
Evolocumab Reduces Oxidative Stress and Lipid Peroxidation in Obese Zucker Rats



Abstract

1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals and Treatment
2.3. Blood Pressure and Weight Parameters
2.4. Glucose and Fructosamine Levels and Lipid Profile
2.5. Thiobarbituric Acid Reactive Substances (TBARS) and Conjugated Diene (CD) Concentrations
2.6. Western Blot Analysis
2.7. Total NO Synthase (NOS) Activity
2.8. Statistical Analysis
3. Results
3.1. Weight Parameters, Blood Pressure, and Glucose and Fructosamine Levels
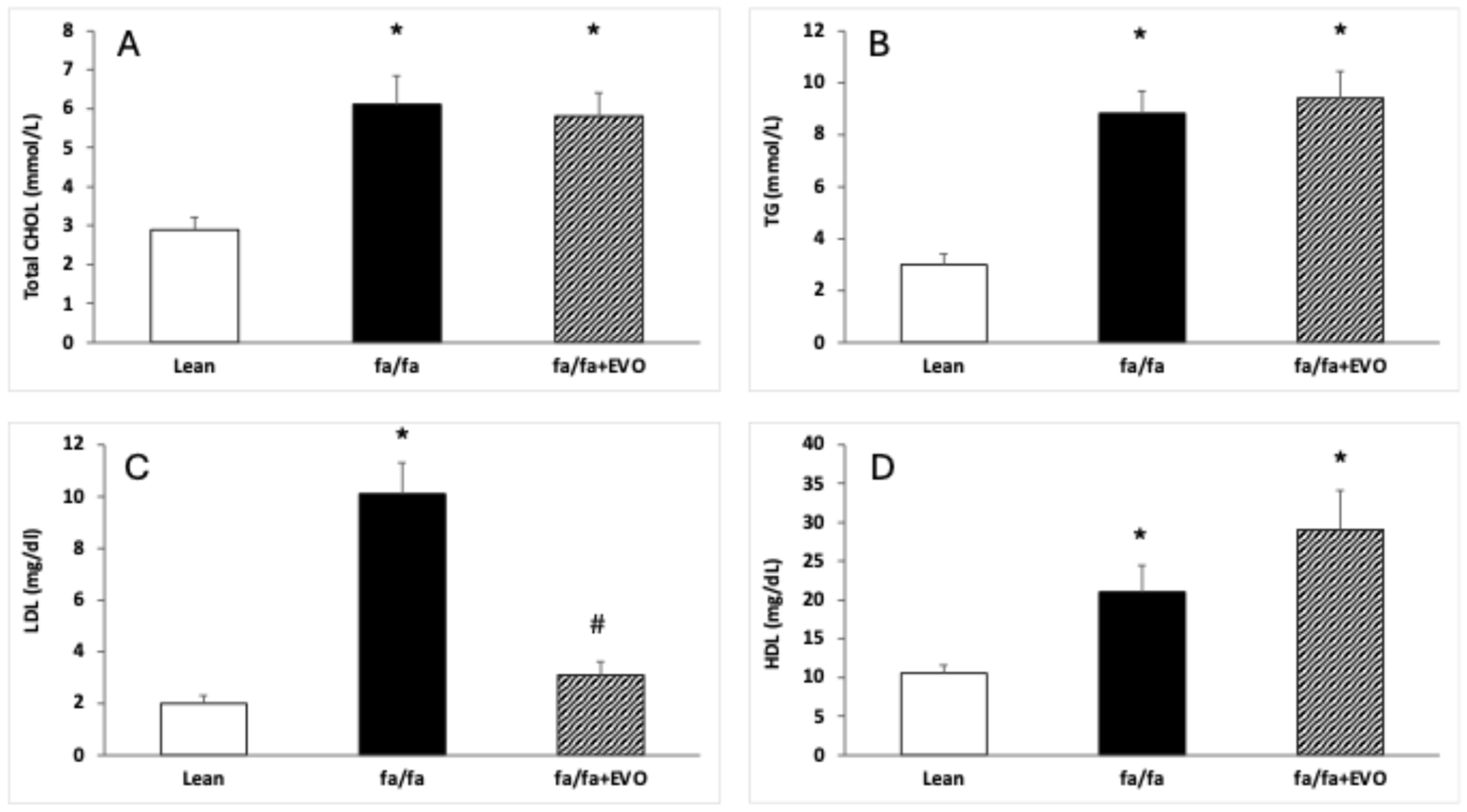
3.2. Lipid Profile
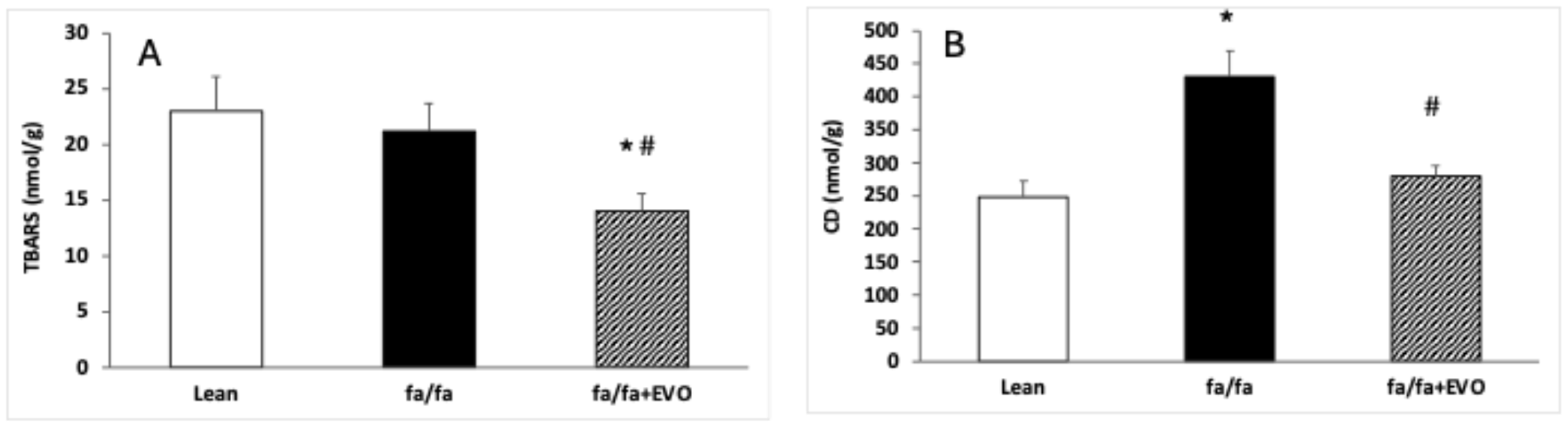
3.3. TBARS and Conjugated Diene Concentrations
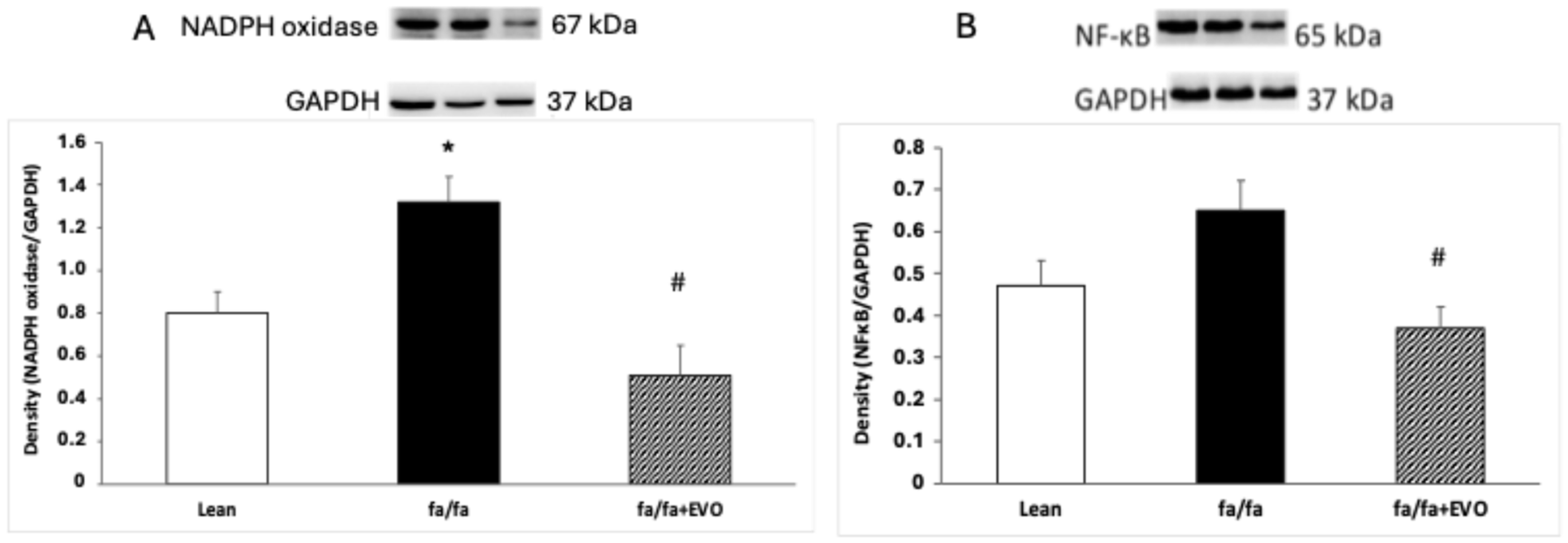
3.4. NADPH Oxidase and NF-κB Protein Expression
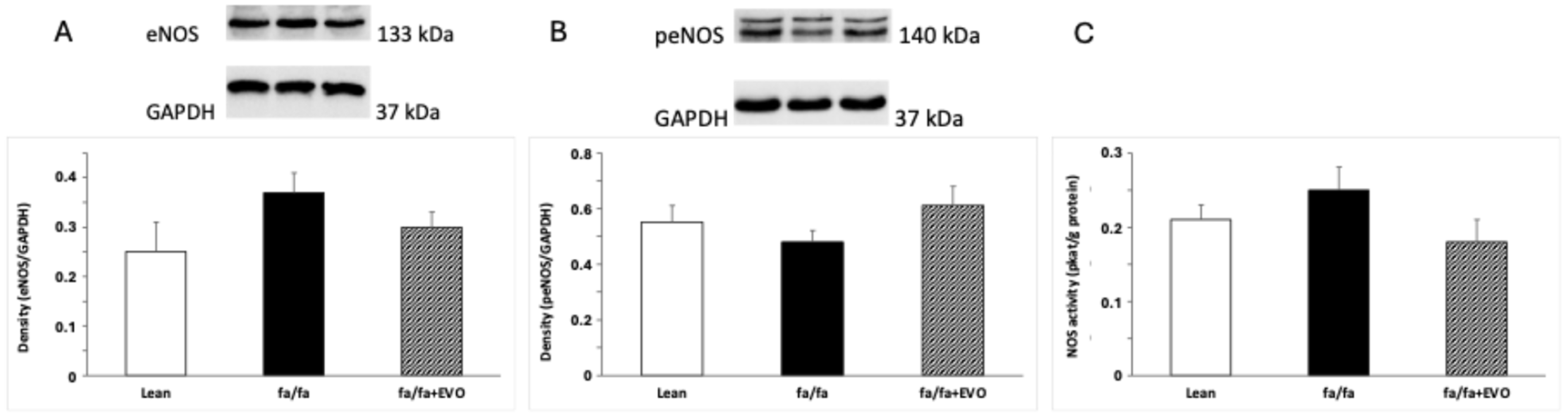
3.5. Endothelial NOS (eNOS) and Phosphorylated eNOS (peNOS) Protein Expression and Total NOS Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaggini, M.; Gorini, F.; Vassalle, C. Lipids in Atherosclerosis: Pathophysiology and the Role of Calculated Lipid Indices in Assessing Cardiovascular Risk in Patients with Hyperlipidemia. Int. J. Mol. Sci. 2022, 24, 75. [Google Scholar] [CrossRef] [PubMed]
- LaRosa, J.C.; Pedersen, T.R.; Somaratne, R.; Wasserman, S.M. Safety and effect of very low levels of low-density lipoprotein cholesterol on cardiovascular events. Am. J. Cardiol. 2013, 111, 1221–1229. [Google Scholar] [CrossRef]
- Dayar, E.; Pechanova, O. Targeted Strategy in Lipid-Lowering Therapy. Biomedicines 2022, 10, 1090. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, C.; Wang, X.; Briggs, M.R.; Admon, A.; Wu, J.; Hua, X.; Goldstein, J.L.; Brown, M.S. SREBP-1, a basic-helix-loop-helix-leucine zipper protein that controls transcription of the low density lipoprotein receptor gene. Cell 1993, 75, 187–197. [Google Scholar] [CrossRef]
- Antonopoulos, A.S.; Margaritis, M.; Lee, R.; Channon, K.; Antoniades, C. Statins as anti-inflammatory agents in atherogenesis: Molecular mechanisms and lessons from the recent clinical trials. Curr. Pharm. Des. 2012, 18, 1519–1530. [Google Scholar] [CrossRef]
