Dietary Supplementation with Resveratrol Attenuates Serum Melatonin Level, Pro-Inflammatory Response and Metabolic Disorder in Rats Fed High-Fructose High-Lipid Diet under Round-the-Clock Lighting

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Subjects
2.3. Experimental Design
2.4. Biochemical and Enzyme-Linked Immunosorbent Assays
2.5. Statistical Analysis
3. Results
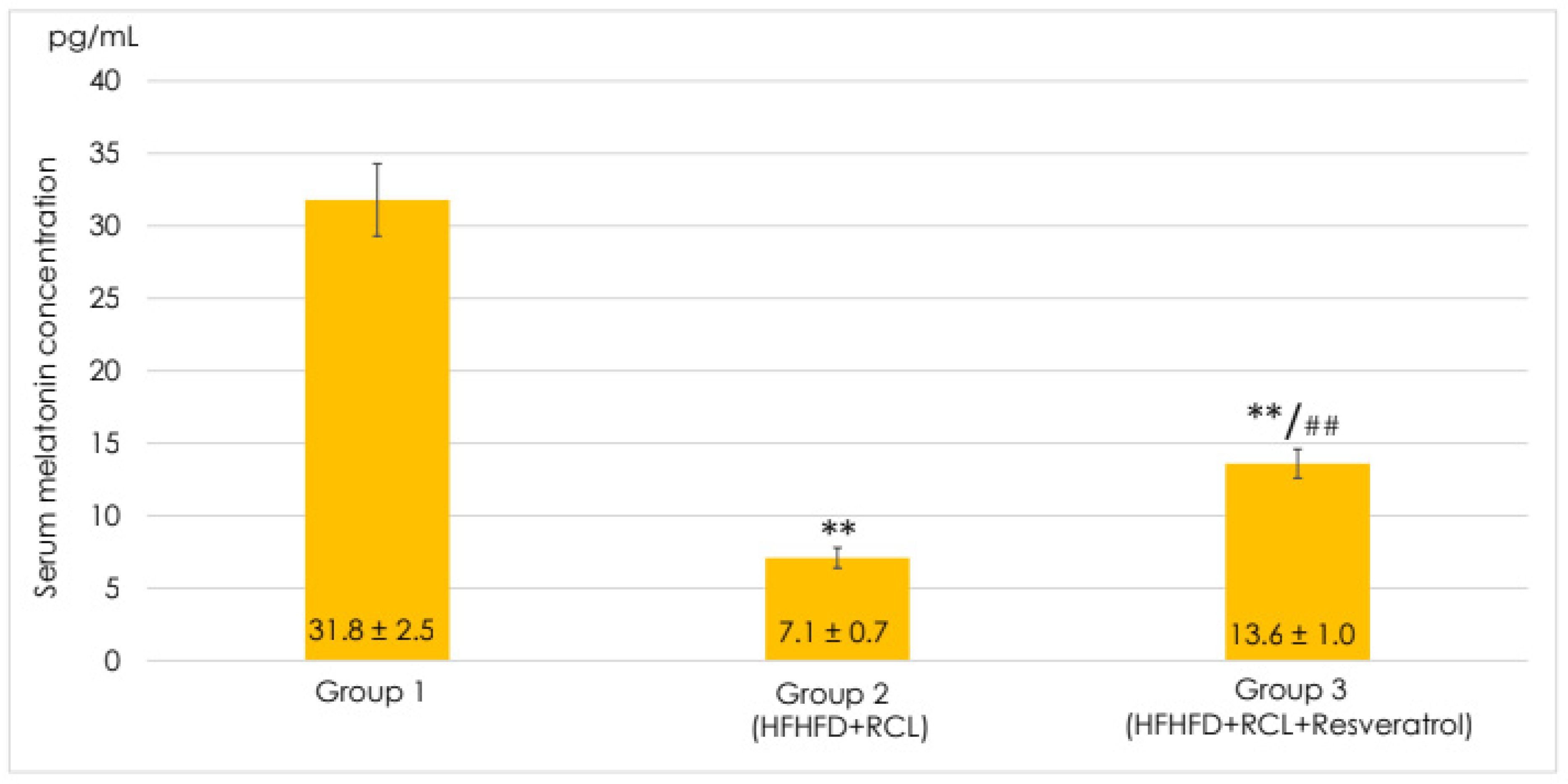
3.1. Effects of Resveratrol on the Melatonin Level in the Serum of the Rats Fed a High-Fructose High-Lipid Diet under Round-the-Clock Lighting
3.2. Effects of Resveratrol on the Systemic Inflammatory Response Indices in the Serum of Rats Fed a High-Fructose High-Lipid Diet under Round-the-Clock Lighting
3.3. Effects of Resveratrol on Lipid Peroxidation in the Blood of Rats Fed a High-Fructose High-Lipid Diet under Round-the-Clock Lighting
3.4. Effects of Resveratrol on Carbohydrate Metabolism in the Serum of Rats Fed a High-Fructose High-Lipid Diet under Round-the-Clock Lighting
3.5. Effects of Resveratrol on Lipid Profile in the Serum of Rats Fed a High-Fructose High-Lipid Diet under Round-the-Clock Lighting
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blume, C.; Garbazza, C.; Spitschan, M. Effects of light on human circadian rhythms, sleep and mood. Somnologie 2019, 23, 147–156. [Google Scholar] [PubMed]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between light at night, melatonin secretion, sleep deprivation, and the internal clock: Health impacts and mechanisms of circadian disruption. Life Sci. 2017, 173, 94–106. [Google Scholar]
- Arango, C.; Buitelaar, J.K.; Fegert, J.M.; Olivier, V.; Pénélaud, P.-F.; Marx, U.; Chimits, D.; Falissard, B.; Barylnik, J.; Birdeanu, L.; et al. Safety and Efficacy of Agomelatine in Children and Adolescents with Major Depressive Disorder Receiving Psychosocial Counselling: A Double-Blind, Randomised, Controlled, Phase 3 Trial in Nine Countries. Lancet Psychiatry 2022, 9, 113–124. [Google Scholar] [PubMed]
- Shkodina, A.D.; Tan, S.C.; Hasan, M.M.; Abdelgawad, M.; Chopra, H.; Bilal, M.; Boiko, D.I.; Tarianyk, K.A.; Alexiou, A. Roles of Clock Genes in the Pathogenesis of Parkinson’s Disease. Ageing Res. Rev. 2022, 74, 101554. [Google Scholar] [PubMed]
- Imenshahidi, M.; Karimi, G.; Hosseinzadeh, H. Effects of melatonin on cardiovascular risk factors and metabolic syndrome: A comprehensive review. NaunynSchmiedebergs Arch. Pharmacol. 2020, 393, 521–536. [Google Scholar]
