
Impact of Tree Species and Substrates on the Microbial and Physicochemical Properties of Reclaimed Mine Soil in the Novel Ecosystems

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Soil Sampling and Measurements
2.3. Statistical Analysis
3. Results
3.1. Soil Texture and Water Holding Capacity
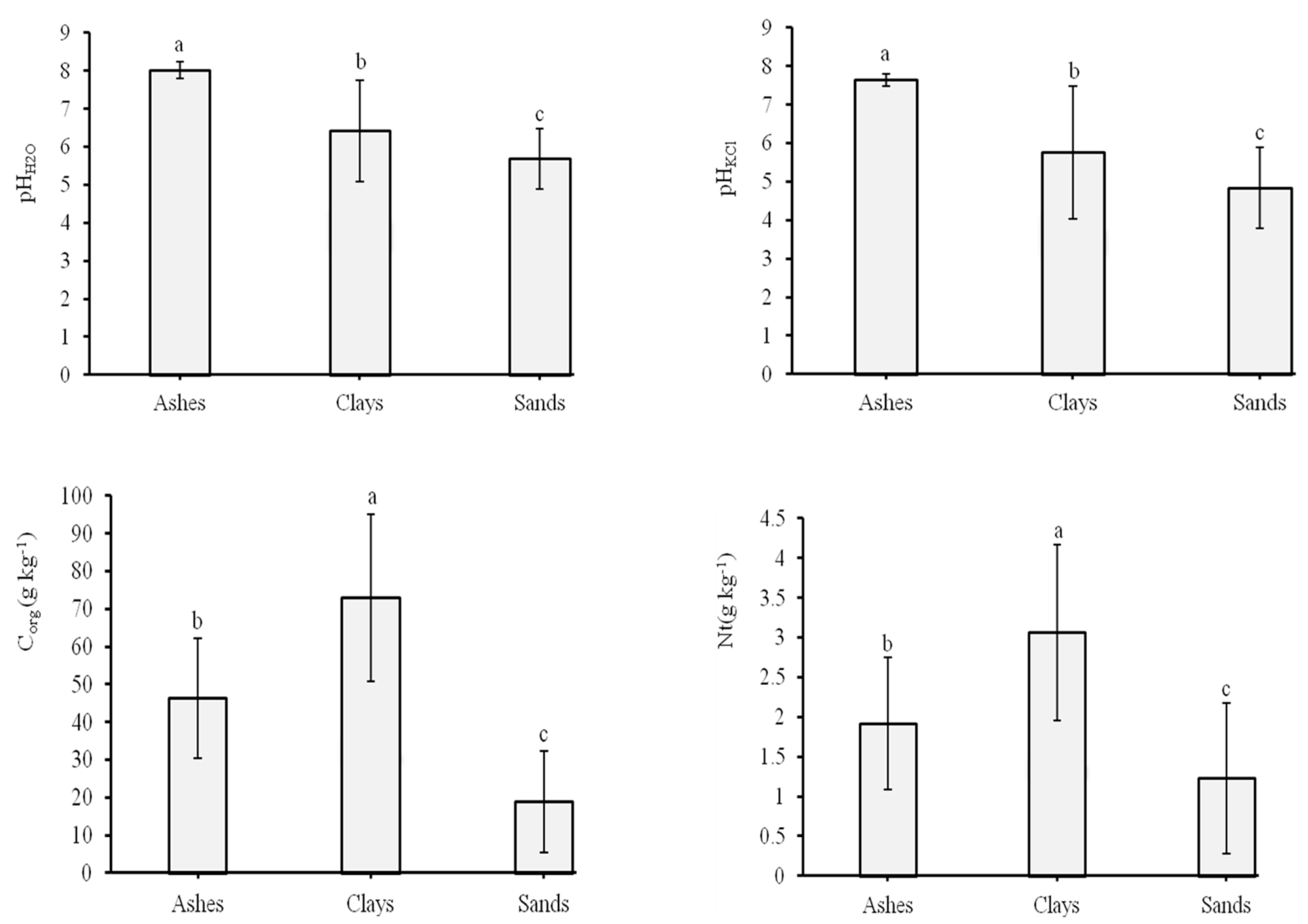
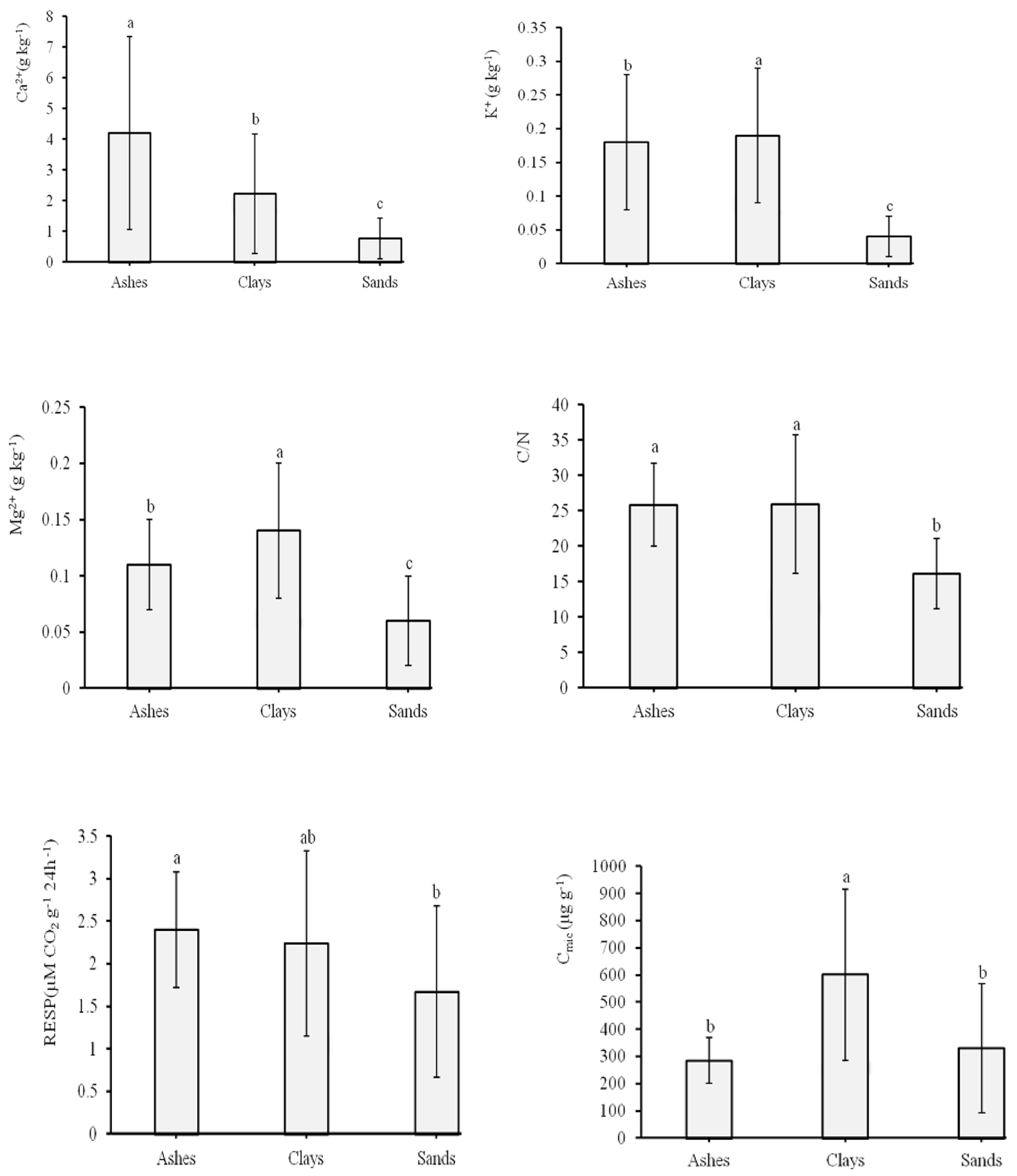
3.2. Soil Chemical Properties and Microbial Activities
3.3. Relationships between Macronutrients and Microbial Activities
4. Discussion
4.1. Tree Species Effect on Soil Properties and Microbial Activities
4.2. Substrate Effect on Soil Properties and Microbial Activities
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Knoche, D. Effects of stand conversion by thinning and under planting on water and element fluxes of a pine ecosystem (P. sylvestris L.) on lignite mine spoil. For. Ecol. Manag. 2005, 212, 214–220. [Google Scholar] [CrossRef]
- Qian, D.; Yan, C.; Xiu, L.; Feng, K. The impact of mining changes on surrounding lands and ecosystem service value in the Southern Slope of Qilian Mountains. Ecol. Complex. 2018, 36, 138–148. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Carney, K.M.; Matson, P.A. Plant Communities, Soil Microorganisms, and Soil Carbon Cycling: Does Altering the World Belowground Matter to Ecosystem Functioning? Ecosystems 2005, 8, 928–940. [Google Scholar] [CrossRef]
- Cheng, F.; Peng, X.; Zhao, P.; Yuan, J.; Zhong, C. Soil Microbial Biomass, Basal Respiration and Enzyme Activity of Main Forest Types in the Qinling Mountains. PLoS ONE 2013, 8, e67353. [Google Scholar] [CrossRef]
- Daniels, W.L.; Stewart, B.R. Reclamation of Appalachian coal refuse disposal areas. In Reclamation of Drastically Disturbed Lands; Barnhisel, R.I., Darmody, R.G., Lee Daniels, W., Eds.; Agronomy Monographs; American Society of Agronomy: Madison, WI, USA, 2000; Volume 41, pp. 433–459. [Google Scholar]
- Wanga, D.; Zhanga, B.; Zhua, L.; Yang, Y.; Li, M. Soil and vegetation development along a 10-year restoration chronosequence in tailing dams in the Xiaoqinling gold region of Central China. Catena 2018, 167, 250–256. [Google Scholar] [CrossRef]
- Pietrzykowski, M. Tree species selection and reaction to mine soil reconstructed at reforested post-mine sites: Central and eastern European experiences. Ecol. Eng. 2019, 142, 100012. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Maiti, S.K.; Masto, R.E. Use of Reclaimed Mine Soil Index (RMSI) for screening of tree species for reclamation of coal mine degraded land. Ecol. Eng. 2013, 57, 133–142. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Woś, B.; Haus, N. Scots pine needles macronutrient (N, P, K, Ca, Mg, and S) supply at different reclaimed mine soil substrates—As an indicator of the stability of developed forest ecosystems. Environ. Monit. Assess. 2013, 185, 7445–7457. [Google Scholar] [CrossRef]
