Abstract
Second-generation bioethanol production’s main bottleneck is the need for a costly and technically difficult pretreatment due to the recalcitrance of lignocellulosic biomass (LCB). Chemical pulping can be considered as a LCB pretreatment since it removes lignin and targets hemicelluloses to some extent. Chemical pulps could be used to produce ethanol. The present study aimed to investigate the batch ethanol production from unbleached Kraft pulp of Eucalyptus globulus by separate hydrolysis and fermentation (SHF). Enzymatic hydrolysis of the pulp resulted in a glucose yield of 96.1 ± 3.6% and a xylose yield of 94.0 ± 7.1%. In an Erlenmeyer flask, fermentation of the hydrolysate using Saccharomyces cerevisiae showed better results than Scheffersomyces stipitis. At both the Erlenmeyer flask and bioreactor scale, co-cultures of S. cerevisiae and S. stipitis did not show significant improvements in the fermentation performance. The best result was provided by S. cerevisiae alone in a bioreactor, which fermented the Kraft pulp hydrolysate with an ethanol yield of 0.433 g·g−1 and a volumetric ethanol productivity of 0.733 g·L−1·h−1, and a maximum ethanol concentration of 19.24 g·L−1 was attained. Bioethanol production using the SHF of unbleached Kraft pulp of E. globulus provides a high yield and productivity.
1. Introduction
According to REN21 (Renewable Energy Policy Network for the 21st Century), in 2016, fossil fuels corresponded to 79.5% of the total final energy consumption [1]. Biofuels appear as a potential renewable energy source in replacing fossil fuels, particularly given their much lower greenhouse gases emissions [2,3].
Bioethanol is currently the most produced biofuel, corresponding to about 74% of the 142.7 billion liters of biofuel produced in 2017. The US and Brazil are the biggest producers, being responsible for 57% and 27% of ethanol production worldwide, respectively [1]. However, bioethanol is almost entirely produced from food crops, i.e., first-generation bioethanol-sugarcane in Brazil, corn in the US, and sugar beet in the EU, resulting in a food–fuel competition [4,5].
Alternatively, bioethanol can be produced from lignocellulosic biomass (LCB), which is known as second-generation bioethanol (2G bioethanol). LCB comprises different types of biomass such as energy crops (e.g., perennial grasses), agricultural residues (e.g., wheat straw, corn stover, and sugarcane bagasse), forest materials (mainly woody materials), and the organic fraction of municipal solid wastes, as recently reviewed by [6]. Therefore, 2G bioethanol does not raise concern about food sustainability, has a low and stable price, and practically does not demand extra land [6,7]. There are some facilities producing 2G bioethanol on a commercial scale. However, large-scale production still faces some technological barriers that must be overcome in order to achieve a cost-competitive production [8]. Due to the recalcitrance of LCB, a costly pretreatment step is required, which is the main technological bottleneck of 2G bioethanol production. The release of enzymatic and fermentation inhibitors during pretreatment is another limitation [9].
Pulp and paper mills have the infrastructures and logistics to handle LCB, and chemical mills employ technology required for LCB fractionation and conversion [10]. Bioethanol has been produced from different feedstocks such as Kraft pulp, spent sulfite liquor, and pulp and paper sludge [11]. Chemical pulping processes can be considered as a LCB pretreatment since they promote delignification and target hemicelluloses to some degree [12]. Chemical pulping represents about 77% of the virgin pulps produced globally, and more than 95% of these chemical pulps are Kraft pulps [13]. These pulps are produced by Kraft pulping involving the reaction of white liquor, i.e., an alkaline aqueous solution of sodium hydroxide and sodium sulfide with a pH of 14, with lignin at high temperature (150–170 °C). This reaction promotes lignin breakdown and degradation with the release of phenolic fragments, removing almost 90% of the lignin from the wood. Kraft pulping also leads to hemicelluloses and some cellulose loss and decreases the degree of polymerization of cellulose [14]. The utilization of Kraft pulping as a pretreatment method for LCB has many advantages such as low sugar degradation, pulps free of inhibitors such as furfural and hydroxymethylfurfural (HMF), the recovery of chemicals used, and finally energy production in the recovery boiler [15,16].
The enzymatic hydrolysis of Kraft pulp of sweet sorghum bagasse [17], pine, poplar, birch, beech, and wheat straw [16,18], hemp [18], eucalyptus [19], moso bamboo [20], spruce, and birch–aspen mixture [21] has been studied. Hydrolysates with sugar profiles adequate for fermentation have been obtained, proving the viability of Kraft pulping as a LCB pretreatment. Bioethanol production from Kraft pulp by several fermentation configurations was studied, namely separate hydrolysis and fermentation (SHF) [22,23], simultaneous saccharification and fermentation (SSF) [15,24,25,26,27], and consolidated bioprocessing [28].
Besides hexose sugars, hydrolysates also have a high content in pentoses, mainly xylose, which can reach 25%, meaning that pentoses fermentation is necessary to attain an economically viable 2G bioethanol production [7,29]. Scheffersomyces stipitis was well as Saccharomyces cerevisiae have already been tested for bioethanol production from different LCB feedstocks, including eucalypt spent sulfite liquor [30,31,32], grape skins [33], sugarcane bagasse [34,35], cardoon, and rockrose [36]. The co-culture of hexose- and pentose-fermenting yeasts is a potential solution for this problem, since most well-known natural microorganisms are not able to efficiently ferment both sugars. By using this type of co-culture, the sugars consumption and ethanol production ratios are expected to increase and, consequently, process productivity should also increase [37].
The present work aimed to study ethanol production from the unbleached Kraft pulp of Eucalyptus globulus using an SHF configuration. Firstly, the Kraft pulp was enzymatically hydrolyzed, and the obtained hydrolysate was used for Erlenmeyer flask assays with Saccharomyces cerevisiae or Scheffersomyces stipitis to evaluate the yeasts’ growth and ethanol fermentation. Then, the fermentation of the Kraft pulp hydrolysate by the sequential or simultaneous co-culture of both yeasts in an Erlenmeyer flask was studied. Finally, bioreactor assays using S. cerevisiae or sequential co-culture with Saccharomyces cerevisiae or Scheffersomyces stipitis were performed. To our knowledge, this is the first time that SHF configuration and a co-culture strategy are applied for ethanol production from unbleached Kraft pulp of Eucalyptus globulus. The results of this study should help to assess the feasibility of converting pulp and paper mills into integrated biorefineries, exploring Kraft pulping as a pretreatment method.
2. Materials and Methods
2.1. Kraft Pulp
Unbleached Kraft pulp of Eucalyptus globulus was supplied by The Navigator Company (Cacia, Portugal). The pulp was composed by 78 wt % cellulose, 19 wt % hemicelluloses, and 2.4 wt % lignin (data provided by RAIZ–Instituto da Floresta e do Papel). Before being submitted to enzymatic hydrolysis, the paper pulp was washed and then centrifuged to remove the excess water.
