Fungi-Based Microbial Fuel Cells


Abstract
1. Introduction
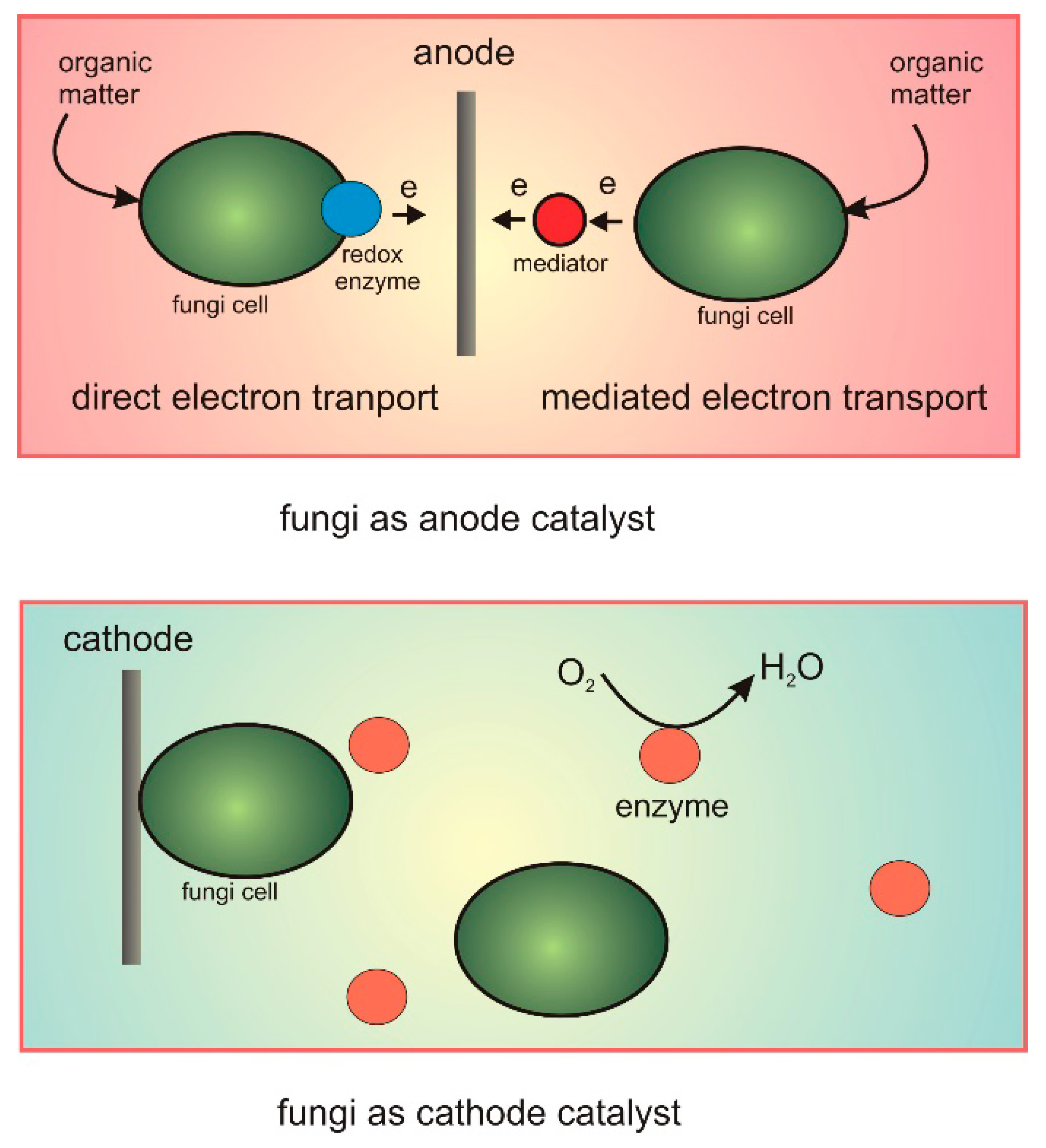
2. Fungi as Biocatalysts in the Anode of MFCs
2.1. Saccharomyces cerevisiae
2.2. Candida sp.
2.2.1. Candida melibiosica
2.2.2. Candida sp. IR11
2.3. Arxula adeninivorans
2.4. Hansenula anomala
2.5. Other Species
3. Fungi Used as a Cathode Catalyst
3.1. Trametes versicolor (Coriolus versicolor)
3.2. Ganoderma lucidum
3.3. Galactomyces reessii
3.4. Other Species
4. Summary
5. Challenges and Perspectives for Fungi-Based MFCs
Author Contributions
Funding
Conflicts of Interest
References
- Logan, B.E. Microbial Fuel Cells; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008. [Google Scholar]
- Franks, A.E.; Nevin, K.P. Microbial Fuel Cells, a Current Review. Energies 2010, 3, 899–919. [Google Scholar] [CrossRef]
- Toczyłowska-Mamińska, R.; Szymona, K.; Kloch, M. Bioelectricity production from wood hydrothermal treatment wastewater. Enhanced power generation in MFC fed mixed wastewaters. Sci. Total Environ. 2018, 634, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Fredrikson, J.K.; Romine, M.F.; Beliaev, A.S.; Auchtung, J.M.; Driscoll, M.E.; Gardner, T.S.; Nealson, K.H.; Osterman, A.L.; Pinchuk, G.; Reed, J.L. Towards environmental system biology of Shewanella. Nat. Rev. Microbiol. 2008, 6, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Ueki, T.; Zhang, T.; Malvankar, N.S.; Shrestha, P.M.; Flanagan, K.A.; Aklujkar, M.; Butler, J.E.; Giloteaux, L.; Rotaru, E.H.; et al. Geobacter: The microbe electric’s physiology, ecology, and practical application. Adv. Microb. Physiol. 2011, 59, 1–100. [Google Scholar] [CrossRef]
- Sanchez, D.V.P.; Jacobs, D.; Gregory, K.; Huang, J.; Hu, Y.; Vidic, R.; Yun, M. Changes in carbon electrode morphology affect microbial fuel cell performance with Shewanella oneidensis MR-1. Energies 2015, 8, 1817–1829. [Google Scholar] [CrossRef]
- Ali, J.; Sohail, A.; Wang, L.; Haider, M.R.; Mulk, S.; Pan, G. Electro-microbiology as a promising approach towards renewable energy and environmental sustainability. Energies 2015, 8, 1817–1829. [Google Scholar] [CrossRef]
- Toczyłowska-Mamińska, R.; Szymona, K.; Król, P.; Gliniewicz, K.; Pielech-Przybylska, K.; Kloch, M.; Logan, B.E. Evolving microbial communities in cellulose-fed microbial fuel cells. Energies 2018, 11, 124. [Google Scholar] [CrossRef]
- Potter, M.C. Electrical effects accompanying the decomposition of organic compounds. Proc. R. Soc. Lond. B Biol. Sci. 1911, 84, 260–276. [Google Scholar] [CrossRef]
- Sayed, E.T.; Abdelkareem, M.A. Yeast as a biocatalyst in microbial fuel cell. In Old Yeast—New Questions; Available online: https://www.intechopen.com/books/old-yeasts-new-questions/yeast-as-a-biocatalyst-in-microbial-fuel-cell (accessed on 19 October 2018).
- Prasad, D.; Arun, S.; Murugesan, M.; Padmanaban, S.; Satyanarayanan, R.S.; Berchmans, S.; Yegnaraman, V. Direct electron transfer with yeast cells and construction of a mediatorless microbial fuel cell. Biosens. Bioelectron. 2007, 22, 2604–2610. [Google Scholar] [CrossRef] [PubMed]
- Ducommun, R.; Favre, M.-F.; Carrard, D.; Fischer, F. Outward electron transfer by Saccharomyces cerevisiae monitored with a bi-cathodic microbial fuel cell-type activity sensor. Yeast 2010, 27, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.; Klar, J.; Applegarth, S. Optimizing biofuel cell performance using a targeted mixed mediator combination. Electroanalysis 2006, 18, 2001–2007. [Google Scholar] [CrossRef]