- Larondelle, N.; Haase, D. Valuing post-mining landscapes using an ecosystem services approach—An example from Germany. Ecol. Indic. 2012, 18, 567–574. [Google Scholar] [CrossRef]
- Placek-Lapaj, A.; Grobelak, A.; Fijalkowski, K.; Singh, B.R.; Almås, Å.R.; Kacprzak, M. Post-Mining soil as carbon storehouse under polish conditions. J. Environ. Manag. 2019, 238, 307–314. [Google Scholar] [CrossRef]
- Wang, Z.; Lechner, A.M.; Yang, Y.; Baumgartl, T.; Wu, J. Mapping the cumulative impacts of long-term mining disturbance and progressive rehabilitation on ecosystem services. Sci. Total Environ. 2020, 717, 137214. [Google Scholar] [CrossRef]
- Laarmann, D.; Korjus, H.; Sims, A.; Kangur, A.; Kiviste, A.; Stanturf, J.A. Evaluation of afforestation development and natural colonization on a reclaimed mine site. Restor. Ecol. 2015, 23, 301–309. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Novo, L.A.B.; Pietrzykowski, M.; Maiti, S.K. Assessment of Forest Ecosystem Development in Coal Mine Degraded Land by Using Integrated Mine Soil Quality Index (IMSQI): The Evidence from India. Forests 2020, 11, 1310. [Google Scholar] [CrossRef]
- Srivastava, N.K.; Ram, L.C.; Masto, R.E. Reclamation of overburden and lowland in coal mining area with fly ash and selective plantation: A sustainable ecological approach. Ecol. Eng. 2014, 71, 479–489. [Google Scholar] [CrossRef]
- Zhao, Z.; Bai, Z.; Zhang, Z.; Guo, D.; Li, J.; Xu, Z. Population structure and spatial distributions patterns of 17 years old plantation in a reclaimed spoil of Pingshuo opencast mine, China. Ecol. Eng. 2012, 44, 147–151. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Jurkšienė, G.; Janušauskaitė, D.; Armolaitis, K.; Baliuckas, V. Leaf litterfall decomposition of pedunculate (Quercus robur L.) and sessile (Q. petraea [Matt.] Liebl.) oaks and their hybrids and its impact on soil microbiota. Dendrobiology 2017, 78, 51–62. [Google Scholar] [CrossRef]
- Kumari, S.; Maiti, S.K. Reclamation of coalmine spoils with topsoil, grass, and legume: A case study from India. Environ. Earth Sci. 2019, 78, 429. [Google Scholar] [CrossRef]
- Yan, M.; Fan, L.; Wang, L. Restoration of soil carbon with different tree species in a post-mining land in eastern Loess Plateau, China. Ecol. Eng. 2020, 158, 106025. [Google Scholar] [CrossRef]
- Singh, A.N.; Raghubanshi, A.S.; Singh, J.S. Plantations as a tool for mine spoil restoration. Curr. Sci. 2002, 82, 1436–1441. [Google Scholar]
- Frouz, J.; Keplin, B.; Pižl, V.; Tojavský, K.; Starý, J.; Lukešová, A.; Nováková, A.; Balik, V.; Háněl, L.; Materna, J.; et al. Soil biota and upper soil layer development in two contrasting post-mining chronosequences. Ecol. Eng. 2001, 17, 275–284. [Google Scholar] [CrossRef]
- Cortines, E.; Valcarcel, R. Influence of pioneer-species combinations on restoration of disturbed ecosystems in the Atlantic Forest, Rio de Janeiro, Brazil. Rev. Árvore 2009, 33, 927–936. [Google Scholar] [CrossRef][Green Version]