2.2. Enzymatic Hydrolysis
Kraft pulp enzymatic hydrolysis batch tests were performed using a cellulase and hemicellulases cocktail supplied by The Navigator Company and RAIZ-Instituto de Investigação da Floresta e do Papel. Tests were performed with an enzymatic activity of approximately 195 FPU·mL−1, an enzymatic load of 25 FPU·g of carbohydrate−1, a stirring rate of 100 rpm, at 50 °C, and pH between 4.5 and 5.5 (adjusted by adding H2SO4 3 M and NaOH 2 M solutions). In each batch, 200 g of pulp were suspended in 4350 mL water; then, 200 mL of enzymatic preparation were added, and the batch was carried out for 24 h. After that, the hydrolysate was filtered using filter paper 545 with particle retention of 12–15 μm (VWR, Vienna, Austria), and, finally, all hydrolysates were mixed together and sterilized by autoclaving for 20 min at 121 °C.
2.3. Microorganisms
Saccharomyces cerevisiae PYCC 5246 (ATCC 24860) was gently supplied by Portuguese Yeast Culture Collection. Scheffersomyces stipitis NRRL Y-7124 was gently supplied by the Agricultural Research Service Culture Collection at National Center for Agricultural Utilization Research, USDA. Both yeasts were grown at 28 °C and maintained at 4 °C in YM (Yeast Mold) solid medium. All media and material were sterilized by autoclaving for 20 min at 121 °C for 20 min.
2.4. Culture Media
YM solid medium was prepared with 3.0 g·L−1 yeast extract, 3.0 g·L−1 malt extract, 5.0 g·L−1 peptone, 10.0 g·L−1 glucose, and 20.0 g·L−1 of agar. YM liquid medium was prepared with 3.0 g·L−1 yeast extract, 3.0 g·L−1 malt extract, 5.0 g·L−1 peptone, and 10.0 g·L−1 glucose. The pH of both YM media was adjusted to 5.5.
2.5. Pre-Inocula and Inocula
Pre-inocula were prepared by transferring a colony from maintenance YM solid medium to YM liquid medium and were incubated at 180 rpm for 24 h at 28 °C in an orbital shaker (B. Braun Certomat S, Melsungen, Germany). Inocula were prepared by the transference of pre-inocula to YM liquid medium in order to obtain an initial biomass concentration of 0.200 g·L−1. The inocula were incubated for 14 h in the described conditions.
For the Erlenmeyer flask assays, the pre-inocula were prepared in 50 mL Falcon tubes with 10 mL of YM liquid medium and the inocula were prepared in 100 mL Erlenmeyer flasks with 40 mL of YM liquid medium. For the bioreactor assays, the pre-inocula were prepared in 100 mL Erlenmeyer flasks with 40 mL of YM liquid medium, and the inocula were prepared in 500 mL Erlenmeyer flasks with 300 mL of YM liquid medium.
2.6. Fermentation
The fermentation media were prepared with 65% (v/v) of Kraft pulp hydrolysate, 25% (v/v) of a supplementary medium (2.0 g·L−1 (NH4)2HPO4, 1.0 g·L−1 (NH4)2SO4, 0.5 g·L−1 MgSO4.7H2O, and 2.5 g·L−1 yeast extract) and 10% (v/v) of inoculum. The pH was adjusted to 5.5 before inoculation. Throughout the fermentations, samples were taken to monitor the pH and concentration of biomass, glucose, xylose, and ethanol.
2.6.1. Erlenmeyer Flask Assays
The Erlenmeyer flask assays were carried out in 250 mL Erlenmeyer flasks with a working volume of 100 mL and incubated at 28 °C and 180 rpm in an orbital shaker (B. Braun Certomat S, Melsungen, Germany). Four Erlenmeyer flask assays were performed: one with S. cerevisiae; one with S. stipitis; one with a sequential co-culture of both yeasts; and one with a simultaneous co-culture of both yeasts. The initial biomass concentration in all assays was about 0.200 g·L−1. In the assay with simultaneous co-culture, both strains were inoculated in 1:1 ratio. In the assay with sequential co-culture, S. cerevisiae was firstly inoculated and S. stipitis was inoculated after 24 h of fermentation with a volume of inoculum that would allow a concentration of S. stipitis of 0.200 g·L−1.
2.6.2. Bioreactor Assays
The bioreactor assays were carried out with a working volume of 3 L in a 5 L BIOSTAT Aplus bioreactor (Sartorius Stedim Biotech, Göttingen, Germany) with automatic control of stirring, temperature, and pH by micro-DCU software and data acquisition by MFCS/DA 3.0 system (Sartorius Stedim Systems, Göttingen, Germany). The pH was measured using an electrode EasyFerm Plus K8 325 (Hamilton, Reno, NV, USA) and controlled to 5.50 ± 0.05 through the addition of KOH 5 M and H2SO4 1 M. The temperature was controlled at 28 °C and the stirring was controlled at 180 rpm by two si-blade disk impellers and four baffles.
Two bioreactor assays were performed: one with S. cerevisiae and one with sequential co-culture of S. cerevisiae and S. stipitis. Inoculation procedures were made as described for Erlenmeyer tests. In the sequential co-culture test, from 15 min before the inoculation of S. stipitis until the end of the fermentation, the bioreactor was aerated using a ring sparger and an air flow of 50 mL·min−1 (4800 series flow controller, Brooks Instrument, Hatfield, PA, USA).
2.7. Analytical Methods
The pH of the samples was measured using an electrode InPro 3030/200 (Mettler Toledo, Columbus, OH, USA) connected to a benchtop meter sensION+ MM340 (Hach, Loveland, CO, US).
Biomass was monitored measuring optical density at 620 nm (UVmini-1240, Shimadzu, Tokyo, Japan) and further converted into concentration using a calibration curve of optical density versus biomass dry weight.
Glucose, xylose, and ethanol concentrations were analyzed by HPLC (high-performance liquid chromatography). Samples were centrifuged and filtered off with centrifuge tube 0.20-m filters CoStar Spin-X (Corning, NY, USA) in MiniSpin centrifuge (Eppendorf, Hamburg, Germany) for 10 min at 8000 rpm. After dilution, the samples were injected by autosampler L-2200 (Hitachi, Ltd., Chiyoda, Japan) to a Rezex ROA-Organic Acid H+ (8%) 50 × 7.8 mm ion-exchange column (Phenomenex, Torrance, CA, USA) at 65 °C (oven Gecko 2000, CIL Cluzeau, Sainte-Foy-la-Grande, France) and detected by a refraction index detector L-2490 (Hitachi, Chiyoda, Japan). The injection volume was 10 μL and the eluent used was H2SO4 0.01 N, with a flow rate of 0.5 mL·min−1 (pump L-2130, Hitachi, Chiyoda, Japan). The concentration of the analyzed compounds was determined using a standard calibration curve obtained by injecting standards for all analyzed compounds.
