Abstract
This study examined the feasibility of using non-sterilized swine wastewater for lipid production by an isolated microalga, Scenedesmus rubescens. Different dilution ratios using municipal wastewater as a diluent were tested to determine the suitable levels of microalgal growth in the wastewaters, its nutrient removal, and its lipid production. The highest lipid productivity (8.37 mg/L/d) and NH4+ removal (76.49%) were achieved in swine wastewater that had been diluted to 30 times using municipal wastewater. Various bacteria coexisted in the wastewaters during the cultivation of S. rubescens. These results suggest the practical feasibility of a system to produce lipids from swine wastewater by using microalgae.
1. Introduction
Biodiesel is a critical need to reduce human dependency on fossil fuels and to decrease greenhouse gas effects. The use of microalgae to produce biodiesel achieves high lipid yields per unit area and high growth rates, and therefore, has the potential to completely replace fossil fuels [1,2,3,4,5]. In addition, this method does not compete with humans by using food plants or agricultural land. However, large-scale microalgal biomass production is not currently practical due to high production costs [6,7]. The production of 1 kg of biodiesel from microalgal biomass requires 3726 kg of water, 0.33 kg of nitrogen, and 0.71 kg of phosphate [8]. Water and nutrients are essential constituents of microalgal cultivation; these are available in agricultural wastewater, so its use has been suggested as an alternative feedstock for microalgal cultivation [3]. This approach has the dual advantages of treating wastewater and producing economically-valuable biomass.
Swine waste treatment has become increasingly necessary, as swine production has increased [9]. In Korea, more than 32 million tons of swine wastewater were generated in 2015 [10]. In general, swine wastewaters are rich in organic matter, nitrogen, and phosphorus [11]. However, microalgal growth in swine wastewaters is impeded by their high turbidity, high NH4+ concentration, and antagonistic microorganisms present in the media. A satisfactory level of microalgal biomass production has been attained using sterilized [12] or ultra-filtered [13] swine wastewater, but these approaches have limited applicability to large-scale cultivation because the pretreatment processes are expensive.
Dilution of swine wastewater is a straightforward and practical pretreatment to reduce its high turbidity and NH4+ concentration. Microalgal biomass production and nutrient removal are influenced by the degree of dilution [14]. However, there might be much controversy over the use of potable water, such as tap or deionized water as a diluent, and a cost-effective diluent is required for a large-scale cultivation system [15]. In this respect, raw municipal wastewater that contains relatively low levels of organic matter and nutrients may be an effective and inexpensive diluent. In addition, swine wastewater and municipal wastewater contain diverse bacteria that stimulate or inhibit microalgal growth, and dilution ratios may affect the relationships among them. Therefore, an investigation of microbial communities at different levels of dilution ratios can help to evaluate the feasibility of stable microalgal biomass production from swine wastewater diluted with municipal wastewater.
In this study, the feasibility of using non-sterilized swine wastewater for lipid production by isolated microalgae was explored. Different dilution ratios using raw municipal wastewater as a diluent were tested to determine the levels to which microalgal growth could adapt in the medium, and to quantify how the dilution ratios affect lipid production.
2. Materials and Methods
2.1. Isolation, Identification, and Cultivation of Microalgae
Raw municipal wastewater was collected from a municipal wastewater treatment plant in Pohang, Korea. Raw swine wastewater was collected from a swine wastewater treatment plant in Gyeongju, Korea (Table 1). Microalgae were not present in the raw swine wastewater, but they were identified in the raw municipal wastewater by using a confocal laser scanning microscope (LSM-5-Exitor, Carl Zeiss, Jena, Germany). The raw municipal wastewater was incubated in 250-mL Erlenmeyer flasks under a 12-h light period (70 µmol/m2/s light intensity) at 25 ± 1 °C for two weeks. Then samples of them were spread on BG-11 agar plates and incubated at 25 ± 1 °C under constant light (70 µmol/m2/s light intensity) until colonies were formed. Each colony was streaked on new BG-11 agar plates and incubated under the same condition, then each colony from the streaked plates was cultivated in BG-11 liquid medium. BG-11 medium contains the following chemicals per liter of distilled water: 1.5 g NaNO3, 0.04 g K2HPO4, 0.075 g MgSO4·7H2O, 0.02 g Na2CO3, 0.036 g CaCl2·2H2O, 0.006 g citric acid, 0.006 g ferric ammonium citrate, 0.001 g EDTA, and trace elements solution (1 mL) containing H3BO3 (2.86 g/L), MnCl2·4H2O (1.81 g/L), Na2MoO4·2H2O (0.39 g/L), ZnSO4·7H2O (0.22 g/L), Co(NO3)2·6H2O (49.4 mg/L), and CuSO4·5H2O (0.079 g/L).