- Şaman, E.; Cebova, M.; Barta, A.; Koneracka, M.; Zavisova, V.; Eckstein-Andicsova, A.; Danko, M.; Mosnacek, J.; Pechanova, O. Combined Therapy with Simvastatin- and Coenzyme-Q10-Loaded Nanoparticles Upregulates the Akt-eNOS Pathway in Experimental Metabolic Syndrome. Int. J. Mol. Sci. 2022, 24, 276. [Google Scholar] [CrossRef] [PubMed]
- Potgieter, M.; Pretorius, E.; Pepper, M.S. Primary and secondary coenzyme Q10 deficiency: The role of therapeutic supplementation. Nutr. Rev. 2013, 71, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Zozina, V.I.; Covantev, S.; Goroshko, O.A.; Krasnykh, L.M.; Kukes, V.G. Coenzyme Q10 in cardiovascular and metabolic diseases: Current state of the problem. Curr. Cardiol. Rev. 2018, 14, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Ward, N.C.; Reid, C.M.; Watts, G.F. Low-density lipoprotein-cholesterol lowering effect of a nutraceutical regimen with or without ezetimibe in hypercholesterolaemic patients with statin intolerance. Front. Cardiovasc. Med. 2022, 9, 1060252. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S. Development of proprotein convertase subtilisin/kexin type 9 inhibitors and the clinical potential of monoclonal antibodies in the management of lipid disorders. Vasc. Health Risk Manag. 2016, 12, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Abifadel, M.; Varret, M.; Rabès, J.-P.; Allard, D.; Ouguerram, K.; Devillers, M.; Cruaud, C.; Benjannet, S.; Wickham, L.; Erlich, D.; et al. Mutations in PCSK9 cause autosomal dominant hypercholesterolemia. Nat. Genet. 2003, 34, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Moon, Y.-A.; Horton, J.D. Post-transcriptional regulation of low density lipoprotein receptor protein by proprotein convertase subtilisin/kexin type 9a in mouse liver. J. Biol. Chem. 2004, 279, 50630–50638. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicki, A.S.; Hardman, T.C.; Viljoen, A. Inhibition of pro-protein convertase subtilisin kexin 9 (PCSK-9) as a treatment for hyperlipidaemia. Expert Opin. Investig. Drugs 2012, 21, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, J.; Chen, H.; Zhang, T.; He, D.; Luo, Q.; Chi, J.; Hong, Z.; Liao, Y.; Zhang, S.; et al. PCSK9 Inhibition: From Current Advances to Evolving Future. Cells 2022, 11, 2972. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.-H.; Peng, J.; Ren, Z.; Yang, J.; Li, T.-T.; Li, T.-H.; Wang, Z.; Wei, D.-H.; Liu, L.-S.; Zheng, X.-S.; et al. New role of PCSK9 in atherosclerotic inflammation promotion involving the TLR4/NF-κB pathway. Atherosclerosis 2017, 262, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Lepor, N.E.; Cantón, G.; Contreras, L.; Hippe, D.S.; Isquith, D.A.; Balu, N.; Kedan, I.; Simonini, A.A.; Yuan, C.; et al. Serial magnetic resonance imaging detects a rapid reduction in plaque lipid content under PCSK9 inhibition with alirocumab. Int. J. Cardiovasc. Imaging 2021, 37, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Liu, M.; Li, Y.; Zhu, Q.; Su, G. Effect of evolocumab on the progression and stability of atherosclerotic plaques as evaluated by grayscale and iMAP-IVUS. Ann. Palliat. Med. 2020, 9, 3078–3088. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Colon, E.; Daum, A.; Yosefy, C. Statins and PCSK9 inhibitors: A new lipid-lowering therapy. Eur. J. Pharmacol. 2020, 878, 173114. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Chaparro-Riggers, J.; Strop, P.; Geng, T.; Sutton, J.E.; Tsai, D.; Bai, L.; Abdiche, Y.; Dilley, J.; Yu, J.; et al. Proprotein convertase substilisin/kexin type 9 antagonism reduces low-density lipoprotein cholesterol in statin-treated hypercholesterolemic nonhuman primates. J. Pharmacol. Exp. Ther. 2012, 340, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Pokrywka, G.S. PCSK9 inhibitors: A non-statin cholesterol-lowering treatment option. Postgrad. Med. 2018, 130, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Lu, X.; Zhou, H.; Li, R.; Huang, Q.; Xiong, X.; Luo, Z.; Li, W. PCSK9 inhibition protects against myocardial ischemia-reperfusion injury via suppressing autophagy. Microvasc. Res. 2022, 142, 104371. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, M.; Zhang, X.; Liu, C.; Xiang, H.; Huang, M.; Ma, Y.; Gao, X.; Jiang, L.; Liu, X.; et al. The novel llama-human chimeric antibody has potent effect in lowering LDL-c levels in hPCSK9 transgenic rats. Clin. Transl. Med. 2020, 9, 16. [Google Scholar] [CrossRef]