- Józefowska, A.; Woś, B.; Pietrzykowski, M. Tree species and soil substrate effects on soil biota during early soil forming stages at afforested mine sites. Appl. Soil Ecol. 2016, 102, 70–79. [Google Scholar] [CrossRef]
- De-Marco, A.; Esposito, F.; Berg, B.; Giordano, M.; De Santo, A. Soil C and N sequestration in organic and mineral layers of two coeval forest stands implanted on pyroclastic material (Mount Vesuvius, South Italy). Geoderma 2013, 209, 128–135. [Google Scholar] [CrossRef]
- Frouz, J.; Livečková, M.; Albrechtová, J.; Chroňáková, A.; Cajthaml, T.; Pižl, V.; Háněl, L.; Starý, J.; Baldrian, P.; Lhotáková, Z.; et al. Is the effect of trees on soil properties mediated by soil fauna? A case study from post-mining sites. For. Ecol. Manag. 2013, 309, 87–95. [Google Scholar] [CrossRef]
- Galka, B.; Labaz, B.; Bogacz, A.; Bojko, O.; Kabala, C. Conversion of Norway spruce forests will reduce organic carbon pools in the mountain soils of SW Poland. Geoderma 2014, 213, 287–295. [Google Scholar] [CrossRef]
- Šourková, M.; Frouz, J.; Fettweis, U.; Bens, O.; Hüttl, R.F.; Šantrucková, H. Soil development and properties of microbial biomass succession in reclaimed post-mining sites near Sokolov (Czech Republic) and near Cottbus (Germany). Geoderma 2005, 129, 73–80. [Google Scholar] [CrossRef]
- Chodak, M.; Niklińska, M. The effect of different tree species on the chemical and microbial properties of reclaimed mine soils. Biol. Fertil. Soils 2010, 46, 555–566. [Google Scholar] [CrossRef]
- Baldrian, P. Enzymes of saprotrophic basidiomycetes. In Ecology of Saprotrophic Basidiomycetes; Boddy, L., Frankland, J., van West, P., Eds.; Academic Press: New York, NY, USA, 2008; pp. 19–41. [Google Scholar]
- Miralles, I.; Cantón, Y.; Solé-Benet, A. Two-dimensional porosity of crusted silty soils: Indicators of soil quality in semiarid rangelands? Soil Sci. Am. J. 2011, 75, 1289–1301. [Google Scholar]
- Bashan, Y.; De-Bashan, L.E. Microbial populations of arid lands and their potential for restoration of deserts. In Soil Biology and Agriculture in the Tropics; Springer: Berlin/Heidelberg, Germany, 2010; pp. 109–137. [Google Scholar]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Kheirfam, H.; Sadeghi, S.H.R.; Homaee, M.; Zarei Darki, B. Quality improvement of an erosion-prone soil through microbial enrichment. Soil Tillage Res. 2017, 165, 230–238. [Google Scholar] [CrossRef]
- Józefowska, A.; Pietrzykowski, M.; Woś, B.; Cajthaml, T.; Frouz, J. Relationships between respiration, chemical and microbial properties of afforested mine soils with different soil texture and tree species: Does the time of incubation matter. Eur. J. Soil Biol. 2017, 80, 102–109. [Google Scholar] [CrossRef]