Table 1.
Characteristics of swine wastewater and municipal wastewater.
Selected microalgal species were identified by DNA sequence analysis (Table 2). Their DNAs were extracted using the NucleoSpin soil kit (Macherey-Nagel, Duren, Germany). A polymerase chain reaction (PCR) of 18S rRNA was conducted: each PCR reagent contained Takara ExTaq DNA polymerase, 10× ExTaq buffer, dNTP mixture, DNA sample, forward primer, reverse primer, and sterilized distilled water. The forward primer used for PCR was Euk1 (5′-CTGGTTGATCCTGCCAG-3′), and the reverse was Euk516 (5′-ACCAGACTTGCCCTCC-3′). PCR conditions consisted of: (1) an initial denaturing step at 94 °C for 130 s; (2) 35 cycles of denaturing at 94 °C for 30 s, annealing at 56 °C for 45 s, and extension at 72 °C for 130 s; and (3) the last extension at 72 °C for 6 min [16]. The PCR samples were sequenced (Solgent, Co., Daejeon, South Korea), and ribosomal RNA genes were identified using the National Center for Biotechnology Information (NCBI) BLAST program to compare the sequences to references. Selected species were cultivated in the 250-mL Erlenmeyer flasks containing 150 mL sterilized BG-11 medium at 25 ± 1 °C, with agitation at 130 rpm, and constant light intensity of 100 µmol/m2/s for 12 days.
Table 2.
Isolation and identification of microalgae from municipal wastewater.
2.2. Design of Experiment
The green microalga Scenedesmus sp. are widely used in wastewater treatment because they accumulate lipid and self-flocculate for harvesting [17,18]. Therefore, S. rubescens was selected for cultivation in a mixture of swine wastewater and municipal wastewater. To determine the effect of nutrient concentrations on the growth of S. rubescens, the swine wastewater was diluted with municipal wastewater by a factor of 5, 10, 20, and 30, respectively (Table 1). Media are denoted as Nx, where N is the dilution factor. Then each mixed wastewater was filtered through 1.2-µm glass microfiber filters (Whatman, Buckinghamshire, UK). S. rubescens was cultured in batch mode without pH control in all media in Erlenmeyer flasks (500-mL working volume) sealed with sterile gauze stoppers. Sixteen percent (v/v) inoculum cultivated in sterilized BG-11 medium was used for batch cultures when the inoculum was in the mid-exponential phase of cell growth. The experiments were carried out in duplicate at 25 ± 1 °C with 130 rpm agitation, 100 µmol/m2/s light intensity, and photoperiod of 12 h.
2.3. Physioco-Chemical Analysis
During cultivation, biomass concentration was determined periodically by the weight difference of GF/C filters before and after filtering and drying (105 °C, 24 h) samples of known volume. For analysis of chemical oxygen demand (COD), total nitrogen (TN), total phosphorus (TP), and NH4+, microalgal suspensions taken from each cultivation were filtered through a 0.45-µm pore-size filter, then the filtered samples were processed using the method of APHA (American Public Health Association, 1998) [19]. The pH of culture media was determined using pH measuring equipment (Orion 5-star, Thermo Scientific, USA). Lipids were extracted from S. rubescens by using the modified Bligh and Dyer method [20].
The cells were centrifuged (2000 rpm for 15 min), washed with distilled water, then lyophilized. The lyophilized biomass was resuspended in 1.6 mL of distilled water. A total of 6 mL of chloroform (HPLC grade, J.T Baker) and methanol (HPLC grade, Samchun) mixture (1:2 of v/v) and 0.8 mL of glass beads were added to the suspension. The suspension was vortexed for 1 min, then 2 mL of chloroform was added before the first incubation of suspension at room temperature for 5 min. A total of 2 mL of distilled water was added before the second incubation at room temperature for 5 min. The suspension was centrifuged at 2000 rpm for 5 min, then the low phase was evaporated using a nitrogen evaporator (Organomation Associates Inc., Berlin, MA, USA) to weigh the lipids. Fatty acid methyl esters (FAMEs) were determined with transesterified lipids by using a gas chromatograph (6890N, Agilent, Santa Clara, CA, USA) equipped with an INNOWAX capillary column (Agilent, Santa Clara, CA, USA). The column temperature was controlled by reference to previously reported procedures [21]. The FAMEs for biodiesel properties were evaluated using the Pinzi model [22].