- Martínez, A.T.; Speranza, M.; Ruiz-Dueñas, F.J.; Ferreira, P.; Camarero, S.; Guillén, F.; Martínez, M.J.; Gutiérrez Suárez, A.; Río Andrade, J.C.D. Biodegradation of lignocellulosics: Microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Int. Microbiol. 2005, 8, 195–204. [Google Scholar] [PubMed]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Rahmanpour, R. Pathways for degradation of lignin in bacteria and fungi. Nat. Prod. Rep. 2011, 28, 1883–1896. [Google Scholar] [CrossRef] [PubMed]
- Wesenberg, D.; Kyriakides, I.; Agathos, S.N. White-rot fungi and their enzymes for the treatment of industrial dyes effluents. Biotechnol. Adv. 2003, 22, 161–187. [Google Scholar] [CrossRef] [PubMed]
- Leonowicz, A.; Cho, N.; Luterek, J.; Wilkolazka, A.; Wojtas-Wasilewska, M.; Matuszewska, A.; Hofrichter, M.; Wesenberg, D.; Rogalski, J. Fungal laccase: Properties and activity on lignin. J. Basic Microbiol. 2001, 41, 185–227. [Google Scholar] [CrossRef]
- Schaetzle, O.; Barriere, F.; Schroder, U. An improved microbial fuel cell with laccase as oxygen reduction catalyst. Energy Environ. Sci. 2009, 2, 96–99. [Google Scholar] [CrossRef]
- Sadhasivam, S.; Savitha, S.; Swaminathan, K.; Lin, F.H. Production, purification and characterization of mid-redox potential laccase from a newly isolated Trichoderma harzianum WL1. Process. Biochem. 2008, 43, 736–742. [Google Scholar] [CrossRef]
- Gallaway, J.; Wheeldon, I.; Rincon, R.; Atanassov, P.; Banta, S.; Barton, S.C. Oxygen-reducing enzymes cathodes produced from SLAC, a small laccase from Streptomyces coelicolor. Biosens. Bioelectron. 2008, 23, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Kipf, E.; Sané, S.; Morse, D.; Messinger, T.; Zengerle, R.; Kerzenmacher, S. An air-breathing enzymatic cathode with extended life time by continuous laccase supply. Bioresour. Technol. 2018, 264, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.H.; Shah, A.A.; Walsh, F.C. Recent progress and continuing challenges in bio-fuel cells, Part II: Microbial. Biosens. Bioelectron. 2010, 26, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Karnicka, K.; Miecznikowski, K.; Kowalewska, B.; Skunik, M.; Opallo, M.; Rogalski, J.; Schuhmann, W.; Kulesza, P.J. ABTS-modified multiwalled carbon nanotubes as an effective mediating system for bioelectrocatalytic reduction of oxygen. Anal. Chem. 2008, 80, 7643–7648. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.; Reading, N.S.; Aust, S.D. Relative stability of recombinant versus native peroxidases from Phanerochaete chrysosporium. Arch. Biochem. Biophys. 1999, 365, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Glenn, J.K.; Gold, M.H. Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-degrading basidiomycete, Phanerochaete chrysosporium. Arch. Biochem. Biophys. 1985, 242, 329–341. [Google Scholar] [CrossRef]
- Glenn, K.K.; Akileswaran, L.; Godls, M.H. Mn(II) oxidation is principal function of the extracellular Mn-peroxidase from Phanerochaete chrysosporium. Arch. Biochem. Biophys. 1986, 251, 688–696. [Google Scholar] [CrossRef]
- Wariishi, H.; Valli, K.; Gold, M.H. Manganese (II) oxidation by manganese peroxidase from basidiomycete Phanoerochaete chrysosporium. Kinetic mechanism and the role of chelators. J. Biol. Chem. 1992, 267, 23688–23695. [Google Scholar] [PubMed]