- Chodak, M.; Niklińska, M. Effect of texture and tree species on microbial properties of mine soils. Appl. Soil Ecol. 2010, 46, 268–275. [Google Scholar] [CrossRef]
- Chodak, M.; Sroka, K.; Pietrzykowski, M. Activity of phosphatases and microbial phosphorus under various tree species growing on reclaimed technosols. Geoderma 2021, 401, 115320. [Google Scholar] [CrossRef]
- Woś, B.; Pietrzykowski, P. Characteristics of technogenic soils developed from Neogene and Quaternary sediments substrate on reclaimed sulphur and sand extraction mine sites. Soil Sci. Annu. 2020, 71, 344–351. [Google Scholar] [CrossRef]
- Krzaklewski, W.; Pietrzykowski, M.; Woś, B. Survival and growth of alders (Alnus glutinosa (L.) Gaertn. and Alnus incana (L.) Moench) on fly ash RMS at different substrate improvement. Ecol. Eng. 2012, 49, 35–40. [Google Scholar] [CrossRef]
- Schlichting, E.; Blume, H.P. Methods of Soil Analysis; Parey: Hamburg, Germany, 1966. [Google Scholar]
- Cejpek, J.; Kuráž, V.; Frouz, J. Hydrological Properties of Soils in Reclaimed and Unreclaimed Sites after Brown-Coal Mining. Pol. J. Environ. Stud. 2013, 22, 645–652. [Google Scholar]
- Šnajdr, J.; Dobiášová, P.; Urbanová, M.; Petránková, M.; Cajthaml, T.; Frouz, J.; Baldrian, P. Dominant trees affect microbial community composition and activity in post-mining afforested soils. Soil Biol. Biochem. 2013, 56, 105–115. [Google Scholar] [CrossRef]
- Józefowska, A.; Pietrzykowski, M.; Woś, B.; Cajthaml, T.; Frouz, J. The effects of tree species and substrate on carbon sequestration and chemical and biological properties in reforested post-mining soils. Geoderma 2017, 292, 9–16. [Google Scholar] [CrossRef]
- Wang, F.; Li, Z.; Xia, H.; Zou, B.; Li, N.; Liu, J.; Zhu, W. Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China. Soil Sci. Plant Nutr. 2010, 56, 297–306. [Google Scholar] [CrossRef]
- Nohrstedt, H.-Ö. Nitrogen fixation (C2H2-reduction) in birch litter. Scand. J. For. Res. 1988, 3, 17–23. [Google Scholar] [CrossRef]
- Smolander, A. Frankia populations in soils under different tree species—With special emphasis on soils under Betula pendula. Plant Soil. 1990, 121, 1–10. [Google Scholar] [CrossRef]
- Resh, S.C.; Binkley, D.; Parrotta, J.A. Greater soil carbon sequestration under nitrogen-fixing trees compared with Eucalyptus species. Ecosystems 2002, 5, 217–231. [Google Scholar] [CrossRef]
- Ussiri, D.A.N.; Lal, R.; Jacinthe, P.A. Soil Properties and Carbon Sequestration of Afforested Pastures in Reclaimed Mine soils of Ohio. Soil Sci. Soc. Am. J. 2006, 70, 1797. [Google Scholar] [CrossRef]
- Cools, N.; Vesterdal, L.; De Vos, B.; Vanguelova, E.; Hansen, K. Tree species is the major factor explaining C:N ratios in European forest soils. For. Ecol. Manag. 2014, 311, 3–16. [Google Scholar] [CrossRef]
- Walmsley, A.; Vachová, P.; Hlava, J. Tree species identity governs the soil macrofauna community composition and soil development at reclaimed post-mining sites on calcium-rich clays. Eur. J. For. Res. 2019, 138, 753–761. [Google Scholar] [CrossRef]