The chemical oxygen demand (COD) was measured with a Spectroquant Kit (Merck, Darmstadt, Germany), and the solutions used were prepared according to Standard Methods [38].
2.8. Calculations
The hydrolysis yield on glucose (Yglucose) and the hydrolysis yield on xylose (Yxylose) were calculated according to the potential glucose or xylose in the unbleached Kraft pulp using Equations (1) and (2), respectively.
The specific growth rate, µ (h−1), was calculated by determining the slope of the linear regression obtained after plotting the natural logarithm of biomass concentration versus time during the exponential phase.
The volumetric substrate consumption rates, rglucose (g·L−1·h−1) or rxylose (g·L−1·h−1), were calculated by determining the module of the slope of the linear regression obtained after plotting each substrate concentration versus time.
The volumetric ethanol productivity, Prodvol (g·L−1·h−1), was calculated using Equation (3) from the beginning of the fermentation until maximum ethanol concentration was achieved.
The ethanol yield, Yethanol/substrate (g·g−1), and the biomass yield, Ybiomass/substrate (g·g−1), were calculated using Equations (4) and (5), respectively, considering both glucose and xylose as substrate.
The conversion efficiency (%) was determined using Equation (6), considering a maximum theoretical ethanol yield of 0.511 g·g−1 [39].
3. Results
3.1. Enzymatic Hydrolysis
The enzymatic hydrolysis of unbleached Kraft pulp of E. globulus resulted in hydrolysates with 65.45 ± 0.80 g·L−1 of glucose and 16.02 ± 1.75 g·L−1 of xylose, corresponding to a Yglucose of 96.1 ± 3.6% and a Yxylose of 94.0 ± 7.1%. These values are very similar to those reported by Huang et al. (2017), 91.5% and 93.2%, using the unbleached Kraft pulp of Phyllostachys heterocycle as feedstock [23]. Arévalo et al. (2017) reported hydrolysis yields in terms of glucose of 75.9% and 80.0% for the enzymatic hydrolysis of E. globulus wood pretreated with autohydrolysis and diluted acid hydrolysis, respectively [40]. After autoclaving, the final hydrolysate had a composition in glucose and xylose of 64.37 g·L−1 and 11.75 g·L−1, respectively. This hydrolysate was used for the subsequent fermentation assays.
3.2. Erlenmeyer Flask Mono-Culture Assays
In the assay with S. cerevisiae (Figure 1A), after a very short lag phase, glucose started to be consumed for biomass growth and also for ethanol production. The maximum ethanol concentration of 19.81 ± 0.15 g·L−1 was reached at 8.9 h simultaneously with glucose exhaustion. Biomass concentration increased while glucose was being consumed and remained constant until 24.0 h, when ethanol started to be assimilated, resulting in a little increase in biomass concentration. One of the reasons pointed out to explain the high tolerance of S. cerevisiae to ethanol is its ability to respire ethanol when the concentration of fermentable sugars has dropped [41]. In this assay, pH decreased simultaneously with the increase in biomass and ethanol concentrations (Figure 1A), which indicates that this decrease is probably due to the production of CO2 during cell growth and alcoholic fermentation. During the assay, S. cerevisiae presented a very slight consumption of xylose with very low rxylose, 0.193 ± 0.003 g·L−1·h−1, since this yeast is known to present very low expression levels of the genes required for xylose utilization [42].
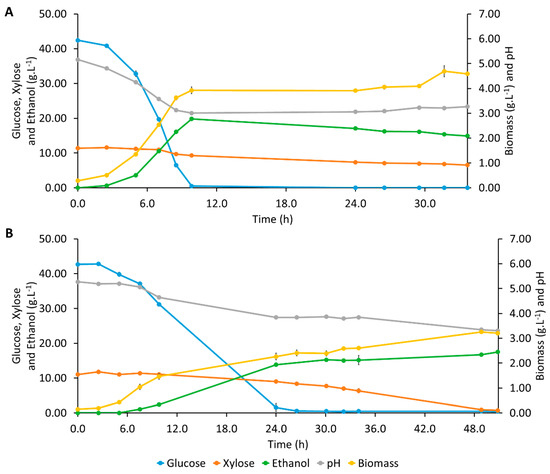
Figure 1.
Profiles of pH and concentrations of glucose, xylose, ethanol, and biomass during Erlenmeyer flask fermentations assays with mono-culture: (A) Mono-culture of Saccharomyces cerevisiae; (B) Mono-culture of Scheffersomyces stipitis.
Although a high Prodvol, 2.01 ± 0.01 g·L−1·h−1, and also a Yethanol/substrate, 0.450 ± 0.009 g·g−1, corresponding to a conversion efficiency of 88.3 ± 1.7%, were observed (Table 1), most xylose was still present in the fermentation medium by the end of the fermentation. In order to promote the utilization of all glucose and xylose, a fermentation by S. stipitis NRRL Y-7124, a hexose- and pentose-fermenting yeast, was evaluated (Figure 1B). In this assay, glucose was exhausted around 28.0 h and, after that, the consumption rate of xylose increased. When the culture medium contains both glucose and xylose, glucose repression through inhibition of the enzymes of the membrane xylose transport system occurs, and there is a preference for glucose as substrate [43,44]. Ethanol concentration increased throughout the fermentation, attaining a maximum ethanol concentration of 17.50 ± 0.09 g·L−1 at 50.8 h, corresponding to a Prodvol of 0.344 ± 0.002 g·L−1·h−1 and a Yethanol/substrate of 0.333 ± 0.011 g·g−1. The production of ethanol became slower when glucose was depleted, and xylose consumption started. Biomass concentration also kept increasing, although more slowly, when xylose uptake started. The pH decreased throughout the fermentation due to the CO2 release by simultaneous biomass growth and ethanol production.

Table 1.
Kinetic and stoichiometric parameters for all fermentation assays.
In the S. stipitis assay, a higher rxylose and full consumption of both glucose and xylose were observed (Figure 1B), yielding a higher percentage of consumed sugars, 97.9 ± 0.2%, than that attained by S. cerevisiae previous assay. However, lower ethanol and biomass concentrations were attained, providing lower biomass and ethanol yields (Table 1). This might be indicative that S. stipitis utilized some of the carbon flux for maintenance pathways, other than growth and ethanol production, which were possibly pathways related to resistance to face the inhibitors present in the hydrolysate. S. stipitis is known to be highly sensitive to several inhibitors present in hydrolysates, namely formic, acetic and levulinic acids, phenolics, and furfural [45]. Although yeasts can detoxify microbial inhibitors, such as furfural and HMF, the detoxification mechanisms compete for key enzymes and cofactors needed to channel the carbon flow to respiration or ethanol production [46]. On the other hand, S. cerevisiae is commonly used as a wine deacidifying yeast and, according to Geng et al. (2010), S. cerevisiae ATCC 24860 showed high resistance to inhibitors usually present in LCB hydrolysates (i.e., formic acid, acetic acid, furfural, and HMF) [47].