- Kuan, I.C.; Tien, M. Stimulation Mn peroxidase activity: A possible role for oxalate in lignin biodegradation. Proc. Natl. Acad. Sci. USA 1993, 90, 1242–1246. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Dolphin, D. The role of manganese in model systems related to lignin biodegradation. Holzforchung 1990, 44, 279–283. [Google Scholar] [CrossRef]
- Glen, J.K.; Morgan, M.A.; Mayfield, M.B.; Kuwahara, M.; Gold, M.H. An extracellular H2O2-requiring enzyme preparation involved in lignin biodegradation by the white rot basidiomycete Phanerochaete chrysosporium. Biochem. Biophys. Res. Commun. 1983, 114, 1077–1083. [Google Scholar] [CrossRef]
- Tien, M.; Kirk, T.K.; Bull, C.; Fee, J.A. Steady-state and transient-state kinetics studies on the oxidation of 3,4-dimethoxybenzyl alcohol catalyzed by ligninase of Phanerochaete chrysosporium Burds. J. Biol. Chem. 1986, 264, 1687–1693. [Google Scholar]
- Ayers, A.R.; Ayers, S.B.; Eriksson, K.E. Cellobiose oxidase, purification and partial characterization of a hemoprotein from Sporotrichumpulverulentum. Eur. J. Biochem. 1978, 90, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, G.; Johansson, G.; Pettersson, G. A critical review of cellobiose dehydrogenases. J. Biotechnol. 2000, 78, 93–113. [Google Scholar] [CrossRef]
- Ludwig, R.; Harreither, W.; Tasca, F.; Gorton, L. Cellobiose dehydrogenase; a versatile catalyst for electrochemical applications. Chem. Phys. Chem. 2010, 11, 2674–2697. [Google Scholar] [CrossRef] [PubMed]
- Tasca, F.; Gorton, L.; Kijawa, M.; Patel, I.; Harreither, W.; Peterbauer, C.K.; Ludwig, R.; Nöll, G. Increasing the coulombic efficiency of glucose biofuels cell anodes by combination of redox enzymes. Biosens. Bioelectron. 2010, 25, 1710–1716. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, R.; Oritz, R.; Schultz, C.; Harreither, W.; Sygmund, C.; Gorton, L. Cellobiose dehydrogenase modified electrodes: Advanced materials science and biochemical engineering. Anal. Bioanal. Chem. 2013, 405, 3637–3658. [Google Scholar] [CrossRef] [PubMed]
- Harreither, W.; Sygmund, C.; Augustin, M.; Narcisco, M.; Rabinovich, M.L.; Gorton, L.; Haltrich, D.; Ludwig, R. Catalytic properties and classification of cellobiose dehydrogenases from ascomycetes. Appl. Environ. Microbiol. 2011, 77, 1804–1815. [Google Scholar] [CrossRef] [PubMed]
- Tasca, F.; Zafar, M.N.; Harreither, W.; Nöll, G.; Ludwig, R.; Gorton, L. A third generation glucose biosensor based on cellobiose dehydrogenase from Corynascus thermophilus and single-walled carbon nanotubes. Analyst 2011, 136, 2033–2036. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.; Kittl, R.; Ludwig, R.; Gorton, L. Direct electron transfer from the FAD cofactor of cellobiose dehydrogenase to electrodes. ACS Catal. 2016, 6, 555–563. [Google Scholar] [CrossRef]
- Bennetto, H.P.; Stirling, J.L.; Tanaka, K.; Vega, C.A. Anodic reactions in microbial fuel cells. Biotechnol. Bioeng. 1983, 25, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.L.; Walker, C.W., Jr. Biological fuel cell and an application as a reserve power source. J. Power Sources 2006, 160, 123–139. [Google Scholar] [CrossRef]
- Permana, D.; Rosdianti, D.; Ishmayana, S.; Rachman, S.; Putra, H.; Rahayuningwulan, D.; Hariyadi, H. Preliminary investigation of electricity production using dual chamber microbial fuel cell (DCMFC) with S. cerevisiae as biocatalyst and methylene blue as an electron mediator. Procedia Chem. 2015, 17, 36–43. [Google Scholar] [CrossRef]
- Gunawardena, A.; Fernando, S.; To, F. Performance of a yeast-mediated biological fuel cell. Int. J. Mol. Sci. 2008, 9, 1893. [Google Scholar] [CrossRef] [PubMed]