- Tu, C.; Koenning, S.R.; Hu, S. Root-parasitic nematodes enhance soil microbial activities and nitrogen mineralization. Microb. Ecol. 2003, 46, 134–144. [Google Scholar] [CrossRef]
- Wang, W.J.; Smith, C.J.; Chen, D. Predicting soil nitrogen mineralization dynamics with a modified double exponential model. Soil Sci. Soc. Am. J. 2004, 68, 1256–1265. [Google Scholar] [CrossRef]
- Schulz, S.; Brankatschk, R.; Dümig, A.; Kögel-Knabner, I.; Schloter, M.; Zeyer, J. The role of microorganisms at different stages of ecosystem development for soil formation. Biogeosciences 2013, 10, 3983–3996. [Google Scholar] [CrossRef]
- Frouz, J.; Prach, K.; Pižl, V.; Háněl, L.; Starý, J.; Tajovský, K.; Materna, J.; Balik, V.; Kalcik, J.; Řehounková, K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Soil Science Division Staff. Soil Survey Manual; Ditzler, C., Scheffe, K., Monger, H.C., Eds.; USDA Handbook 18; Government Printing Office: Washington, DC, USA, 2017; 199p. [Google Scholar]
- Haynes, R.J. Reclamation and revegetation of fly ash disposal sites—Challenges and research needs. J. Environ. Manag. 2009, 90, 43–53. [Google Scholar] [CrossRef]
- Fettweis, U.; Bens, O.; Hüttl, R.F. Accumulation and properties of soil organic carbon at reclaimed sites in the Lusatian lignite mining district afforested with Pinus sp. Geoderma 2005, 129, 81–91. [Google Scholar] [CrossRef]
- Woś, B.; Pietrzykowski, M. Simulation of birch and pine litter influence on early stage of reclaimed soil formation process under controlled conditions. J. Environ. Qual. 2015, 44, 1091. [Google Scholar] [CrossRef]
- Chenu, C.; Plante, A.F. Clay-sized organo-mineral complexes in a cultivation chronosequence: Revisiting the concept of the ‘primary organo-mineral complex’. Eur. J. Soil Sci. 2006, 57, 596–607. [Google Scholar] [CrossRef]
- Laird, D.A.; Martens, D.A.; Kingery, W.L. Nature of Clay-Humic Complexes in an Agricultural Soil. Soil Sci. Soc. Ae. J. 2001, 65, 1413–1418. [Google Scholar] [CrossRef]
- Uzarowicz, Ł.; Zagórski, Z.; Mendak, E.; Bartmiński, P.; Szara, E.; Kondras, M.; Oktaba, L.; Turek, A.; Rogoziński, R. Technogenic soils (Technosols) developed from fly ash and bottom ash from thermal power stations combusting bituminous coal and lignite. Part I. Properties, classification, and indicators of early pedogenesis. Catena 2017, 157, 75–89. [Google Scholar]
- Külaots, I.; Hurt, R.H.; Suuberg, E.M. Size distribution of unburned carbon in coal fly ash and its implications. Fuel 2004, 83, 223–230. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Woś, B.; Pająk, M.; Wanic, T.; Krzaklewski, W.; Chodak, M. The impact of alders (Alnus spp.) on the physicochemical properties of technosols on a lignite combustion waste disposal site. Ecol. Eng. 2018, 120, 180–186. [Google Scholar] [CrossRef]