- Peuhkuri, K.; Sihvola, N.; Korpela, R. Dietary factors and fluctuating levels of melatonin. Food Nutr. Res. 2012, 56. [Google Scholar] [CrossRef]
- Belikova, O.I.; Cherno, V.S.; Frenkel, Y.D.; Kostenko, V.O. Influence of chronic hypomelatoninemia on carbohydrate and lipid metabolism of rats kept on “Western pattern diet”. Fiziol. Zh. 2018, 64, 52–60. [Google Scholar] [CrossRef]
- Belikova, E.I.; Frenkel, Y.D.; Cherno, V.S. Influence of exogenous melatonin on free radical processes in rats exposed to light around the clock under modeling of insulin resistance syndrome. Mod. Probl. Hyg. Radiat. Environ. Med. 2017, 7, 35–51. [Google Scholar]
- Belikova, O.I.; Frenkel, Y.D.; Cherno, V.S.; Kostenko, V.O. Influence of nuclear factor κB inhibitor on biochemical markers of insulin resistance syndrome under hypopinealism and high-calorie carbohydrate-lipid diet. World Med. Biol. 2017, 3, 80–82. [Google Scholar] [CrossRef]
- Frenkel, Y.D.; Cherno, V.S.; Kostenko, V.O. Nrf2 induction alleviates metabolic disorder and systemic inflammatory response in rats under a round-the-clock lighting and high-carbohydrate-lipid diet. Rom. J. Diabetes Nutr. Metab. Dis. 2022, 29, 194–201. [Google Scholar]
- Yelins’ka, A.M.; Shvaykovs’ka, O.O.; Kostenko, V.O. Epigallocatechin-3-gallate prevents disruption of connective tissue in periodontium and salivary glands of rats during systemic inflammation. Wiad. Lek. 2018, 71, 869–873. [Google Scholar]
- Kozaeva, R.; Klymenko, M.O.; Katrushov, O.V.; Kostenko, V.O. Bioflavonoids as agents for correcting nitro-oxidative stress and salivary gland functions in rats exposed to alcohol during modeled lipopolysaccharide-induced systemic inflammatory response. Wiad. Lek. 2022, 75, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Yelins’ka, A.M.; Liashenko, L.I.; Kostenko, V.O. Quercetin potentiates antiradical properties of epigallocatechin-3-gallate in periodontium of rats under systemic and local administration of lipopolisaccharide of Salmonella typhi. Wiad. Lek. 2019, 72, 1499–1503. [Google Scholar] [CrossRef] [PubMed]
- Yavtushenko, I.V.; Nazarenko, S.M.; Katrushov, O.V.; Kostenko, V.O. Quercetin limits the progression of oxidative and nitrosative stress in the rats’ tissues after experimental traumatic brain injury. Wiad. Lek. 2020, 73, 2127–2132. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, Q.; Wang, M.; Liang, M.; Yang, X.; Xu, X.; Zou, H.; Qiu, J. Activation of Sirt1 by resveratrol inhibits TNF-α induced inflammation in fibroblasts. PLoS ONE 2011, 6, e27081. [Google Scholar] [CrossRef] [PubMed]