- Kasem, E.; Tsujiguchi, T.; Nakagawa, N. Effect of metal modification to carbon paper anodes on the performance of yeast-based microbial fuel cells part I: In the case without exogenous mediator. Key Eng. Mater. 2013, 534, 76–81. [Google Scholar] [CrossRef]
- Kasem, E.; Tsujiguchi, T.; Nakagawa, N. Effect of metal modification to carbon paper anodes on the performance of yeast-based microbial fuel cells part II: In the case with exogenous mediator, methylene blue. Key Eng. Mater. 2013, 534, 82–87. [Google Scholar] [CrossRef]
- Christwardana, M.; Kwon, Y. Yeast and carbon nanotube based biocatalyst developed by synergetic effects of covalent bonding and hydrophobic interaction for performance enhancement of membraneless microbial fuel cell. Bioresour. Technol. 2017, 225, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Rahimnejad, M.; Najafpour, G.D.; Ghoreyshi, A.A.; Talebnia, F.; Premier, G.C.; Bakeri, G.; Kim, J.R.; Oh, S.-E. Thionine increases electricity generation from microbial fuel cell using S. cerevisiae and electrogenic mixed culture. J. Microbiol. 2012, 50, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Ganguli, R.; Dunn, B.S. Kinetics of anode reactions for a yeast-catalysed microbial fuel cell. Fuel Cell 2009, 9, 44–52. [Google Scholar] [CrossRef]
- Fishilevich, S.; Amir, L.; Fridman, Y.; Aharoni, A.; Alfonta, L. Surface display of redox enzymes in microbial fuel cells. J. Am. Chem. Soc. 2009, 131, 12052–12053. [Google Scholar] [CrossRef] [PubMed]
- Gal, I.; Schlesinger, O.; Amir, L.; Alfonta, L. Yeast surface display of dehydrogenases in microbial fuel-cells. Bioelectrochemistry 2016, 112, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Raghavulu, V.; Goud, R.K.; Sarma, P.N.; Mohan, S.V. S. cerevisiae as anodic biocatalyst for power generation in biofuels cell: Influence of redox condition and substrate load. Bioresour. Technol. 2011, 102, 2751–2757. [Google Scholar] [CrossRef] [PubMed]
- Sayed, E.; Tsujiguchi, T.; Nakagawa, N. Catalytic activity of baker’s yeast in a mediator-less microbial fuel cell. Bioelectrochemistry 2012, 86, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Rah imnejad, M.; Ghoreyshi, A.A.; Najafpour, G.D.; Jafary, T. Power generation from organic substrate in batch and continuous flow microbial fuel cell operations. Appl. Eng. 2011, 88, 3999–4004. [Google Scholar] [CrossRef]
- Rahimnejad, M.; Najafpour, G.D.; Ghoreyshi, A.A.; Shakeri, M.; Zare, H. Methylene blue as electron promoters in microbial fuel cell. Int. J. Hydrogen Energy 2011, 36, 13335–13341. [Google Scholar] [CrossRef]
- Arbianti, R.; Hermansyah, H.; Utami, T.S.; Zahara, N.C.; Trisnawati, I.; Kristin, E. The usage of Saccharomyces cerevisiae in microbial fuel cell system for electricity energy production. J. Chem. Eng. 2012, 6, 814–819. [Google Scholar]
- Lin, T.; Bai, X.; Hu, Y.; Li, B.; Yuan, Y.-J.; Song, H.; Yang, Y.; Wang, J. Synthetic Saccharomyces cerevisiae-Shewanella oneidensis consortium enables glucose-fed high-performance microbial fuel cell. AIChE J. 2017, 63, 1830–1838. [Google Scholar] [CrossRef]
- Hubenova, Y.; Mitov, M. Potential application of Candida melibiosica in biofuel cells. Bioelectrochemistry 2010, 78, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Babanova, S.; Hubenova, Y.; Mitov, M. Influence of artificial mediators on yeast-based fuel cell performance. J. Biosci. Bioeng. 2011, 112, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Hubenova, Y.V.; Rashkov, R.S.; Buchvarov, V.D.; Arnaudova, M.H.; Babanova, S.M.; Mitov, M.Y. Improvement of yeast−biofuel cell output by electrode modifications. Ind. Eng. Chem. Res. 2011, 50, 557–562. [Google Scholar] [CrossRef]