- Trivedi, P.S.; Barouch, L.A. Cardiomyocyte apoptosis in animal models of obesity. Curr. Hypertens. Rep. 2008, 10, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Debin, R.; Lauzier, B.; Sicard, P.; Delemasure, S.; Amoureux, S.; Duvillard, L.; Vergely, C.; Cottin, Y.; Rochette, L. Are Zucker obese rats a useful model for cardiovascular complications in metabolic syndrome? Physical, biochemical and oxidative stress considerations. Fundam. Clin. Pharmacol. 2009, 23, 59–67. [Google Scholar] [CrossRef] [PubMed]
- van Ham, W.B.; Kessler, E.L.; Oerlemans, M.I.; Handoko, M.L.; Sluijter, J.P.; van Veen, T.A.; Ruijter, H.M.D.; de Jager, S.C. Clinical phenotypes of heart failure with preserved ejection fraction to select preclinical animal models. JACC Basic Transl. Sci. 2022, 7, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Folino, A.; Losano, G.; Rastaldo, R. Balance of nitric oxide and reactive oxygen species in myocardial reperfusion injury and protection. J. Cardiovasc. Pharmacol. 2013, 62, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.D.; Stein, E.A.; Hovingh, G.K.; Blom, D.J.; Soran, H.; Watts, G.F.; López, J.A.G.; Bray, S.; Kurtz, C.E.; Hamer, A.W.; et al. Long-term evolocumab in patients with familial hypercholesterolemia. J. Am. Coll. Cardiol. 2020, 75, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Cebova, M.; Rehakova, R.; Kosutova, M.; Pechanova, O. Simvastatin Does Not Affect Nitric Oxide Generation Increased by Sesame Oil in Obese Zucker Rats. Oxidative Med. Cell. Longev. 2018, 2018, 5413423. [Google Scholar] [CrossRef] [PubMed]
- Dayar, E.; Cebova, M.; Lietava, J.; Panghyova, E.; Pechanova, O. Beneficial Effects of Cornelian Cherries on Lipid Profile and NO/ROS Balance in Obese Zucker Rats: Comparison with CoQ10. Molecules 2020, 25, 1922. [Google Scholar] [CrossRef]
- Paulis, L.; Pechanova, O.; Zicha, J.; Krajcirovicova, K.; Barta, A.; Pelouch, V.; Adamcova, M.; Simko, F. Melatonin prevents fibrosis but not hypertrophy development in the left ventricle of NG-nitro-L-arginine-methyl ester hypertensive rats. J. Hypertens. 2009, 27, S11–S16. [Google Scholar] [CrossRef]