S. stipitis also showed lower rglucose, 1.98 ± 0.03 g·L−1·h−1, and Prodvol, 0.344 ± 0.002 g·L−1·h−1, than S. cerevisiae (Table 1). Hence, in order to enhance the consumption of sugars until the full exhaustion of glucose and xylose and, consequently, increase the amount of ethanol produced and the fermentation efficiency, the co-culture of both yeasts was studied next.
3.3. Erlenmeyer Flask Co-Culture Assays
In a sequential co-culture assay, in order to alleviate the repression of xylose consumption by glucose, S. cerevisiae was inoculated first, and S. stipitis was inoculated at 24 h of fermentation when glucose was already exhausted (Figure 2A). A maximum ethanol concentration of 20.03 ± 0.61 g·L−1 was detected at 23.7 h of fermentation, at the same time glucose was exhausted and before S. stipitis inoculation, meaning that all the ethanol was produced exclusively by S. cerevisiae fermentation. Thus, the Yethanol/substrate observed, 0.464 ± 0.009 g·g−1, was similar to that obtained in S. cerevisiae previous culture, 0.450 ± 0.009 g·g−1, and only 84.3 ± 0.3% of the sugars were consumed at the time of maximum ethanol concentration (Table 1).
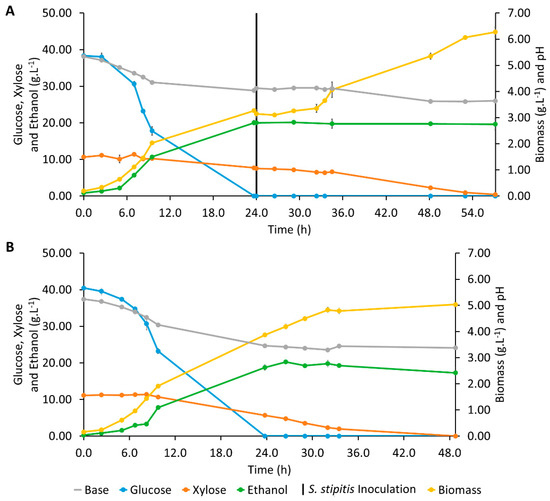
Figure 2.
Profiles of pH and concentrations of glucose, xylose, ethanol, and biomass during Erlenmeyer flask fermentations assays with co-culture of Saccharomyces cerevisiae and Scheffersomyces stipitis: (A) Sequential co-culture; (B) Simultaneous co-culture.
After S. stipitis inoculation, no more ethanol production was observed, which was most likely because the aeration conditions were not adequate for fermentation to ethanol by S. stipitis, since for this metabolism, this yeast requires specific microaerophilic conditions [48,49]. Additionally, product inhibition might have happened, according to Gutiérrez-Rivera et al. (2015), who investigated the sequential co-culture of these two yeasts for ethanol production from sugarcane bagasse, and S. stipitis inhibition when ethanol concentrations were still lower than 30 g·L−1 was observed [50]. In this assay, xylose was consumed slowly until 34.5 h of fermentation when rxylose increased to 0.309 ± 0.003 g·L−1·h−1, which is a value higher than that presented by the S. cerevisiae previous assay, but still lower than that observed for the S. stipitis assay (Table 1). A second lag phase was observed during the first 8 h after S. stipitis inoculation, followed by an exponential biomass growth. The pH, which remained constant for the same period, also started to decrease (Figure 2A). This lag phase was longer than that observed in the assay with S. stipitis (Figure 1B), since no more glucose was present in the medium at the time of S. stipitis inoculation. The inoculum was prepared with glucose and without xylose, meaning that S. stipitis had more difficulty to adapt, needing to synthesize the enzymatic machinery required to metabolize xylose. Considering the small amount of xylose consumed and the great increase in biomass observed, the latter could additionally be explained by the growth of S. cerevisiae due to the consumption of other compounds present in the hydrolysate medium, as suggested by preliminary COD (chemical oxygen demand) analysis. Figure 3A shows the evolution of COD determined according to Standard Methods and COD calculated from the concentrations of glucose, xylose, and ethanol obtained in the HPLC in the S. cerevisiae assay in Erlenmeyer. The evolution of Experimental COD and Calculated COD got closer as the fermentation progressed. The higher decrease observed on Experimental COD could be an indication that S. cerevisiae consumed other compounds present in the hydrolysate than glucose and xylose. Figure 3B shows the evolution of Experimental COD and calculated COD in the assay with S. stipitis. The difference between both CODs is approximately the same during the fermentation, indicating that probably only glucose and xylose were consumed by S. stipitis.
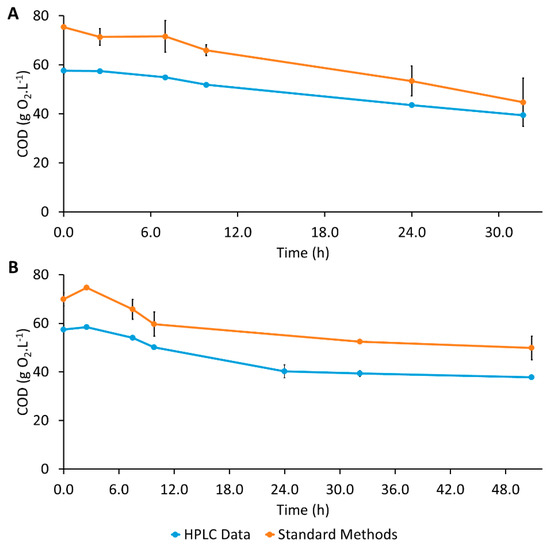
Figure 3.
Chemical oxygen demand (COD) evolution in Erlenmeyer assays: (A) Saccharomyces cerevisiae assay; (B) Scheffersomyces stipitis assay.