- Haslett, N.D.; Rawson, F.J.; Barriëre, F.; Kunze, G.; Pasco, N.; Gooneratne, R.; Baronian, K.H.R. Characterization of yeast microbial fuel cell with the yeast Arxula adeninivorans as the biocatalyst. Biosens. Bioelectron. 2011, 26, 3742–3747. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-Y.; Kim, T.G.; Cho, K.-S. Isolation and characterization of a novel electricity-producing yeast, Candida sp. IR11. Bioresour. Technol. 2015, 192, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Hubenova, Y.; Rashkov, R.; Buchvarov, V.; Babanova, S.; Mitov, M. Nanomodified NiFe- and NiFeP-carbon felt as anode electrocatalysts in yeast-biofuel cell. J. Mater. Sci. 2011, 46, 7074–7081. [Google Scholar] [CrossRef]
- Kaneshiro, H.; Takano, K.; Takada, Y.; Wakisaka, T.; Tachibana, T.; Azuma, M. A milliliter-scale yeast-based fuel cell with high performance. Biochem. Eng. J. 2014, 83, 90–96. [Google Scholar] [CrossRef]
- Islam, M.A.; Ethiraj, B.; Cheng, C.K.; Youuf, A.; Thiruvenkadam, S.; Prasad, R.; Khan, M.M.R. Enhanced current generation using mutualistic interaction of yeast-bacterial coculture in dual chamber microbial fuel cell. Ind. Eng. Chem. Res. 2018, 57, 813–821. [Google Scholar] [CrossRef]
- Carbajosa, S.; Malki, M.; Caillard, R.; Lopez, M.F.; Palomares, F.J.; Martin-Gago, J.A.; Rodríguez, N.; Amils, R.; Fernández, V.M.; De Lacey, A.L. Electrochemical growth of Acidithiobacillus ferrooxidans on a graphite electrode for obtaining a biocathode for direct electrocatalytic reduction of oxygen. Biosens. Bioelectron. 2010, 26, 877–880. [Google Scholar] [CrossRef] [PubMed]
- Clauwaert, P.; van der Ha, D.; Boon, N.; Verbeken, K.; Verhaege, M.; Rabaey, K.; Verstraete, W. Open air biocathode enables effective electricity generation with microbial fuel cell. Environ. Sci. Technol. 2007, 41, 7564–7569. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Liu, X.-W.; Li, W.-W.; Sheng, G.-P.; Zang, G.-L.; Cheng, Y.-Y.; Shen, N.; Yang, Y.-P.; Yu, H.-Q. A white-rot fungus is used as a biocathode to improve electricity production of a microbial fuel cell. Appl. Energy 2012, 98, 594–596. [Google Scholar] [CrossRef]
- Watanabe, K. Recent development in microbial fuel technologies for sustainable bioenergy. J. Biosci. Bioeng. 2008, 6, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.E.; Logan, B.E. Proton exchange membrane and electrode surface areas as factors that affect power generation in microbial fuel cells. Appl. Microbiol. Biotechnol. 2006, 70, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Fernández de Dios, M.Á.; González del Campo, A.; Fernández, F.J.; Rodrigo, M.; Pazos, M.; Sanromán, M.Á. Bacterial-fungal interactions enhance power generation in microbial fuel cells and drive dye decolourisation by an ex situ and in situ electro-Fenton process. Bioresour. Technol. 2013, 148, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-Y.; Wu, C.-H.; Meng, C.-T.; Lin, C.-W. Decolorization of azo dye and generation of electricity by microbial fuel cell with laccase-producing white-rot fungus on cathode. Appl. Energy 2017, 188, 392–398. [Google Scholar] [CrossRef]