- Cebova, M.; Klimentova, J.; Janega, P.; Pechanova, O. Effect of Bioactive Compound of Aronia melanocarpa on Cardiovascular System in Experimental Hypertension. Oxidative Med. Cell. Longev. 2017, 2017, 8156594. [Google Scholar] [CrossRef] [PubMed]
- Mancini, G.J.; Baker, S.; Bergeron, J.; Fitchett, D.; Frohlich, J.; Genest, J.; Gupta, M.; Hegele, R.A.; Ng, D.; Pope, J. Diagnosis, prevention, and management of statin adverse effects and intolerance: Proceedings of a Canadian Working Group Consensus Conference. Can. J. Cardiol. 2011, 27, 635–662. [Google Scholar] [CrossRef]
- Marks, D.; Thorogood, M.; Neil, H.A.W.; Humphries, S.E. A review on the diagnosis, natural history, and treatment of familial hypercholesterolaemia. Atherosclerosis 2003, 168, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.H.; Lebeau, P.F.; Platko, K.; Carlisle, R.E.; Faiyaz, M.; Chen, J.; MacDonald, M.E.; Makda, Y.; Yousof, T.; Lynn, E.G.; et al. Inhibitory Antibodies against PCSK9 Reduce Surface CD36 and Mitigate Diet-Induced Renal Lipotoxicity. Kidney360 2022, 3, 1394–1410. [Google Scholar] [CrossRef]
- Wang, F.; Li, M.; Zhang, A.; Li, H.; Jiang, C.; Guo, J. PCSK9 Modulates Macrophage Polarization-Mediated Ventricular Remodeling after Myocardial Infarction. J. Immunol. Res. 2022, 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sabatine, M.S.; Giugliano, R.P.; Wiviott, S.D.; Raal, F.J.; Blom, D.J.; Robinson, J.; Ballantyne, C.M.; Somaratne, R.; Legg, J.; Wasserman, S.M.; et al. Efficacy and safety of evolocumab in reducing lipids and cardiovascular events. N. Engl. J. Med. 2015, 372, 1500–1509. [Google Scholar] [CrossRef] [PubMed]
- Cammisotto, V.; Baratta, F.; Simeone, P.G.; Barale, C.; Lupia, E.; Galardo, G.; Santilli, F.; Russo, I.; Pignatelli, P. Proprotein Convertase Subtilisin Kexin Type 9 (PCSK9) Beyond Lipids: The Role in Oxidative Stress and Thrombosis. Antioxidants 2022, 11, 569. [Google Scholar] [CrossRef]