- Knops, J.M.H.; Tilman, D. Dynamics of soil nitrogen and carbon accumulation for 61 years after agricultural abandonment. Ecology 2000, 81, 88–98. [Google Scholar] [CrossRef]
- Zhou, Y.; Boutton, T.W.; Wu, X.B. Soil phosphorus does not keep pace with soil carbon and nitrogen accumulation following woody encroachment. Glob. Change Biol. 2018, 24, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Treschevskaya, E.; Tichonova, E.; Golyadkina, I.; Malinina, T. Soil development processes under different tree species at afforested post-mining sites. IOP Conf. Ser. Earth Environ. Sci. 2019, 226, 012012. [Google Scholar] [CrossRef]
- Nuaklong, P.; Wongsa, A.; Sata, V.; Boonserm, K.; Sanjayan, J.; Chindaprasirt, P. Properties of high-calcium and low-calcium fly ash combination geopolymer mortar containing recycled aggregate. Heliyon 2019, 5, e02513. [Google Scholar] [CrossRef] [PubMed]
- Kome, G.K.; Enang, R.K.; Tabi, F.O.; Yerima, B.P.K. Influence of Clay Minerals on Some Soil Fertility Attributes: A Review. Open J. Soil Sci. 2019, 9, 155–188. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Kapusta, P.; Szarek-Łukaszewska, G.; Grodzińska, K.; Niklińska, M.; Vogt, R.D. Soil fertility and plant diversity enhance microbial performance in metal-polluted soils. Sci. Total Environ. 2012, 439, 211–219. [Google Scholar] [CrossRef]
- Girvan, M.S.; Bullimore, J.; Pretty, J.N.; Osborn, A.M.; Ball, A.S. Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soils. Appl. Environ. Microbiol. 2003, 69, 1800–1809. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Macdonald, L.M.; Rogers, S.L.; Gregga, A.L.; Bolgerd, T.P.; Baldock, J.A. Habitat selective factors influencing the structural composition and functional capacity of microbial communities in agricultural soils. Soil Biol. Biochem. 2008, 40, 803–813. [Google Scholar] [CrossRef]
- Carletti, P.; Vendramin, E.; Pizzeghello, D.; Concheri, G.; Zanella, A.; Nardi, S.; Squartini, A. Soil humic compounds and microbial communities in six spruce forests as function of parent material, slope aspect and stand age. Plant Soil 2009, 315, 47–65. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Study Site | Mean Annual Temperature (°C) | Mean Annual Precipitation (mm Year−1) | Age of Forest Stands (Years) | Substrate Type | Reclamation Treatments |
|---|---|---|---|---|---|
| Szczakowa sand pit 50°16′ N; 19°26′ E | 8.1 | 700 | 30–35 | Quaternary sand | Cultivation of lupine (Lupinus polyphyllus Lindl.) as a green manure for 1 year, mineral fertilization with NPK (70 kg N ha−1, 120 kg P ha−1, 120 kg K ha−1), and tree planting. |
| Bełchatów external spoil heap 51°13′ N; 19° 25′ E | 7.6 | 580 | 30–35 | Quaternary sand | Surface forming and leveling, mineral fertilization with NPK (60 kg N ha−1, 70 kg P ha−1, 60 kg K ha−1), cultivation of leguminous plants and grasses for 1 year, and planting of trees. |