- Lai, C.-Y.; Liu, S.-H.; Wu, G.-P.; Lin, C.-W. Enhanced bio-decolorization of acid orange 7 and electricity generation in microbial fuel cells with superabsorbent-containing membrane and laccase-based bio-cathode. J. Clean. Prod. 2017, 166, 381–386. [Google Scholar] [CrossRef]
- Chaijak, P.; Sukkase, C.; Lertworapreecha, M.; Boonsawang, P.; Wijasika, S.; Sato, C. Enhancing electricity generation using a laccase-based microbial fuel cell with yeast Galactomyces reessii on the cathode. J. Microbiol. Biotechnol. 2018, 28, 1360–1366. [Google Scholar] [PubMed]
- Morant, K.V.; da Silva, P.H.; de Campos-Takaki, G.M.; La Rotta Hernández, C.E. Isolation and bioelectrochemical characterization of novel fungal sources with oxidasic activity applied in situ for the cathodic oxygen reduction in microbial fuel cells. Enzym. Microb. Technol. 2014, 66, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Rosales, E.; Pazos, M.; Sanroman, M.A. Advances in the electro-Fenton process for remediation of recalcitrant organic compounds. Chem. Eng. Technol. 2012, 35, 609–617. [Google Scholar] [CrossRef]
- Rozendal, R.A.; Leone, E.; Keller, J.; Rabaey, K. Efficient hydrogen peroxide generation from organic matter in a bioelectrochemical system. Electrochem. Commun. 2009, 11, 1752–1755. [Google Scholar] [CrossRef]
- Fu, L.; You, S.; Yang, F.; Gao, M.; Fang, X.; Zhang, G. Synthesis of hydrogen peroxide in microbial fuel cell. J. Chem. Technol. Biotechnol. 2010, 85, 715–719. [Google Scholar] [CrossRef]
- Furuno, S.; Foss, S.; Wild, E.; Jones, K.C.; Semple, K.T.; Harms, H.; Wick, L.Y. Mycelia promote active transport and spatial dispersion of polycyclic aromatic hydrocarbons. Environ. Sci. Technol. 2012, 46, 5463–5470. [Google Scholar] [CrossRef] [PubMed]
- Fernando, E.; Keshavarz, T.; Kyazze, G. Enhanced bio-decolourisation of acid orange 7 by Shewanella oneidensis through co-metabolism in a microbial fuel cell. Int. Biodeterior. Biodegrade 2012, 72, 1–9. [Google Scholar] [CrossRef]
- Mani, P.; Keshavar, T.; Chnadra, T.S.; Kyazze, G. Decolourisation of acid orange 7 in a microbial fuel cell with a laccase-based biocathode: Influence of mitigating pH changes in the cathode chamber. Enzym. Microb. Technol. 2017, 96, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Watson, V.J.; Logan, B.E. Power production in MFCs inoculated with Shewanella oneidensis MR-1 or mixed cultures. Biotechnol. Bioeng. 2010, 105, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E. Exoelectrogenic bacteria that power microbial fuel cells. Nat. Rev. Microbiol. 2009, 7, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Chandrasekhar, A.; Sathiyamoorthi, E.; Kim, B.S. Electro-fermentation in aid of bioenergy and biopolymers. Energies 2018, 11, 343. [Google Scholar] [CrossRef]


{kind=link}
| Fungus | MFC Type | Cathode | Anode Material | Separator | Substrate | Electron Transfer Mechanism | Max. Power Density | Ref. | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Electron Acceptor | Cathode Material | Mediatorless | Exogenous Mediator | mW/m2 | mW/m3 | ||||||
| Saccharomyces cerevisiae | Double chamber | Potassium ferricyanide | Platinium mesh | Platinium mesh | Nafion | glucose | N/A | MB | 65 | N/A | [41] |
| S. cerevisiae | Dual chamber | Potassium permanganate | Copper electrode | Copper electrode | SPEEK | glucose | + N/A | MB | 2.12 4.48 | N/A | [42] |
| S. cerevisiae | Double chamber | Potassium ferricyanide | RVC | RVC | Nafion | Glucose | + N/A | N/A MB | N/A N/A | 40 146.71 ± 7.7 | [43] |