Although a high conversion efficiency of 91.0 ± 1.8% was obtained in sequential co-culture, it is exclusively attributed to S. cerevisiae metabolic activity; hence, simultaneous co-culture was investigated next, and the results are present in Figure 2B. Glucose exhaustion was detected around 24.0 h of fermentation but given the rglucose, 3.83 ± 0.36 g·L−1·h−1, it is certain that glucose was fully consumed around 16.0 h of fermentation. Therefore, when S. cerevisiae and S. stipitis were cultivated simultaneously, glucose was consumed more slowly than when S. cerevisiae was alone but faster than in S. stipitis assay (Table 1). As it was observed in the assay with S. stipitis, the rxylose increased after glucose exhaustion due to the end of glucose repression, and xylose was fully consumed by the end of the fermentation. The rxylose observed, 0.410 ± 0.003 g·L−1·h−1, was the highest among all the Erlenmeyer flask assays (Table 1). The maximum ethanol concentration, 20.25 ± 0.29 g·L−1, which was achieved at 26.5 h (Figure 2B), was similar to that observed for S. cerevisiae, although in the simultaneous co-culture assay, there was a higher percentage of consumed sugars, 90.8 ± 0.3%. Nevertheless, the maximum ethanol concentration was achieved 13 h later, and a lower ethanol productivity of 0.754 ± 0.021 g·L−1·h−1 was observed. A similar ethanol yield, 0.426 ± 0.016 g·g−1, but higher biomass yield, 0.086 ± 0.002 g·g−1, were observed (Table 1). Therefore, it can be concluded that carbon flow was deviated more toward growth when both yeasts were present in the fermentation medium; this was most likely by S. stipitis since, contrary to S. cerevisiae, which regulates fermentation depending on sugar concentration, this yeast is a Crabtree-negative one. It shows a fully respiratory metabolism under aerobic conditions, even if glucose is present in excess, favoring cell growth. Papini et al. (2012) reported that S. stipitis produces ethanol when oxygen becomes limiting, since reduced oxygen tension induces pyruvate decarboxylase and alcohol dehydrogenase activity [51]. The optimum oxygen concentration is the main bottleneck of S. stipitis bioethanol production [45], and a new strategy of bioreactor aeration was just described for bioethanol fermentation [30].
At the end of the fermentation, biomass grew very slightly, and slow ethanol consumption was observed (Figure 2B). Ethanol consumption after sugar exhaustion in simultaneous co-culture has already been reported in the literature [52,53].
3.4. Bioreactor Assays
The effect of increasing the working volume—from a 100 mL Erlenmeyer flask to a 3 L bioreactor—on growth and fermentation was studied next. In the bioreactor assay with S. cerevisiae, better fermentation performance was expected, since the pH was automatically controlled to 5.5. While in the correspondent Erlenmeyer assay, the pH dropped to values as low as 3.01 and, although S. cerevisiae ATCC 24860 presents a high pH tolerance, this yeast strain is known to be affected when the pH drops below 4 [47].
Ethanol concentration increased rapidly until 9.3 h, and when glucose was exhausted at 12.8 h, 16.39 g·L−1 of ethanol were present in the medium (Figure 4A). A maximum ethanol concentration of 19.24 g·L−1 was observed 13.2 h after glucose depletion. Xylose concentration decreased very slightly during this period of fermentation, corresponding to an rxylose of 0.385 g·L−1·h−1. Although S. cerevisiae ATCC 24860 has been reported to have the ability to produce a slight amount of ethanol from xylose [54], the increase of ethanol concentration after glucose depletion was most likely due to the fermentation of other compounds of the hydrolysate by S. cerevisiae.
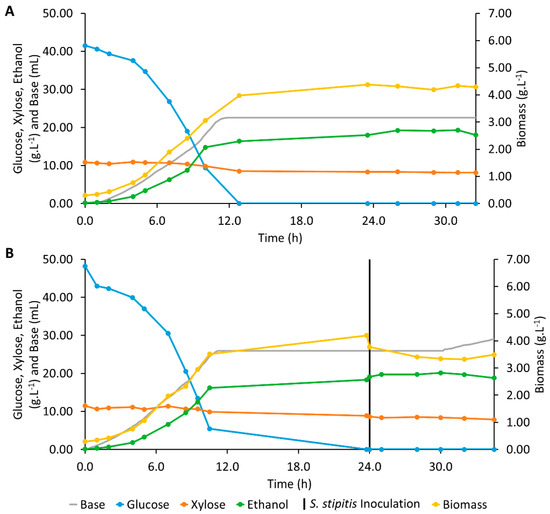
Figure 4.
Profiles of base addition and concentrations of glucose, xylose, ethanol, and biomass during bioreactor fermentations assays: (A) Mono-culture of Saccharomyces cerevisiae; (B) Sequential co-culture of Saccharomyces cerevisiae and Scheffersomyces stipitis.
The automatic addition of KOH to adjust the pH of the fermentation medium was registered for the first 12 h of operation, corresponding to the increase in biomass and ethanol concentrations and due to CO2 release.
Although the maximum values of ethanol concentration achieved in both fermentations (Erlenmeyer and bioreactor) by S. cerevisiae were very close, in this assay with S. cerevisiae in a bioreactor, a μ of 0.251 h−1, a rglucose of 4.66 g·L−1·h−1, a Prodvol of 0.733 g·L−1·h−1, and a Yethanol/substrate of 0.433 g·g−1, corresponding to a conversion efficiency of 84.8%, were observed. These values are lower than that obtained in the corresponding Erlenmeyer flask assay (Table 1). When increasing the scale of a biological process, problems in obtaining a perfect mixture might occur, leading to decreased yields and productivities. The hydrolysate has a complex composition, containing some particulate suspended matter and presenting some viscosity. Hence, a 30-fold increase in volume and a change in the shape of the fermentation vessel without increasing the stirring rate may have hindered mass transfer, resulting in a decrease of consumption and production rates and ethanol yield. Hence, future research should focus on studying the conditions, namely stirring rate, and fed-batch fermentation, for the maximum ethanol production from Kraft pulp hydrolysate using S. cerevisiae.
Table 2 presents the results of different S. cerevisiae fermentations using the same microbial strain, pretreatment, and/or feedstock as in the present work. Zambare et al. (2011 and 2012) used S. cerevisiae ATCC 24860 and an SHF configuration for ethanol production from Spartina pectinata and corn stover, respectively [55,56]. Both studies reported lower ethanol concentrations and productivities than the present work. However, a higher ethanol yield was obtained from extrusion-pretreated corn stover hydrolysates fermentation, 0.47 g·g−1. On the other hand, extrusion-pretreated Spartina pectinata hydrolysate fermentation resulted in an ethanol yield of 0.426 g·g−1, which is close to that obtained in this work. Similar to in the present work, Ko et al. (2012) and Monrroy et al. (2012) used E. globulus as feedstock and Kraft pulping as a pretreatment, but they employed a SSF configuration, reporting lower ethanol yields, 0.042 g·g d.w. wood−1 and 0.469 g·g d.w. pulp−1, respectively [15,24]. Kelbert et al. (2016) applied autohydrolysis as the pretreatment of E. globulus and used SHF configuration for the production of ethanol. Although a higher ethanol concentration was achieved, 39.7 g·L−1, lower ethanol productivity and yield were obtained, 0.551 g·L−1·h−1 and 0.38 g·g−1, when compared with this work [57].
Table 2.
Comparison between the results obtained for bioreactor assay with S. cerevisiae mono-culture in this work and in the literature.