2.4. Microbial Community Analysis
Bacterial communities were identified using denaturing gradient gel electrophoresis (DGGE). DNA was extracted using the same method as described above. PCR amplification was conducted using primer 338F (5′-ACTCCTACGGGAGGCAG-3′) and primer 805R (5′-GACTACCAGGGTATCTAATCC-3′). PCR conditions consisted of: (1) an initial denaturing step at 95 °C for 300 s; (2) 30 cycles of denaturing at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 30 s; and (3) the last cycle extension at 72 °C for 10 min [23]. DGGE was conducted using a DCODE system (BioRAD). The PCR product (25 μL) was loaded onto 8% polyacrylamide gel with a 30–70% denaturing gradient. The DGGE condition was 17-h running time at 100 V and 60 °C in 0.5× TAE buffer. After electrophoresis, polyacrylamide gel was stained with ethidium bromide and analyzed under trans UV using Gel DOC XR (BioRAD, Hercules, CA, USA). Visible bands in the gel were excised and DNA in the excised gel was extracted in 40 μL of deionized water. The eluted DNA was amplified using the same primers (without GC clamp) and conditions as described above. The PCR samples were sequenced (Solgent, Co., Daejeon, South Korea), and ribosomal RNA genes were identified using the NCBI BLAST program to compare the sequences to references.
3. Results and Discussion
3.1. Biomass Production and Nutrient Removal
Biomass concentration increased in all cultures until day 3 (Figure 1a). However, the growth profiles in 5× and 10× cultures were different from those in 20× and 30× cultures. In 5× and 10× cultures, the biomass achieved its maximum level on day 3 and on day 1, respectively, then decreased to a stationary level. In 20× and 30× cultures, the biomass continually increased and achieved its peak level on day 14. The highest biomass productivity was attained in 30× culture (55.0 mg/L/d), followed by 20× (28.6 mg/L/d), 5× (18.6 mg/L/d), and 10× (10.7 mg/L/d) cultures (Table 3). To check the microalgal growth, changes in the color of media were monitored during cultivation. At the beginning of the experiment, all mixed media were brown. UV400nm represents the level of color darkness regarding wastewater [24]; raw swine wastewater was darker than raw municipal wastewater, so brown color decreased in the order of 5× medium, followed by 10×, 20×, and 30× (Table 1). As time elapsed, the colors of 5× and 10× media remained brown during the 14 days of cultivation, whereas those of the 20× and 30× media gradually turned green; this result indicates the growth of microalgal populations in 20× and 30× cultures.
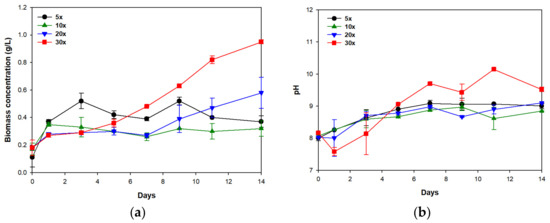
Figure 1.
(a) Biomass production and (b) pH variation during the cultivation of S. rubescens in swine wastewater differently diluted with municipal wastewater. Error bars indicate standard deviations of duplicate tests.
Table 3.
Biomass and lipid productivities of S. rubescens in swine wastewater differently diluted with municipal wastewater.
The pH in the media also increased; this change represents microalgal growth, which entails the production of alkalinity [25]. After 14 days of cultivation, the pH was highest in the 30× culture (Figure 1b); this result seemed to be related to the predominant microalgal growth in the 30× culture. Distinct growth of microalgal populations in the 20× culture was identified by the gradual change in the color of the medium, but the increase in the pH value was smaller in the 20× culture than in the 30× culture; the difference might be attributed to relatively lower microalgal growth and biomass concentrations in the 20× medium than in the 30× medium (Figure 1a).