- Izkhakov, E.; Shacham, Y.; Serebro, M.; Yaish, I.; Marcus, Y.; Shefer, G.; Tordjman, K.; Greenman, Y.; Stern, N.; Ziv-Baran, T. The Effect of the PCSK9 Inhibitor Evolocumab on Aldosterone Secretion among High Cardiovascular Risk Patients: A Pilot Study. J. Clin. Med. 2021, 10, 2504. [Google Scholar] [CrossRef]
- Ceballos-Macías, J.J.; Lara-Sánchez, C.; Flores-Real, J.; Aguilar-Salinas, C.A.; Ortega-Gutiérrez, G.; Vargas-Sánchez, J.; Madriz-Prado, R.; Derosa, G.; Rodríguez-Benítez, H.; Baltazar-Romero, R.; et al. PCSK-9 Inhibitors in a Real-World Setting and a Comparison Between Alirocumab and Evolocumab in Heterozygous FH Patients. J. Endocr. Soc. 2020, 5, bvaa180. [Google Scholar] [CrossRef] [PubMed]
- Rosenson, R.S.; Daviglus, M.L.; Handelsman, Y.; Pozzilli, P.; Bays, H.; Monsalvo, M.L.; Elliott-Davey, M.; Somaratne, R.; Reaven, P. Efficacy and safety of evolocumab in individuals with type 2 diabetes mellitus: Primary results of the randomised controlled BANTING study. Diabetologia 2019, 62, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Monami, M.; Sesti, G.; Mannucci, E. PCSK9 inhibitor therapy: A systematic review and meta-analysis of metabolic and cardiovascular outcomes in patients with diabetes. Diabetes Obes. Metab. 2019, 21, 903–908. [Google Scholar] [CrossRef]
- Palee, S.; McSweeney, C.M.; Maneechote, C.; Moisescu, D.M.; Jaiwongkam, T.; Kerdphoo, S.; Chattipakorn, S.C.; Chattipakorn, N. PCSK9 inhibitor improves cardiac function and reduces infarct size in rats with ischaemia/reperfusion injury: Benefits beyond lipid-lowering effects. J. Cell. Mol. Med. 2019, 23, 7310–7319. [Google Scholar] [CrossRef]
- Ding, Z.; Liu, S.; Wang, X.; Deng, X.; Fan, Y.; Sun, C.; Wang, Y.; Mehta, J.L. Hemodynamic shear stress via ROS modulates PCSK9 expression in human vascular endothelial and smooth muscle cells and along the mouse aorta. Antioxid. Redox Signal. 2015, 22, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Safaeian, L.; Mirian, M.; Bahrizadeh, S. Evolocumab, a PCSK9 inhibitor, protects human endothelial cells against H2O2-induced oxidative stress. Arch. Physiol. Biochem. 2020, 126, 1681–1686. [Google Scholar] [CrossRef]