| S. cerevisiae | Single chamber | O2 (air) | Pt/C over carbon paper | Carbon paper Co sputtered carbon paper Au sputtered carbon paper | Nafion | glucose | + | N/A | 12.9 20.2 2 | N/A | [44] |
| S. cerevisiae | Single chamber | O2 (air) | Pt/C over carbon paper | Carbon paper Co sputtered carbon paper Au sputtered carbon paper | Nafion | glucose | N/A | MB | 80 148 120 | N/A | [45] |
| S. cerevisiae | Single chamber | O2 (air) | CNT | Bare CNT/GA/Yeast PEI/CNT | Membrane less | glucose | + | N/A | 138 344 | N/A | [46] |
| S. cerevisiae | Double chamber | Potassium ferricyanide | RVC | RVC | Nafion | dextrose | N/A | MB NR MB and NR | 400 80 500 | N/A | [13] |
| S. cerevisiae | Double chamber | O2 (air) | Graphite plate | Graphite plate | Nafion | glucose | + N/A | Thionine | 3 60 | N/A | [47] |
| S. cerevisiae | Double chamber | Potassium ferricyanide | Carbon felt | Carbon felt | Nafion | glucose | N/A | MB | 1500 | N/A | [48] |
| S. cerevisiae | Double chamber | O2 (air) | Graphite plate | Graphite plate | Nafion | glucose | N/A | MB | ca. 7 | N/A | [49] |
| S. cerevisiae displaying GOx | Double chamber | O2 (air) | Graphite plate | Graphite plate | Nafion | glucose | N/A | MB | 13.6 | N/A | [49] |
| S. cerevisiae | Double chamber | O2 (air) | Graphite rods | Graphite rods/MWCNT | Nafion | lactose | + | N/A | 2.7 | N/A | [50] |
| S. cerevisiae displaying CDH | Double chamber | O2 (air) | Graphite rods | Graphite rods/MWCNT | Nafion | lactose | + | N/A | 33 | N/A | [50] |
| S. cerevisiae displaying GOx | Double chamber | O2 (air) | Graphite rods | Graphite rods/MWCNT | Nafion | d-glucose | + | N/A | 2.8 | N/A | [50] |
| S. cerevisiae | Single chamber air cathode | O2 (air) | Pt/C over carbon cloth | Graphite plate | Teflon | d-xylose | N/A | MB | 8 | N/A | [50] |
| S. cerevisiae displaying PDH | Single chamber air cathode | O2 (air) | Pt/C over carbon cloth | Graphite plate | Teflon | d-glucose d-xylose l-arabinose d-cellobiose d-galactose | N/A | MB | 31 39 32 22 14 | N/A | [50] |
| S. cerevisiae | Single chamber | O2 (air) | Graphite plate | Graphite plate | Nafion | Synthetic wastewater | + | N/A | 25.51 | N/A | [51] |
| S. cerevisiae | Single chamber/air cathode | O2 (air) | Pt/C over carbon paper | Carbon paper | Nafion | glucose | + | N/A | 3 | 17 | [52] |
| S. cerevisiae | Double chamber | O2 (air) Potassium permanganate | Graphite plates | Graphite plates | Nafion | glucose | N/A | NR | 60 133 | N/A | [53] |
| S. cerevisiae | Double chamber | O2 (air) | Graphite plate | Graphite plate | Nafion | glucose | + N/A | N/A MB | 0.414 2 | N/A | [54] |
| S. cerevisiae | Double chamber | Potassium ferricyanide | graphite | Graphite | Nafion | glucose | N/A | Riboflavin | 33 | N/A | [55] |
| S. cerevisiae-Shewanellaoneidensis | Double chamber | Potassium ferricyanide | Carbon cloth | Carbon cloth | Nafion | glucose | + | N/A | 123.4 | N/A | [56] |
| Candida melibiosica 2491 | Double chamber | Potassium ferricyanide | Plane graphite rods | Plane graphite rods | Salt bridge | Fructose YPfru YPfru | + + N/A | N/A N/A MB | N/A | 60 180 185 | [57] |
| C. melibiosica | Double chamber | Potassium ferricyanide | Carbon felt | Carbon felt | Nafion | YPfru | + N/A N/A N/A N/A N/A | N/A BcG NR MR MO MB | 20 46 89 113 137 640 | N/A | [58] |
| C. melibiosica | Double chamber | Potassium ferricyanide | Carbon felt | NME Ni(g) Ni(p) | Nafion | Fructose | + | N/A | 36 720 390 | N/A | [59] |