The pattern of organic carbon reduction was similar in all cultures (Figure 2a). COD decreased by more than 70% within the first three days, then remained relatively stationary. In all cultures, organic carbon removal did not differ much between day 3 and day 14 (Table 4). There were slight fluctuations in COD removal between day 3 and day 14; the decrease in COD removal on day 14 might have been associated with the production of organic matter, such as glycolic acid by microalgae [26]. NH4+ removal differed among cultures. In the 5× and 10× cultures, it decreased during the first three days, then almost remained constant. In the 20× and 30× cultures, NH4+ decreased continuously (Figure 2b; Table 4). In general, both microalgae and heterotrophic bacteria can compete for organic carbons as a carbon source in the media, but microalgae grow more slowly than heterotrophic bacteria [27,28]. The decrease in NH4+ concentration is mainly attributed to the absorption by microalgae or bacteria. However, NH4+ of 200–500 mg/L can inhibit microalgal growth in media during axenic culture [29,30]. Therefore, the initial concentration of NH4+ in 5× and 10× media seemed to be inhibitory or toxic to microalgal growth, so in cultures, heterotrophic bacteria may mainly contribute to the removal of NH4+ and organic carbon within the first three days; afterwards, further heterotrophic growth of bacteria or microalga after day 3 may have been suppressed because the residual organic carbons may be recalcitrant, and therefore, unusable. In addition, relatively limited light penetration due to high turbidity in 5× and 10× media might suppress the photosynthetic growth of microalga. In contrast, in the 20× and 30× cultures, microalga and heterotrophic bacteria may both contribute to the removal of NH4+ and of organic carbon within the first three days, then photosynthetic growth of microalga after day 3 would have resulted in further predominant NH4+ removal after day 3 (Table 4). The highest NH4+ removal efficiency was achieved in the 30× culture (76.49%), followed by 20× (52.87%), 5× (28.41%), and 10× (26.22%) cultures (Table 4). The trend of NH4+ removal efficiency was similar to that of biomass productivity; this result indicates that the 30× medium contains appropriate concentrations of organic matter and nutrients to sustain microalgal growth in swine wastewater mixed with municipal wastewater.
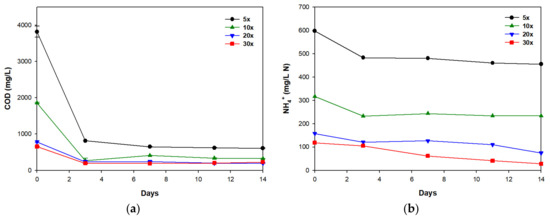
Figure 2.
(a) Chemical oxygen demand (COD) and (b) NH4+ removals during the cultivation of S. rubescens in swine wastewater differently diluted with municipal wastewater. Error bars indicate standard deviations of duplicate tests.
Table 4.
COD and NH4+ removals in swine wastewater differently diluted with municipal wastewater.
3.2. Lipid and FAMEs Profile
The highest lipid content was obtained in the 20× culture (16.39%), followed by the 30× (15.06%), 10× (4.10%), and 5× (2.72%) cultures; lipid contents in both the 5× and 10× cultures were significantly lower than those in the 20× and 30× cultures. This result might be due to the higher concentration of NH4+ in 5× and 10× media than in 20× and 30× media. In general, lipid accumulation of microalgae occurs under nitrogen-deficient conditions [31]. The highest lipid productivity was achieved in the 30× culture (8.37 mg/L/d), followed by the 20× (4.44 mg/L/d), 5× (0.49 mg/L/d), and 10× (0.44 mg/L/d) cultures (Table 3). Although lipid content was higher in the 20× culture than in the 30× culture, lipid productivity was significantly higher in the 30× culture (8.37 mg/L/d) than in the 20× culture (4.44 mg/L/d) due to the high biomass productivity in the 30× culture. The trend of lipid productivity was similar to that of biomass productivity in all cultures; this result signifies that the difference in biomass productivity was greater than the difference in lipid content.
Fatty acid composition of the biomass on day 14 was determined to check its suitability for use in biodiesel production (Figure 3). In all cultures, the main fatty acids had chain lengths from C16 to C18. However, in the 5× and 10× cultures, saturated (C16:0) and mono-unsaturated (C16:1 and C18:1) fatty acids dominated the fatty acid composition, whereas, in 20× and 30× cultures, the proportion of poly-unsaturated fatty acids such as C18:2 and C18:3 increased. This difference might be related to the relatively stable microalgal growth in 20× and 30× cultures in that fatty acid compositions in 20× and 30× cultures were more similar to those in axenic cultures of Scenedesmus spp. [26,32] than were the compositions in 5× and 10× cultures.
