moderate | High cost due to the utilization of organic solvents. Recycling and reusing the organic solvents after absorption can save on costs and reduce waste. | Energy intensive, if involving the cell-wall disruption method, for example microwave and ultrasound pre-treatment | Ranjith Kumar, Hanumantha Rao and Arumugam [73] |
| Supercritical fluid | Supercritical CO2 | High | High cost due to energy consumption, maintenance fees, infrastructure requirements, and operation costs | Energy intensive due to the use of high pressure | Lorenzen, et al. [82], McKennedy, et al. [83], Obeid, et al. [84], Patel, et al. [85], Patil, et al. [86] | |
| Accelerated solvent extraction (ASE) | Methanol, chloroform, hexane, ether | High | High relative capital cost as compared to Soxhlet extraction | Energy intensive if involving the cell-wall disruption method, for example microwave and ultrasound pre-treatment | Yadav, Krishnan, Gupta, Prasad, Amin, Cabral-Pinto, Sharma, Marzouki, Jeon, Kumar, Singh, Kumar, Rezania and Islam [5] | |
| Deep eutectic solvents (DES) | Choline chloride-oxalic acid, Choline-chloride-acetic acid | High | Low cost for organic components. The unit cost of DESs is primarily determined by the DES structure, which determines absorption capacity and regeneration cycles. Recycling and reusing DES after absorption reduces annual operating costs and waste | Energy intensive if the pre-treatment steps are involved, for example microwave and ultrasound treatment | Cicci, et al. [87], Lu, et al. [88], Pan, et al. [89], Sed, et al. [90], Tommasi, et al. [91], Słupek, et al. [92] |
| Technology | Operational Conditions | |||
|---|---|---|---|---|
| Temperature (°C) | Heating Rate (°Cs−1) | Time Duration | Other Parameters | |
| Combustion | 700–800 | - | - | Air flow |
| Gasification | 700–1000 | - | - | Controlled O2 |
| Pyrolysis -Slow pyrolysis -Fast pyrolysis -Flash pyrolysis | 300–700 | 0.1–1 | 5–20 min | - |
| 500–800 | 10–200 | 0.1–0.3 s | ||
| 850–1000 | >103 | 0.5–2 s | ||
| Liquefaction | 200–400 | - | - | 5–20 MPa |
| Source | Density at 15 °C (kg/m3) | Viscosity at −20 °C Maximum 8 mm2 s−1 | Flash Point (C) | Heating Value (MJ/Kg) | Sulphur Total (wt%) | References |
|---|---|---|---|---|---|---|
| Microalgae | 1380 | 2.8 | 68 | 44.0 | 0.27 | Bwapwa, et al. [129] |
| Jatropha | 751–850 | 3.66 | 46.5 | 44.3 | - | Li, et al. [130] |
| Tung | 839 | - | 39 | 42.3 | - | McAfee [131] |
| Soyabean | >775 | - | >38 | 43.4 | - | Li, Coppola, Rine, Miller and Walker [130] |
| Tallow | 758 | 5.3 | 55 | 44.1 | <0.001 | Corporan, et al. [132] |
| Castor | 758 | 5.3 | 55 | - | 0 | Liu, et al. [133] |
| Camelina | 751 | 3.3 | 43 | 44.1 | <0.001 | Corporan, Edwards, Shafer, DeWitt, Klingshirn, Zabarnick, West, Striebich, Graham and Klein [132] |
| Alcohol (C2 and C5) | 757.1 | 4.795 | 47.5 | - | - | Scheuermann, et al. [134] |
| Used cooking oil | 760 | 3.8 | 42 | 44 | <0.00003 | Buffi, et al. [135] |
| Jet-A (ASTM D1655) | 775–840 | 8 | 38 | 42.8 | 0.30 | Gutiérrez-Antonio, et al. [136] |
| Jet-A1 (ASTM D 1655) | 775–840 | 8 | 38 | 42.8 | 0.30 | Gutiérrez-Antonio, Gómez-Castro, de Lira-Flores and Hernández [136] |
| Microalga | Product | Yield (%) | References |
|---|---|---|---|
| Chlorella sp. | Bio-jet | 76 | Gómez-De la Cruz, et al. [137] |
| Desmodesmus sp. | Bio-oil | 49.40 | Garcia Alba, et al. [138] |
| Chlorella PTCC 6010 | Bio H2 | 41.46 | Samiee-Zafarghandi, et al. [139] |
| Schizochytrium sp. ABC 101 | Bio jet | 20.4 | Kim, et al. [140] |
| Chlamydomonas reinhardtii | Bio H2 | 56 | Fakhimi and Tavakoli [122] |
| Arthrospira platensis, Chlorella marina, Ulva linza | Bio ethanol | 45.49 | Ismail, et al. [141] |
| Anabaena sp. | Bio H2 | 57.6 | Vargas, Santos, Zaiat and Calijuri [123] |
| Nannochloropsis oculate and Tetraselmis suecica | Bio ethanol | 7.26 | Reyimu and Özçimen [142] |