- Lankin, V.Z.; Tikhaze, A.K.; Viigimaa, M.; E Chazova, I. PCSK9 Inhibitor causes a decrease in the level of oxidatively modified low-density lipoproteins in patients with coronary artery diseases. Ter. Arkhiv 2018, 90, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ma, X.; Niu, D.; Sun, Y.; Chai, X.; Deng, Y.; Wang, J.; Dong, J. PCSK9 inhibitors suppress oxidative stress and inflammation in atherosclerotic development by promoting macrophage autophagy. Am. J. Transl. Res. 2023, 15, 5129–5144. [Google Scholar] [PubMed]
- Yang, C.L.; Zeng, Y.D.; Hu, Z.X.; Liang, H. PCSK9 promotes the secretion of pro-inflammatory cytokines by macrophages to aggravate H/R-induced cardiomyocyte injury via activating NF-κB signalling. Gen. Physiol. Biophys. 2020, 39, 123–134. [Google Scholar] [CrossRef]
- Shin, D.; Kim, S.; Lee, H.; Lee, H.-C.; Lee, J.; Park, H.-W.; Fukai, M.; Choi, E.; Choi, S.; Koo, B.-J.; et al. PCSK9 stimulates Syk, PKCδ, and NF-κB, leading to atherosclerosis progression independently of LDL receptor. Nat. Commun. 2024, 15, 2789. [Google Scholar] [CrossRef]
- Lei, L.; Li, X.; Yuan, Y.J.; Chen, Z.L.; He, J.H.; Wu, J.H.; Cai, X.S. Inhibition of proprotein convertase subtilisin/kexin type 9 attenuates 2,4,6-trinitrobenzenesulfonic acid-induced colitis via repressing toll-like receptor 4/nuclear factor-kappa B. Kaohsiung J. Med. Sci. 2020, 36, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Abuelezz, S.A.; Hendawy, N. HMGB1/RAGE/TLR4 axis and glutamate as novel targets for PCSK9 inhibitor in high fat cholesterol diet induced cognitive impairment and amyloidosis. Life Sci. 2021, 273, 119310. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yuan, L.; Liu, Z.; Dong, W.; Huang, L.; Liao, A.; Xie, Y.; Liu, R.; Lan, W.; Cai, Y.; et al. Inhibition of PCSK9 Protects against Cerebral Ischemia—Reperfusion Injury via Attenuating Microcirculatory Dysfunction. Neurochem. Res. 2024, 50, 10. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, Y.; Cheng, Z.; Lv, Z.; Luo, S.; Xia, Y. PCSK9 Promotes Endothelial Dysfunction During Sepsis Via the TLR4/MyD88/NF-κB and NLRP3 Pathways. Inflammation 2022, 46, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.-J.; Wu, F.-Y.; Liu, Y.-J.; Li, L.; Lin, Y.-J.; Kang, Y.-T.; Peng, Y.-M.; Liu, Y.-F.; Wang, C.; Ma, Z.-S.; et al. Increase of PCSK9 expression in diabetes promotes VEGFR2 ubiquitination to inhibit endothelial function and skin wound healing. Sci. China Life Sci. 2024, 67, 2635–2649. [Google Scholar] [CrossRef] [PubMed]