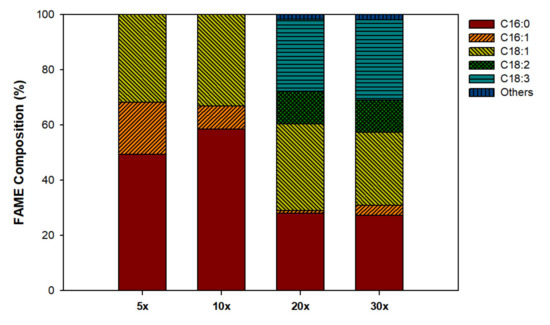
Figure 3.
Fatty acid profiles during the cultivation of S. rubescens in swine wastewater differently diluted with municipal wastewater.
The quality of biodiesel mainly depends on fatty acid composition in triacylglycerol. Therefore, the lipids from microalgal biomass in 20× and 30× cultures were evaluated based on Pinzi’s model to check their feasibility as a biodiesel feedstock [22,33]. The lipids from microalgal biomass met cetane number, kinematic viscosity, and flash point requirements of the European biodiesel standard (EN 14214) (Table 5). However, the lower calorific value (LCV) was lower in the microalgal lipids than in typical petroleum diesel; this biodiesel from microalgal biomass might have lower energy density than does conventional diesel.
Table 5.
The standard of biodiesel based on EN14214 and biodiesel quality of S. rubescens cultivated in swine wastewater diluted with municipal wastewater.
3.3. Microbial Community Structure
The relative abundance of bacterial communities was investigated by DGGE analysis (Figure 4). Ten bacterial strains were identified. Arcobacter cloacae and Acidovorax sp. were observed on day 11 in all culture conditions. However, Pseudomonas fragi was detected during 11 days in the 5× culture, while it disappeared on day 11 in the 10×, 20×, and 30× cultures. Flavobacterium sp. was identified only in 5× and 10× cultures. Members of these genera are denitrifiers [35,36,37,38]. Bacteria can suppress microalgal growth by ways such as competition for nutrients, excretion of algicidal matter, and degradation of microalgal cells through direct contact [39]. Pseudomonas sp. and Flavobacterium sp. may contaminate mass cultures of microalgae in open systems and can decompose microalgae [40]. Porphyrobacter sp. was observed on day 11 in all culture conditions. Members of this genus are associated with the biosynthesis and transmission of vitamin B12, which can promote microalgal growth [41]. Therefore, this strain seems to contribute to microalgal growth. Erythromicrobium sp., Aquimonas voraii, and Roseomonas sp. were only detected in the 30× culture. Erythromicrobium sp. is an aerobic phototrophic bacterium; it can have positive interactions between plants and microalgae when they are co-cultured in wastewater [42]. Aquimonas voraii has an extracellular phosphatase activity, which can enhance the availability of phosphorus to algae [43]. Roseomonas sp. has the ability to mineralize organic matter when co-cultured with algae [44]. Therefore, these strains may contribute to the predominant microalgal growth in the 30× culture.
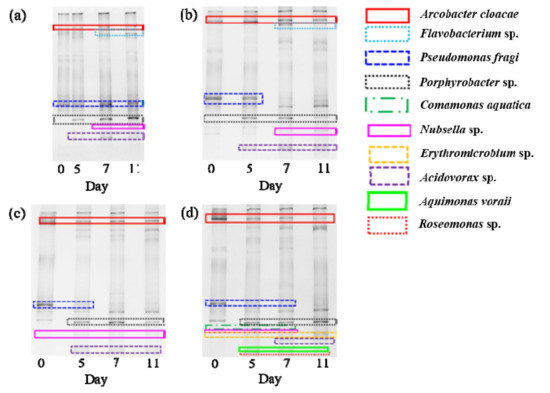
Figure 4.
Denaturing gradient gel electrophoresis (DGGE) profiles of bacterial communities during the cultivation of S. rubescens. in swine wastewater differently diluted with municipal wastewater: (a) 5×, (b) 10×, (c) 20×, (d) 30×.
Microalgal resistance to bacterial contamination is one of the most important factors when selecting species for biodiesel production from wastewater. The bacterial community structure indicates that coexisting bacteria influenced microalgal growth and nutrient removal in the wastewater. To enhance microalgal growth and nutrient removal in this wastewater, co-cultivation of microalgal biomass and an artificial consortium composed of isolated bacteria that promote microalgal growth might be beneficial to microalgal-bacterial systems.