| Lubień lignite combustion waste disposal site 51° 27′ N, 19° 27′ E | 7.6 | 580 | 25–30 | Ashes after lignite combustion | Mineral fertilization with NPK (60 kg N ha−1, 36 kg P ha−1, 36 kg K ha−1), hydro-seeding of a mixture of grasses (Dactylis glomerata L., Lolium multiflorum Lam.) and a sewage sludge and tree planting. |
| Turów external spoil heap 50°52′ N; 14°52′ E | 8.3 | 706 | 38–44 | Acid Miocene clays | Mineral fertilization with NPK (50 kg N ha−1, 28 kg P ha−1, 16 kg K ha−1), cultivation of grasses (Festuca rubra L., Phleum pratense L.) and legumes (e.g., Lupinus polyphyllus Lindl, Melilotus albus Desr), and tree planting. |
| Piaseczno external spoil heap 50°35′ N; 21°47′ E | 7.0 | 650 | 38–44 | Alkaline Miocene clays | Mineral fertilization with NPK (80 kg N ha−1, 50 kg P ha−1 and 60 kg K ha−1), cultivation of Melilotus albus L. and grasses, and tree planting. |
| Species-Substrate | Sand (%) | Silt (%) | Clay (%) | WHC (%) |
|---|---|---|---|---|
| A-As | 71 ± 12 ab | 21 ± 10 abc | 8 ± 3 c | 97.4 ± 11.9 ab |
| A-Cl | 22 ± 8 c | 14 ± 5 bcd | 64 ± 11 a | 75.9 ± 22.3 bcd |
| A-Sa | 82 ± 7 ab | 8 ± 5 cd | 10 ± 3 c | 29.8 ± 6.4 g |
| B-As | 74 ± 9 ab | 20 ± 10 abc | 6 ± 2 c | 104.7 ± 7 a |
| B-Cl | 27 ± 8 c | 19 ± 5 abc | 54 ± 11 ab | 68.6 ± 7.7 cde |
| B-Sa | 78 ± 6 ab | 13 ± 5 bcd | 9 ± 2 c | 41.4 ± 10.3 fg |
| Bl-As | 63 ± 12 b | 30 ± 10 a | 7 ± 2 c | 93.1 ± 12.3 abc |
| Bl-Cl | 28 ± 18 c | 14 ± 3 bcd | 58 ± 18 ab | 69.8 ± 17.4 cde |
| Bl-Sa | 78 ± 8 ab | 12 ± 7 bcd | 10 ± 2 c | 47.3 ± 14.8 efg |
| P-As | 70 ± 5 b | 23 ± 5 ab | 7 ± 2 c | 86.7 ± 8.3 abcd |
| P-Cl | 37 ± 9 c | 16 ± 2 bcd | 47 ± 11 b | 62.1 ± 15 def |
| P-Sa | 89 ± 2 a | 5 ± 2 d | 6 ± 2 c | 30.7 ± 11.8 g |
| Df | SS | MS | F | P | ||
|---|---|---|---|---|---|---|
| pHH2O | TS | 3 | N.S. | N.S. | N.S. | N.S. |
| SU | 2 | 68.51 | 34.26 | 38.44 | 0.000 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. | |
| pHKCl | TS | 3 | N.S. | N.S. | N.S. | N.S. |
| SU | 2 | 98.04 | 49.02 | 32.30 | 0.000 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. | |
| Corg (g kg−1) | TS | 3 | N.S. | N.S. | N.S. | N.S. |
| SU | 2 | 35,072.52 | 17,536.26 | 62.30 | 0.01 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. | |
| Nt (g kg−1) | TS | 3 | 15.59 | 5.20 | 7.10 | 0.000 |
| SU | 2 | 40.98 | 20.49 | 28.01 | 0.000 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. | |
| Ca2+ (g kg−1) | TS | 3 | N.S. | N.S. | N.S. | N.S. |
| SU | 2 | 142.51 | 71.26 | 36.80 | 0.000 | |
| N.S. | 6 | N.S. | N.S. | N.S. | N.S. | |
| K+ (g kg−1) | TS | 3 | N.S. | N.S. | N.S. | N.S. |
| SU | 2 | 0.34 | 0.17 | 26.08 | 0.01 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. | |
| Mg2+ (g kg−1) | TS | 3 | N.S. | N.S. | N.S. | N.S. |
| SU | 2 | 0.08 | 0.04 | 13.78 | 0.000 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. | |
| C:N | TS | 3 | 1069.38 | 356.46 | 9.45 | 0.000 |