- Breder, I.; Breder, J.C.; Bonilha, I.; Munhoz, D.B.; Medorima, S.T.K.; Oliveira, D.C.; Carmo, H.R.D.; Moreira, C.; Kontush, A.; Zimetti, F.; et al. Rationale and design of the expanded combination of evolocumab plus empagliflozin in diabetes: EXCEED-BHS3 trial. Ther. Adv. Chronic Dis. 2020, 11, 2040622320959248. [Google Scholar] [CrossRef] [PubMed]
- Zulkapli, R.; Muid, S.A.; Wang, S.M.; Nawawi, H. PCSK9 Inhibitors Reduce PCSK9 and Early Atherogenic Biomarkers in Stimulated Human Coronary Artery Endothelial Cells. Int. J. Mol. Sci. 2023, 24, 5098. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BW [g] | HW [g] | HW/TL [g/cm] | BP [mmHg] | Glucose [mmol/L] | Fructosamine [μmol/L] | |
|---|---|---|---|---|---|---|
| lean | 383 ± 27 | 1.21 ± 0.09 | 0.325 ± 0.021 | 132 ± 4 | 8.0 ± 0.9 | 177 ± 5 |
| fa/fa | 521 ± 3 2 ** | 1.22 ± 0.03 | 0.351 ± 0.003 | 133 ± 5 | 7.7 ± 0.5 | 190 ± 8 |
| fa/fa+EVO | 508 ± 29 ** | 1.19 ± 0.05 | 0.352 ± 0.015 | 119 ± 8 | 6.9 ± 0.4 | 191 ± 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cebova, M.; Bulkova, R.; Pechanova, O. Evolocumab Reduces Oxidative Stress and Lipid Peroxidation in Obese Zucker Rats. Pathophysiology 2025, 32, 5. https://doi.org/10.3390/pathophysiology32010005
Cebova M, Bulkova R, Pechanova O. Evolocumab Reduces Oxidative Stress and Lipid Peroxidation in Obese Zucker Rats. Pathophysiology. 2025; 32(1):5. https://doi.org/10.3390/pathophysiology32010005
Chicago/Turabian StyleCebova, Martina, Radoslava Bulkova, and Olga Pechanova. 2025. "Evolocumab Reduces Oxidative Stress and Lipid Peroxidation in Obese Zucker Rats" Pathophysiology 32, no. 1: 5. https://doi.org/10.3390/pathophysiology32010005
APA StyleCebova, M., Bulkova, R., & Pechanova, O. (2025). Evolocumab Reduces Oxidative Stress and Lipid Peroxidation in Obese Zucker Rats. Pathophysiology, 32(1), 5. https://doi.org/10.3390/pathophysiology32010005