4. Conclusions
This study demonstrated the suitable levels of S. rubescens growth in non-sterilized swine wastewater with municipal wastewater as a diluent. Different levels of dilution ratios resulted in different lipid production, NH4+ removal, and bacterial communities in the media during microalgal cultivation. The highest lipid productivity (8.37 mg/L/d) and NH4+ removal (76.49%) were achieved in swine wastewater that had been diluted to 30 times using municipal wastewater. Although we illuminated that high-strength swine wastewater at appropriate dilutions with municipal wastewater can be a suitable medium for S. rubescens growth, further research should consider a reduction in the amount of added municipal wastewater to strengthen the attractiveness of this process for large-scale applications.
Author Contributions
J.M.P. and H.U.C. conceived the experiments and J.C.U. performed the experiments. J.C.U. and H.U.C. analyzed the data. J.C.U., H.U.C., and J.M.P. wrote the manuscript. Y.M.K. reviewed the manuscript. H.U.C. and J.M.P. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the development fund foundation, Gyeongsang National University, 2019. This work was also supported by the Korea Institute of Energy Technology Evaluation and planning (KETEP) funded by the Ministry of Trade, Industry and Energy of the Republic of Korea (No. 20194030202330).
Conflicts of Interest
The authors declare no conflict of interest.
References
- An, M.; Gao, L.; Zhao, W.; Chen, W.; Li, M. Effects of nitrogen forms and supply mode on lipid production of microalga Scenedesmus obliquus. Energies 2020, 13, 697. [Google Scholar] [CrossRef]
- Kim, D.G.; Hong, C.-H.; Choi, Y.-E. Efficient algal biodiesel production with an optimal harvest condition obtained via response surface methodology. Bioresour. Technol. Rep. 2019, 7, 100213. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Ling, T.C.; Show, P.L.; Juan, J.C.; Chang, J.-S.; Lee, D.-J. Cultivation in wastewaters for energy: A microalgae platform. Appl. Energy 2016, 179, 609–625. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Fu, D.; Chu, H.; Wang, Z.; Qi, H. Biofuel production from microalgae: A review. Environ. Chem. Lett. 2020, 18, 285–297. [Google Scholar] [CrossRef]
- Bodenes, P.; Wang, H.-Y.; Lee, T.-H.; Chen, H.-Y.; Wang, C.-Y. Microfluidic techniques for enhancing biofuel and biorefinery industry based on microalgae. Biotechnol. Biofuels 2019, 12, 33. [Google Scholar] [CrossRef]
- Kim, G.-Y.; Roh, K.; Han, J.-I. The use of bicarbonate for microalgae cultivation and its carbon footprint analysis. Green Chem. 2019, 21, 5053–5062. [Google Scholar] [CrossRef]
- Yang, J.; Xu, M.; Zhang, X.; Hu, Q.; Sommerfeld, M.; Chen, Y. Life-cycle analysis on biodiesel production from microalgae: Water footprint and nutrients balance. Bioresour. Technol. 2011, 102, 159–165. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, L.; Riddicka, B.A.; Li, R.; Able, J.R.; Boakye-Boaten, N.A.; Shahbazi, A. Sustainable production of algal biomass and biofuels using swine wastewater in North Carolina, US. Sustainability 2016, 8, 477. [Google Scholar] [CrossRef]
- Kwon, G.; Nam, J.-H.; Kim, D.-M.; Song, C.; Jahng, D. Growth and nutrient removal of Chlorella vulgaris in ammonia-reduced raw and anaerobically-digested piggery wastewaters. Environ. Eng. Res. 2020, 25, 135–146. [Google Scholar] [CrossRef]
- Zou, G.; Liu, Y.; Zhang, Q.; Zhou, T.; Xiang, S.; Gu, Z.; Huang, Q.; Yan, H.; Zheng, H.; Wu, X.; et al. Cultivation of Chlorella vulgaris in a light-receiving-plate (LRP)-enhanced raceway pond for ammonium and phosphorus removal from pretreated pig urine. Energies 2020, 13, 1644. [Google Scholar] [CrossRef]