One assay with sequential co-culture in bioreactor was performed (Figure 4B) and the microaerophilic conditions necessary for S. stipitis fermentation [48,49] were set after the inoculation of this yeast in order to activate xylose fermentation by S. stipitis. Similar to the assay in an Erlenmeyer flask, S. stipitis was inoculated in the fermentation medium at 24.0 h of fermentation, when all glucose had been consumed. After the inoculation of S. stipitis, a slight consumption of xylose was observed and the ethanol concentration increased very slowly, from 19.10 g·L−1 to 20.13 g·L−1, reaching this maximum ethanol concentration at 30.0 h. Therefore, contrary to the sequential co-culture assay in Erlenmeyer, ethanol production was verified after S. stipitis inoculation in the sequential co-culture assay in a bioreactor. However, the maximum ethanol concentration was only slightly higher than that observed with S. cerevisiae culture assays in both scales, and lower ethanol productivity, 0.668 g·L−1·h−1, and ethanol yield, 0.391 g·g−1, was observed. The low xylose consumption observed may be due to the S. stipitis inoculum, which was prepared in a medium that contained glucose but no xylose, forcing the culture to synthesize or at least to activate the enzymatic machinery required for xylose metabolization. Additionally, the airflow rate may not have been sufficiently low to promote xylose fermentation. After maximum ethanol concentration was attained, slight ethanol consumption was observed until the end of the fermentation. These consumptions resulted in an increase in biomass concentration with a simultaneous alkali addition to pH adjustment, reinforcing that the aeration of the medium was not adequate for changing the xylose metabolism of biomass growth and respiration to the ethanol fermentation by S. stipitis. Hence, S. cerevisiae and S. stipitis sequential co-culture should be further investigated through the optimization of aeration conditions for the xylose fermentation by S. stipitis.
Using the xylose fraction of the hydrolysate to produce high value-added compounds, such as succinic acid, lactic acid, and xylitol, instead of bioethanol, is a promising alternative. According to Yamakawa et al. (2018), using a hemicellulose fraction to produce high-value chemicals could improve the revenue of the biorefinery when compared with using both the cellulosic and hemicellulosic fractions of LCB for bioethanol production [8].
4. Conclusions
The present studied demonstrated that batch ethanol production from unbleached Kraft pulp of Eucalyptus globulus using an SHF configuration with a high yield and productivity is possible. Enzymatic hydrolysis of the Kraft pulp produced a hydrolysate with an adequate sugar profile for ethanol production and with high hydrolysis yields. S. cerevisiae showed better fermentation performance than S. stipitis. In an Erlenmeyer flask, the sequential co-culture of S. cerevisiae and S. stipitis showed no improvement, as no ethanol production was observed after inoculation with S. stipitis. Simultaneous co-culture resulted in the consumption of both glucose and xylose but with lower ethanol productivity. S. cerevisiae fermentation of Kraft pulp hydrolysate in bioreactor resulted in a slightly lower ethanol yield and productivity compared with the corresponding assay in Erlenmeyer flask, certainly resulting from mass transfer problems. Ethanol production was observed after S. stipitis inoculation in sequential co-culture at the bioreactor scale; however, no improvement in ethanol yield and productivities were attained.
Considering the present results, instead of good-quality E. globulus Kraft pulp, low-quality Kraft pulp or Kraft pulp obtained from wastes resulting from wood preparation before pulping should be tested for enzymatic hydrolysis and subsequent ethanol production in order to implement integrated biorefineries in the existing Kraft pulp mills.
Author Contributions
R.H.R.B. carried out the experiments, collected and analyzed data, as well as wrote the manuscript; M.S.T.A. helped with the experiments, L.S.S. and A.M.R.B.X. developed the concept, supervised the work, and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was developed within the scope of the project CICECO-Aveiro Institute of Materials, FCT Ref. UID/CTM/50011/2019, financed by national funds through the FCT/MCTES.
Acknowledgments
The authors are thankful to The Navigator Company and RAIZ—Instituto de Investigação da Floresta e do Papel for supplying the pulp, the enzymatic solution, and all the equipment required for the enzymatic hydrolysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- REN21. Renewables 2018 Global Status Report; REN21 Secretariat: Paris, France, 2018. [Google Scholar]
- Liew, W.H.; Hassim, M.H.; Ng, D.K.S. Review of evolution, technology and sustainability assessments of biofuel production. J. Clean. Prod. 2014, 71, 11–29. [Google Scholar] [CrossRef]
- Berni, M.D.; Dorileo, I.L.; Prado, J.M.; Forster-Carneiro, T.; Meireles, M.A.A. Advances in biofuel production. In Biofuels Production; Babu, V., Thapliyal, A., Patel, G.K., Eds.; Scrivener Publishing: Boston, MA, USA, 2014. [Google Scholar]
- Dutta, K.; Daverey, A.; Lin, J.-G. Evolution retrospective for alternative fuels: First to fourth generation. Renew. Energy 2014, 69, 114–122. [Google Scholar] [CrossRef]
- Manochio, C.; Andrade, B.R.; Rodriguez, R.P.; Moraes, B.S. Ethanol from biomass: A comparative overview. Renew. Sustain. Energy Rev. 2017, 80, 743–755. [Google Scholar] [CrossRef]
- Zabed, H.; Sahu, J.N.; Suely, A.; Boyce, A.N.; Faruq, G. Bioethanol production from renewable sources: Current perspectives and technological progress. Renew. Sustain. Energy Rev. 2017, 71, 475–501. [Google Scholar] [CrossRef]
- Zabed, H.; Sahu, J.N.; Boyce, A.N.; Faruq, G. Fuel ethanol production from lignocellulosic biomass: An overview on feedstocks and technological approaches. Renew. Sustain. Energy Rev. 2016, 66, 751–774. [Google Scholar] [CrossRef]