| Arxulaadeninivorans | Continuous flow, dual chamber | Potassium permanganate | Carbon fibre cloth | Carbon fibre cloth | Nafion | Dextrose and glucose | + N/A | N/A TMPD | 28 1030 | N/A | [60] |
| Candida sp.IR11 | Single chamber | O2 (air) | ADE 75 | Carbon felt | N/A | glucose rejected wastewater | + | N/A | 20.6 ± 1.52 | N/A | [61] |
| C. melibiosica | Double chamber | Potassium ferricyanide | Carbon felt | NME NiFe-(g) NiFe-(p) NiFeP-(g) NiFeP-(p) | Nafion | YPfru | + | N/A | 52 ± 9 83.2 93.6 155 ± 6 260 ± 8 | N/A | [62] |
| Hansenulaanomala | Double chamber | Potassium ferricyanide | Plain graphite | Plain graphite Plane graphite/polyaniline-Pt Graphite felt | N/A | Glucose, dextrose, malt extract | + | N/A | N/A | 690–2900 | [11] |
| Kluyveromycesmarxianus | dual chamber | Potassium ferricyanide | Carbon rods | carbon fiber bundles | Cation-specific membrane | glucose | N/A | 2-hydroxy-1,4-naphthoquinone | N/A | 850,000 | [63] |
| Lipomycesstarkeyi-Klebsiellapneumoniae | Dual chamber | Potassium permanganate | Stainless steel | stainless steel | Nafion | Palm oil mill effluent | + | N/A | N/A | 12,870 | [64] |
| Microorganism | MFC Type | Cathode | Anode Material | Separator | Anolyte | Max. Power Density | Ref. | ||
|---|---|---|---|---|---|---|---|---|---|
| Electron Acceptor | Cathode Material | mW/m2 | mW/m3 | ||||||
| Trametes versicolor | Double chamber/H-type | O2 (air) | Activated carbon fiber | Activated carbon fiber | Proton exchange membrane | Potassium ferricyanide | N/A | 320 ± 30 | [67] |
| T. versicolor-S. oneidensis | Double chamber/H-type | O2 (air) | Graphite rods | Graphite rods | Sterion cation exchange membrane | Acetate | N/A | 1200 | [70] |
| Ganoderma lucidum | Single chamber | O2 (air) | N/A | N/A | PVA gel | Acid orange 7 | 13.38 | N/A | [71] |
| G. lucidum | Double chamber | O2 (air) | CPC rings | CPC rings | SAP-containing PVA gel | Sludge from a wastewater treatment plant from the dying industry | 207.74 | N/A | [72] |
| Galactomycesreessii | Double chamber | O2 (air) | Vulcan-carbon cloth coated with Pt Plain carbon cloth with coconut coir | Plain carbon cloth | Nafion | Rubber wastewater sludge | 59 59 | 1163 1162 | [73] |
| Rhizopus sp. | Double chamber | O2 (air) | Pt-black carbon PTFE carbon felt Pt-free-Pt carbon PTFE carbon felt | Graphite plates | Salt bridge | Potassium ferricyanide | N/A | 317.3 197.8 | [74] |
| Aspergillus sp. | Double chamber | O2 (air) | Pt-black carbon PTFE carbon felt Pt-free-Pt carbon PTFE carbon felt | Graphite plates | Salt bridge | Potassium ferricyanide | N/A | 438.16 328.73 | [74] |
| Penicillium sp. | Double chamber | O2 (air) | Pt-black carbon PTFE carbon felt Pt-free-Pt carbon PTFE carbon felt | Graphite plates | Salt bridge | Potassium ferricyanide | N/A | 344.1 288.9 | [74] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sekrecka-Belniak, A.; Toczyłowska-Mamińska, R. Fungi-Based Microbial Fuel Cells. Energies 2018, 11, 2827. https://doi.org/10.3390/en11102827
Sekrecka-Belniak A, Toczyłowska-Mamińska R. Fungi-Based Microbial Fuel Cells. Energies. 2018; 11(10):2827. https://doi.org/10.3390/en11102827
Chicago/Turabian StyleSekrecka-Belniak, Anna, and Renata Toczyłowska-Mamińska. 2018. "Fungi-Based Microbial Fuel Cells" Energies 11, no. 10: 2827. https://doi.org/10.3390/en11102827
APA StyleSekrecka-Belniak, A., & Toczyłowska-Mamińska, R. (2018). Fungi-Based Microbial Fuel Cells. Energies, 11(10), 2827. https://doi.org/10.3390/en11102827