- Kuo, C.-M.; Chen, T.-Y.; Lin, T.-H.; Kao, C.-Y.; Lai, J.-T.; Chang, J.-S.; Lin, C.-S. Cultivation of Chlorella sp. GD using piggery wastewater for biomass and lipid production. Bioresour. Technol. 2015, 194, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Erol, H.B.U.; Menegazzo, M.L.; Sanderfur, H.; Gottberg, E.; Vaden, J.; Asgharpour, M.; Hestekin, C.N.; Hestekin, J.A. Porphyridium cruentum grown in ultra-filtered swine wastewater and its effects on microalgae growth productivity and fatty acid composition. Energies 2020, 13, 3194. [Google Scholar] [CrossRef]
- Franchino, M.; Tigini, V.; Varese, G.C.; Sartor, R.M.; Bona, F. Microalgae treatment removes nutrients and reduces ecotoxicity of diluted piggery digestate. Sci. Total Environ. 2016, 569, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Cheng, J.J.; Cobb, K.; Zhou, C.; Zhou, N.; Addy, M.; Chen, P.; Yan, X.; Ruan, R. Tribonema sp. and Chlorella zofingiensis co-culture to treat swine wastewater diluted with fishery wastewater to facilitate harvest. Bioresour. Technol. 2020, 297, 122516. [Google Scholar] [CrossRef] [PubMed]
- Eland, L.E.; Davenport, R.; Mota, C.R. Evaluation of DNA extraction methods for freshwater eukaryotic microalgae. Water Res. 2012, 46, 5355–5364. [Google Scholar] [CrossRef]
- Lv, J.; Guo, B.; Feng, J.; Liu, Q.; Nan, F.; Liu, X.; Xie, S. Integration of wastewater treatment and flocculation for harvesting biomass for lipid production by a newly isolated self-flocculating microalga Scenedesmus rubescens SX. J. Clean Prod. 2019, 240, 118211. [Google Scholar] [CrossRef]
- Abinandan, S.; Shanthankumar, S. Challenges and opportunities in application of microalgae (Chlorophyta) for wastewater treatment: A review. Renew. Sustain. Energy Rev. 2015, 52, 123–132. [Google Scholar] [CrossRef]
- Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Bourque, S.D.; Titorenko, V.I. A quantitative assessment of the yeast lipidome using electrospray ionization mass spectrometry. J. Vis. Exp. 2009, 30, 1513. [Google Scholar] [CrossRef]
- Xu, X.; Kim, J.Y.; Cho, H.U.; Park, H.R.; Park, J.M. Bioconversion of volatile fatty acids from macroalgae fermentation into microbial lipids by oleaginous yeast. Chem. Eng. J. 2015, 264, 735–743. [Google Scholar] [CrossRef]
- Pinzi, S.; Leiva, D.; Arzamendi, G.; Gandia, L.M.; Dorado, M.P. Multiple response optimization of vegetable oils fatty acid composition to improve biodiesel physical properties. Bioreour. Technol. 2011, 102, 7280–7288. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Hwang, K.; Shin, S.G.; Lee, S.; Hwang, S. Effect of high temperature on bacterial community dynamics in anaerobic acidogenesis using mesophilic sludge inoculum. Bioreour. Technol. 2010, 101, S17–S22. [Google Scholar] [CrossRef] [PubMed]
- Hatt, J.W.; Germain, E.; Judd, S.J. Powered activated carbon-microfiltration for waste-water reuse. Sep. Sci. Technol. 2013, 48, 690–698. [Google Scholar] [CrossRef]
- Yeh, K.L.; Chen, C.Y.; Chang, J.S. PH-stat photoheterotrophic cultivation of indigenous Chlorella vulgaris ESP-31 for biomass and lipid production using acetic acid as the carbon source. Biochem. Eng. J. 2012, 64, 1–7. [Google Scholar] [CrossRef]
- Shin, D.Y.; Cho, H.U.; Utomo, J.C.; Choi, Y.N.; Xu, X.; Park, J.M. Biodiesel production from Scenedesmus bijuga grown in anaerobically digested food wastewater effluent. Bioresour. Technol. 2015, 184, 215–221. [Google Scholar] [CrossRef]
- Cho, H.U.; Kim, Y.M.; Park, J.M. Enhanced microalgal biomass and lipid production from a consortium of indigenous microalgae and bacteria present in municipal wastewater under gradually mixotrophic culture conditions. Bioresour. Technol. 2017, 228, 290–297. [Google Scholar] [CrossRef]
- Fenchel, T. Intrinsic rate of natural increase: The relationship with body size. Oecologia 1974, 14, 317–326. [Google Scholar] [CrossRef]
- He, P.J.; Mao, B.; Shen, C.M.; Shao, L.M.; Lee, D.J.; Chang, J.S. Cultivation of Chlorella vulgaris on wastewater containing high levels of ammonia for biodiesel production. Bioresour. Technol. 2013, 129, 177–181. [Google Scholar] [CrossRef]