- Yamakawa, C.K.; Qin, F.; Mussatto, S.I. Advances and opportunities in biomass conversion technologies and biorefineries for the development of a bio-based economy. Biomass Bioenergy 2018, 119, 54–60. [Google Scholar] [CrossRef]
- Bhutto, A.W.; Qureshi, K.; Harijan, K.; Abro, R.; Abbas, T.; Bazmi, A.A.; Karim, S.; Yu, G. Insight into progress in pre-treatment of lignocellulosic biomass. Energy 2017, 122, 724–745. [Google Scholar] [CrossRef]
- Stoklosa, R.J.; Hodge, D.B. Integration of (hemi)-cellulosic biofuels technologies with chemical pulp production. In Biorefineries: Integrated Biochemical Processes for Liquid Biofuels; Qureshi, N., Hodge, D., Vertès, A.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Branco, R.; Serafim, L.S.; Xavier, A.M.R.B. Second generation bioethanol production: On the use of pulp and paper industry wastes as feedstock. Fermentation 2018, 5, 4. [Google Scholar] [CrossRef]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef]
- FAO. Yearbook of Forest Products 2015; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Lachenal, D. Kraft pulping. In Lignocellulosic Fibers and Wood Handbook; Belgacem, M.N., Pizzi, A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Monrroy, M.; García, J.R.; Mendonça, R.T.; Baeza, J.; Freer, J. Kraft pulping of Eucalyptus globulus as a pretreatment for bioethanol production by simultaneous saccharification and fermentation. J. Chil. Chem. Soc. 2012, 57, 1113–1117. [Google Scholar] [CrossRef]
- Buzała, K.; Przybysz, P.; Rosicka-Kaczmarek, J.; Kalinowska, H. Production of glucose-rich enzymatic hydrolysates from cellulosic pulps. Cellulose 2015, 22, 663–674. [Google Scholar] [CrossRef]
- Solihat, N.N.; Fajriutami, T.; Adi, D.T.N.; Fatriasari, W.; Hermiati, E. Reducing sugar production of sweet sorghum bagasse kraft pulp. In Proceedings of the AIP Conference Proceedings, Tangerang Banten, Indonesia, 3–5 October 2016. [Google Scholar]
- Buzała, K.P.; Kalinowska, H.; Przybysz, P.; Małachowska, E. Conversion of various types of lignocellulosic biomass to fermentable sugars using kraft pulping and enzymatic hydrolysis. Wood Sci. Technol. 2017, 51, 873–885. [Google Scholar] [CrossRef]
- Troncoso, E.; Castillo, R.; Valenzuela, R.; Reyes, P.; Freer, J.; Norambuena, M.; RodríGuez, J.; Parra, C. Chemical and microstructural changes in Eucalyptus globulus fibers subjected to four different pretreatments and their influence on the enzymatic hydrolysis. J. Chil. Chem. Soc. 2017, 62, 3442–3446. [Google Scholar] [CrossRef]
- Huang, C.; He, J.; Min, D.; Lai, C.; Yong, Q. Understanding the nonproductive enzyme adsorption and physicochemical properties of residual lignins in moso bamboo pretreated with sulfuric acid and Kraft pulping. Appl. Biochem. Biotech. 2016, 180, 1508–1523. [Google Scholar] [CrossRef]
- Novozhilov, E.V.; Sinel’nikov, I.G.; Aksenov, A.S.; Chukhchin, D.G.; Tyshkunova, I.V.; Rozhkova, A.M.; Osipov, D.O.; Zorov, I.N.; Sinitsyn, A.P. Biocatalytic conversion of kraft pulp using cellulase complex of Penicillium verruculosum. Catal. Ind. 2016, 8, 95–100. [Google Scholar] [CrossRef]
- Buzała, K.P.; Kalinowska, H.; Małachowska, E.; Przybysz, P. The utility of selected kraft hardwood and softwood pulps for fuel ethanol production. Ind. Crops Prod. 2017, 108, 824–830. [Google Scholar] [CrossRef]
- Huang, C.; He, J.; Chang, H.; Jameel, H.; Yong, Q. Coproduction of ethanol and lignosulfonate from moso bamboo residues by fermentation and sulfomethylation. Waste Biomass Valorization 2017, 8, 965–974. [Google Scholar] [CrossRef]
- Ko, C.-H.; Wang, Y.-N.; Chang, F.-C.; Chen, J.-J.; Chen, W.-H.; Hwang, W.-S. Potentials of lignocellulosic bioethanols produced from hardwood in Taiwan. Energy 2012, 44, 329–334. [Google Scholar] [CrossRef]
- Wistara, N.J.; Pelawi, R.; Fatriasari, W. The effect of lignin content and freeness of pulp on the bioethanol productivity of Jabon wood. Waste Biomass Valorization 2016, 7, 1141–1146. [Google Scholar] [CrossRef]
- Edgardo, A.; Carolina, P.; Manuel, R.; Juanita, F.; Baeza, J. Selection of thermotolerant yeast strains Saccharomyces cerevisiae for bioethanol production. Enzyme Microb. Technol. 2008, 43, 120–123. [Google Scholar] [CrossRef]
- Bauer, N.A.; Gibbons, W.R. Saccharification versus simultaneous saccharification and fermentation of kraft pulp. Int. J. Agric. Biol. Eng. 2012, 5, 48–55. [Google Scholar]
- Amoah, J.; Ishizue, N.; Ishizaki, M.; Yasuda, M.; Takahashi, K.; Ninomiya, K.; Yamada, R.; Kondo, A.; Ogino, C. Development and evaluation of consolidated bioprocessing yeast for ethanol production from ionic liquid-pretreated bagasse. Bioresour. Technol. 2017, 245, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.V.; de Barros Grassi, M.C.; Gallardo, J.C.M.; Pirolla, R.A.S.; Calderón, L.L.; de Carvalho-Netto, O.V.; Parreiras, L.S.; Camargo, E.L.O.; Drezza, A.L.; Missawa, S.K.; et al. Second-generation ethanol: The need is becoming a reality. Ind. Biotechnol. 2016, 12, 40–57. [Google Scholar] [CrossRef]
- Henriques, T.; Pereira, S.; Serafim, L.; Xavier, A. Two-stage aeration fermentation strategy to improve bioethanol production by Scheffersomyces stipitis. Fermentation 2018, 4, 97. [Google Scholar] [CrossRef]
- Pereira, S.R.; Ivanuša, Š.; Evtuguin, D.V.; Serafim, L.S.; Xavier, A.M.R.B. Biological treatment of eucalypt spent sulphite liquors: A way to boost the production of second generation bioethanol. Bioresour. Technol. 2012, 103, 131–135. [Google Scholar] [CrossRef]
- Pereira, S.R.; Sànchez i Nogué, V.; Frazão, C.J.R.; Serafim, L.S.; Gorwa-Grauslund, M.F.; Xavier, A.M.R.B. Adaptation of Scheffersomyces stipitis to hardwood spent sulfite liquor by evolutionary engineering. Biotechnol. Biofuels 2015, 8, 50. [Google Scholar] [CrossRef]
- Mendes, J.A.S.; Xavier, A.M.R.B.; Evtuguin, D.V.; Lopes, L.P.C. Integrated utilization of grape skins from white grape pomaces. Ind. Crops Prod. 2013, 49, 286–291. [Google Scholar] [CrossRef]