| SU | 2 | 1531.99 | 766 | 20.30 | 0.000 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. | |
| RESP (µM CO2 g−1 24 h−1) | TS | 3 | 9.51 | 3.17 | 4.18 | 0.010 |
| SU | 2 | 7.02 | 3.51 | 4.62 | 0.014 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. | |
| Cmic (µg g−1) | TS | 3 | N.S. | N.S. | N.S. | N.S. |
| SU | 2 | 1,405,123.85 | 702,561.93 | 13.34 | 0.000 | |
| TS × SU | 6 | N.S. | N.S. | N.S. | N.S. |
| Pine | Birch | Alder | Black Locust | |
|---|---|---|---|---|
| pHH2O | 6.61 ± 1.38 a | 6.78 ± 1.3 a | 6.68 ± 1.35 a | 6.74 ± 1.4 a |
| pHKCl | 5.94 ± 1.79 a | 6.2 ± 1.66 a | 6.07 ± 1.63 a | 6.09 ± 1.65 a |
| Corg (g kg−1) | 39.36 ± 32.02 a | 46.08 ± 24.64 a | 48.79 ± 28.85 a | 49.68 ± 27.81 a |
| Nt (g kg−1) | 1.38 ± 1.04 b | 2.01 ± 0.95 ab | 2.24 ± 1.23 a | 2.67 ± 1.35 a |
| Ca2+ (g kg−1) | 2.37 ± 1.97 a | 2.16 ± 2.02 a | 2.47 ± 2.22 a | 2.62 ± 1.92 a |
| K+ (g kg−1) | 0.14 ± 0.13 a | 0.13 ± 0.07 a | 0.13 ± 0.12 a | 0.17 ± 0.12 a |
| Mg2+ (g kg−1) | 0.1 ± 0.07 a | 0.11 ± 0.07 a | 0.1 ± 0.06 a | 0.1 ± 0.06 a |
| C:N | 28.87 ± 10.16 a | 22.69 ± 5.40 b | 20.34 ± 8.25 b | 18.73 ± 5.94 b |
| RESP (µM CO2 g−1 24 h −1) | 1.76 ± 0.9 b | 2.70 ± 0.9 a | 2.08 ± 1.04 ab | 1.87 ± 0.89 b |
| Cmic (µg g−1) | 377.67 ± 329.24 a | 503.32 ± 243.80 a | 362.67 ± 201.16 a | 376.46 ± 284.77 a |
| Corg | Nt | Ca2+ | K+ | Mg2+ | RESP | Cmic | |
|---|---|---|---|---|---|---|---|
| Corg | 1 | ||||||
| Nt | 0.81 * | 1 | |||||
| Ca2+ | 0.41 * | 0.49 * | 1 | ||||
| K+ | 0.62 * | 0.64 * | 0.77 * | 1 | |||
| Mg2+ | 0.75 * | 0.6 * | 0.54 * | 0.69 * | 1 | ||
| RESP | 0.48 * | 0.58 * | 0.62 * | 0.46 * | 0.54 * | 1 | |
| Cmic | 0.54 * | 0.67 * | 0.31 * | 0.42 * | 0.53 * | 0.70 * | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietrzykowski, M.; Misebo, A.M.; Pająk, M.; Woś, B.; Sroka, K.; Chodak, M. Impact of Tree Species and Substrates on the Microbial and Physicochemical Properties of Reclaimed Mine Soil in the Novel Ecosystems. Forests 2022, 13, 1858. https://doi.org/10.3390/f13111858
Pietrzykowski M, Misebo AM, Pająk M, Woś B, Sroka K, Chodak M. Impact of Tree Species and Substrates on the Microbial and Physicochemical Properties of Reclaimed Mine Soil in the Novel Ecosystems. Forests. 2022; 13(11):1858. https://doi.org/10.3390/f13111858
Chicago/Turabian StylePietrzykowski, Marcin, Amisalu Milkias Misebo, Marek Pająk, Bartłomiej Woś, Katarzyna Sroka, and Marcin Chodak. 2022. "Impact of Tree Species and Substrates on the Microbial and Physicochemical Properties of Reclaimed Mine Soil in the Novel Ecosystems" Forests 13, no. 11: 1858. https://doi.org/10.3390/f13111858
APA StylePietrzykowski, M., Misebo, A. M., Pająk, M., Woś, B., Sroka, K., & Chodak, M. (2022). Impact of Tree Species and Substrates on the Microbial and Physicochemical Properties of Reclaimed Mine Soil in the Novel Ecosystems. Forests, 13(11), 1858. https://doi.org/10.3390/f13111858