| Aurantiochytrium sp. | Biodiesel | 89.5 | Kim, et al. [143] |
| Chlorella sorokiniana | Bio oil | 62 | Shirazi, et al. [144] |
| Botryococcus braunii | Biodiesel | 95 | Hidalgo, et al. [145] |
| Nannochloropsis gaditana | Biodiesel | 96.4 | Im, et al. [146] |
| Chlorella vulgaris | Biodiesel | 100 | Levine, et al. [147] |
| Microalgal Feedstock | TGA Operating Conditions | References | |||
|---|---|---|---|---|---|
| Initial Cell Mass (mg) | Temperature (°C) | Heating Rate (°C min −1) | Inert Gas Flow Rate (mL/min) | ||
| Dunaliella tertiolecta | 10 | 900 | 5–40 | 50 | Shuping, et al. [150] |
| Chlorella sp. | 100–1000 | 800 | 15 | 8000 | Rizzo, et al. [151] |
| Chlorella vulgaris | 10–50 | 800 | 5–40 | 100 | Agrawal and Chakraborty [152] |
| Chlorococcum humicola | - | 700 | 15–70 | - | Peng, et al. [153] |
| Consortia growth | 20 | 800 | 5–40 | 30 | Kim, et al. [154] |
| Chlorella protothecoides | - | 800 | 15–80 | 60 | Peng, Wu, and Tu [153] |
| Nannochloropsis gaditana | 4–24 | 1200 | 40 | 50–150 | Sanchez-Silva, et al. [155] |
| Isochrysis sp. | 5 | 900 | 5–25 | 100 | Zhao, et al. [156] |
| Microalgae Strains | Technology Used | Targeted Genes | Result | References |
|---|---|---|---|---|
| Chlamydomonas reinhardtii | de novo synthesis | Ferrochelatase gene and leghaemoglobin gene | Enhancement in hemH-lba protein | Wu, et al. [175] |
| Nannochloropsis oceanica | Overexpression of No MCAT (Malonyl CoA-acyl carrier protein transacylase) | Malonyl CoA-acyl carrier protein transacylase | Increment of the composition of eicosapentaenoic acid (C20:5) by 8% | Chen, et al. [176] |
| Mychonastesafer | Cloning and expression of 3-ketoacyl-coA synthase gene | 3-ketoacyl-coA synthase gene | Lipid content enhancement | Fan, et al. [177] |
| Chlamydomonas reinhardtii | dzCas9/sgRNA system suppressed the exogeneous gene | CrPEPC1 (CRISPRi mediated phosphoenol pyruvate carboxylase regulation) | Increment of CrCIS activity | Deng, et al. [178] |
| Dunaliella parva | Proteome analysis by iTRAQ | Upregulation of protein and downregulation of protein assisted genes | Proteins | Shang, et al. [179] |
| Scenedesmus sp. IITRIND2 | Physiological and metabolic elucidation under high saline condition | Upregulation of protein | Lipid enhancement | Arora, et al. [180] |
| Phaeodactylum tricornutum | De novo transcriptome analysis | Recombinant plastid electroporation | Promising role in AGPAT (1-acyl-sn-glycerol-3-phosphate acyltransferase) | Balamurugan, et al. [181] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasistha, S.; Khanra, A.; Rai, M.P.; Khan, S.A.; Ma, Z.; Munawaroh, H.S.H.; Tang, D.Y.Y.; Show, P.L. Exploring the Pivotal Significance of Microalgae-Derived Sustainable Lipid Production: A Critical Review of Green Bioenergy Development. Energies 2023, 16, 531. https://doi.org/10.3390/en16010531
Vasistha S, Khanra A, Rai MP, Khan SA, Ma Z, Munawaroh HSH, Tang DYY, Show PL. Exploring the Pivotal Significance of Microalgae-Derived Sustainable Lipid Production: A Critical Review of Green Bioenergy Development. Energies. 2023; 16(1):531. https://doi.org/10.3390/en16010531
Chicago/Turabian StyleVasistha, Shrasti, Anwesha Khanra, Monika Prakash Rai, Shakeel Ahmad Khan, Zengling Ma, Heli Siti Halimatul Munawaroh, Doris Ying Ying Tang, and Pau Loke Show. 2023. "Exploring the Pivotal Significance of Microalgae-Derived Sustainable Lipid Production: A Critical Review of Green Bioenergy Development" Energies 16, no. 1: 531. https://doi.org/10.3390/en16010531
APA StyleVasistha, S., Khanra, A., Rai, M. P., Khan, S. A., Ma, Z., Munawaroh, H. S. H., Tang, D. Y. Y., & Show, P. L. (2023). Exploring the Pivotal Significance of Microalgae-Derived Sustainable Lipid Production: A Critical Review of Green Bioenergy Development. Energies, 16(1), 531. https://doi.org/10.3390/en16010531