- Jiang, C.-X.; He, Y.-C.; Chong, G.-G.; Di, J.-H.; Tang, Y.-J.; Ma, C.-L. Enzymatic in situ saccharification of sugarcane bagasse pretreated with low loading of alkalic salts Na2SO3/Na3PO4 by autoclaving. J. Biotechnol. 2017, 259, 73–82. [Google Scholar] [CrossRef]
- Chandel, A.K.; Antunes, F.A.; Silva, M.B.; da Silva, S. Unraveling the structure of sugarcane bagasse after soaking in concentrated aqueous ammonia (SCAA) and ethanol production by Scheffersomyces (Pichia) stipitis. Biotechnol. Biofuels 2013, 6, 102. [Google Scholar] [CrossRef]
- Fernandes, M.C.; Ferro, M.D.; Paulino, A.F.C.; Chaves, H.T.; Evtuguin, D.V.; Xavier, A.M.R.B. Comparative study on hydrolysis and bioethanol production from cardoon and rockrose pretreated by dilute acid hydrolysis. Ind. Crops Prod. 2018, 111, 633–641. [Google Scholar] [CrossRef]
- Chen, Y. Development and application of co-culture for ethanol production by co-fermentation of glucose and xylose: A systematic review. J. Ind. Microbiol. Biotechnol. 2011, 38, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.W.; Baird, R.B.; Eaton, A.D. Standard Methods for Examination of Water and Wastewater, 23rd ed.; AWWA: City and County of Denve, CO, USA, 2017. [Google Scholar]
- Kang, Q.; Appels, L.; Tan, T.; Dewil, R. Bioethanol from lignocellulosic biomass: Current findings determine research priorities. Sci. World J. 2014, 2014, 298153. [Google Scholar] [CrossRef]
- Arévalo, C.; Freer, J.; Naulin, P.A.; Barrera, N.P.; Troncoso, E.; Araya, J.; Peña-Farfal, C.; del P Castillo, R. Study of the ultrastructure of Eucalyptus globulus wood substrates subjected to auto-hydrolysis and diluted acid hydrolysis pre-treatments and its influence on enzymatic hydrolysis. Bioenergy Res. 2017, 10, 714–727. [Google Scholar] [CrossRef]
- Snoek, T.; Verstrepen, K.J.; Voordeckers, K. How do yeast cells become tolerant to high ethanol concentrations? Curr. Genet. 2016, 62, 475–480. [Google Scholar] [CrossRef]
- Hahn-Hägerdal, B.; Karhumaa, K.; Fonseca, C.; Spencer-Martins, I.; Gorwa-Grauslund, M.F. Towards industrial pentose-fermenting yeast strains. Appl. Microbiol. Biotechnol. 2007, 74, 937–953. [Google Scholar] [CrossRef]
- Agbogbo, F.K.; Coward-Kelly, G.; Torry-Smith, M.; Wenger, K.S. Fermentation of glucose/xylose mixtures using Pichia stipitis. Process Biochem. 2006, 41, 2333–2336. [Google Scholar] [CrossRef]
- Agbogbo, F.K.; Coward-Kelly, G. Cellulosic ethanol production using the naturally occurring xylose-fermenting yeast, Pichia stipitis. Biotechnol. Lett. 2008, 30, 1515–1524. [Google Scholar] [CrossRef]
- Pereira, S.R.; Portugal-Nunes, D.J.; Evtuguin, D.V.; Serafim, L.S.; Xavier, A.M.R.B. Advances in ethanol production from hardwood spent sulphite liquors. Process Biochem. 2013, 48, 272–282. [Google Scholar] [CrossRef]
- Slininger, P.J.; Gorsich, S.W.; Liu, Z.L. Culture nutrition and physiology impact the inhibitor tolerance of the yeast Pichia stipitis NRRL Y-7124. Biotechnol. Bioeng. 2009, 102, 778–790. [Google Scholar] [CrossRef]
- Geng, A.; Wang, Z.; Lai, K.S.; Tan, M.W.Y. Bioenergy II: Comparison of laboratory and industrial Saccharomyces cerevisiae strains for their stress tolerance. Int. J. Chem. React. Eng. 2010, 8, A87. [Google Scholar] [CrossRef]
- Silva, J.P.A.; Mussatto, S.I.; Roberto, I.C.; Teixeira, J.A. Fermentation medium and oxygen transfer conditions that maximize the xylose conversion to ethanol by Pichia stipitis. Renew. Energy 2012, 37, 259–265. [Google Scholar] [CrossRef]
- Silva, D.D.V.; Dussán, K.J.; Hernández, V.; Silva, S.S.d.; Cardona, C.A.; Felipe, M.d.G.d.A. Effect of volumetric oxygen transfer coefficient (kLa) on ethanol production performance by Scheffersomyces stipitis on hemicellulosic sugarcane bagasse hydrolysate. Biochem. Eng. J. 2016, 112, 249–257. [Google Scholar] [CrossRef]
- Gutiérrez-Rivera, B.; Ortiz-Muñiz, B.; Gómez-Rodríguez, J.; Cárdenas-Cágal, A.; Domínguez González, J.M.; Aguilar-Uscanga, M.G. Bioethanol production from hydrolyzed sugarcane bagasse supplemented with molasses “B” in a mixed yeast culture. Renew. Energy 2015, 74, 399–405. [Google Scholar] [CrossRef]
- Papini, M.; Nookaew, I.; Uhlén, M.; Nielsen, J. Scheffersomyces stipitis: A comparative systems biology study with the Crabtree positive yeast Saccharomyces cerevisiae. Microb. Cell Fact. 2012, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Santosh, I.; Ashtavinayak, P.; Amol, D.; Sanjay, P. Enhanced bioethanol production from different sugarcane bagasse cultivars using co-culture of Saccharomyces cerevisiae and Scheffersomyces (Pichia) stipitis. J. Environ. Chem. Eng. 2017, 5, 2861–2868. [Google Scholar] [CrossRef]
- Sreemahadevan, S.; Singh, V.; Roychoudhury, P.K.; Ahammad, S.Z. Mathematical modeling, simulation and validation for co-fermentation of glucose and xylose by Saccharomyces cerevisiae and Scheffersomyces stipitis. Biomass Bioenergy 2018, 110, 17–24. [Google Scholar] [CrossRef]
- Beck, M.J.; Johnson, R.D.; Baker, C.S. Ethanol production from glucose/xylose mixes by incorporating microbes in selected fermentation schemes. Appl. Biochem. Biotechnol. 1990, 24, 415–424. [Google Scholar] [CrossRef]
- Zambare, V.P.; Bhalla, A.; Muthukumarappan, K.; Sani, R.K.; Christopher, L.P. Bioprocessing of agricultural residues to ethanol utilizing a cellulolytic extremophile. Extremophiles 2011, 15, 611–618. [Google Scholar] [CrossRef]
- Zambare, V.P.; Christopher, L.P. Optimization of enzymatic hydrolysis of corn stover for improved ethanol production. Energy Explor. Exploit. 2012, 30, 193–205. [Google Scholar] [CrossRef]
- Kelbert, M.; Romaní, A.; Coelho, E.; Pereira, F.B.; Teixeira, J.A.; Domingues, L. Simultaneous saccharification and fermentation of hydrothermal pretreated lignocellulosic biomass: Evaluation of process performance under multiple stress conditions. Bioenergy Res. 2016, 9, 750–762. [Google Scholar] [CrossRef][Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).