- Park, J.; Jin, H.-F.; Lim, B.-R.; Park, K.-Y.; Lee, K. Ammonia removal from anaerobic digestion effluent of livestock waste using green alga Scenedesmus sp. Bioresour. Technol. 2010, 101, 8649–8657. [Google Scholar] [CrossRef]
- Cheng, P.; Wang, Y.; Osei-Wusu, D.; Wang, Y.; Liu, T. Development of nitrogen supply strategy for Scenedesmus rubescens attached cultivation toward growth and lipid accumulation. Bioprocess. Biosyst. Eng. 2018, 41, 435–442. [Google Scholar] [CrossRef]
- Ye, S.; Gao, L.; Zhao, J.; An, M.; Wu, H.; Li, M. Simultaneous wastewater treatment and lipid production by Scenedesmus sp. HXY2. Bioreour. Technol. 2020, 302, 122903. [Google Scholar] [CrossRef] [PubMed]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process. Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]
- Suh, H.K.; Lee, C.S. A review on atomization and exhaust emissions of a biodiesel-fueled compression ignition engine. Renew. Sustain. Energy Rev. 2016, 58, 1601–1620. [Google Scholar] [CrossRef]
- Levican, A.; Collado, L.; Figueras, M.J. Arcobacter cloacae sp. nov. and Arcobacter suis sp. nov., two new species isolated from food and sewage. Syst. Appl. Microbiol. 2013, 36, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Heylen, K.; Lebbe, L.; Vos, P.D. Acidovorax caeni sp. nov., a denitrifying species with genetically diverse isolates from activated sludge. Int. J. Syst. Evol. Microbiol. 2008, 58, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Guo, L.; Shao, M.; Hu, F.; Wang, G.; Zhao, Y.; Gao, M.; Jin, C.; She, Z. Heterotrophic denitrification strategy for marine recirculating aquaculture wastewater treatment using mariculture solid wastes fermentation liquid as carbon source: Optimization of COD/NO3− -N ratio and hydraulic retention time. Bioreour. Technol. 2020, 304, 122982. [Google Scholar] [CrossRef]
- Ryu, S.H.; Park, M.; Jeon, Y.; Lee, J.R.; Park, W.; Jeon, C.O. Flavobacterium filum sp. nov., isolated from a wastewater treatment plant in Korea. Int. J. Syst. Evol. Microbiol. 2007, 57, 2026–2030. [Google Scholar] [CrossRef]
- Han, J.; Zhang, L.; Wang, S.; Yang, G.; Zhao, L.; Pan, K. Co-culturing bacteria and microalgae in organic carbon containing medium. J. Biol. Res. 2016, 23, 8. [Google Scholar] [CrossRef]
- Afi, L.; Metzger, P.; Largeau, C.; Connan, J.; Berkaloff, C.; Rousseau, B. Bacterial degradation of green microalgae: Incubation of Chlorella emersonii and Chlorella vulgaris with Pseudomonas oleovorans and Flavobacterium aquatile. Org. Geochem. 1996, 25, 117–130. [Google Scholar] [CrossRef]
- Krohn-Molt, I.; Alawi, M.; Forstner, K.U.; Wiegandt, A.; Burkhardt, L.; Indenbirken, D.; Thieb, M.; Grundhoff, A.; Kehr, J.; Tholey, A.; et al. Insights into microalga and bacteria interactions of selected phycosphere biofilms using metagenomic, transcriptomic, and proteomic approaches. Front. Microbiol. 2017, 8, 1941. [Google Scholar] [CrossRef]
- Xinjie, W.; Xin, N.; Qilu, C.; Ligen, X.; Yuhua, Z.; Qifa, Z. Vetiver and Dictyosphaerium sp. co-culture for the removal of nutrients and ecological inactivation of pathogens in swine wastewater. J. Adv. Res. 2019, 20, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Murinda, S.E.; Murry, M.A.; Schwartz, G.; Lundquist, T. Microbial community structures in high rate algae ponds for bioconversion of agricultural wastes from livestock industry for feed production. Sci. Total Environ. 2017, 580, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, Y.; Yang, T.; He, Z.; Yan, Q. Size-fractioned aggregates within phycosphere define functional bacterial communities related Microcystis aeruginosa and Euglena sanguinea blooms. Aquat. Ecol. 2020, 54, 609–623. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).