- Farkhondeh, T.; Folgado, S.L.; Pourbagher-Shahri, A.M.; Ashrafizadeh, M.; Samarghandian, S. The therapeutic effect of resveratrol: Focusing on the Nrf2 signalling pathway. Biomed. Pharmacother. 2020, 127, 110234. [Google Scholar] [CrossRef]
- Chekalina, N.I.; Kazakov, Y.M.; Mamontova, T.V.; Vesnina, L.E.; Kaidashev, I.P. Resveratrol more effectively than quercetin reduces endothelium degeneration and level of necrosis factor α in patients with coronary artery disease. Wiad. Lek. 2016, 69, 475–479. [Google Scholar] [PubMed]
- Akgun-Unal, N.; Ozyildirim, S.; Unal, O.; Gulbahce-Mutlu, E.; Mogulkoc, R.; Baltaci, A.K. The effects of resveratrol and melatonin on biochemical and molecular parameters in diabetic old female rat hearts. Exp. Gerontol. 2023, 172, 112043. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Endale, M.; Wang, W.; Morris, A.R.; Francey, L.J.; Harold, R.L.; Hammers, D.W.; Huo, Z.; Partch, C.L.; Hogenesch, J.B.; et al. NF-κB modifies the mammalian circadian clock through interaction with the core clock protein BMAL1. PLoS Genet. 2021, 17, e1009933. [Google Scholar] [CrossRef] [PubMed]
- Wible, R.S.; Ramanathan, C.; Sutter, C.H.; Olesen, K.M.; Kensler, T.W.; Liu, A.C.; Sutter, T.R. NRF2 regulates core and stabilizing circadian clock loops, coupling redox and timekeeping in Mus musculus. eLife 2018, 7, e31656. [Google Scholar] [CrossRef]
- Mozafari, M.; Nekooeian, A.A.; Panjeshahin, M.R.; Zare, H.R. The effects of resveratrol in rats with simultaneous type 2 diabetes and renal hypertension: A study of antihypertensive mechanisms. Iran J. Med. Sci. 2015, 40, 152–160. [Google Scholar] [PubMed]
- Vesely, O.; Baldovska, S.; Kolesarova, A. Enhancing Bioavailability of Nutraceutically Used Resveratrol and Other Stilbenoids. Nutrients 2021, 13, 3095. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, Y.D.; Zyuzin, V.O.; Cherno, V.S.; Kostenko, V.O. Effect of epigallocatechin-3-gallate and quercetin on the production of reactive oxygen and nitrogen species in liver of rats exposed to round-the-clock light and kept on carbohydrate-lipid diet. Fiziol. Zh. 2022, 68, 20–27. [Google Scholar] [CrossRef]
- Akimov, O.Y.; Kostenko, V.O. Oxidative-Nitrosative Stress and Methods of Its Research; Mahnoliya: Lviv, Ukraine; p. 152p.
- Chao, P.C.; Li, Y.; Chang, C.H.; Shieh, J.P.; Cheng, J.T.; Cheng, K.C. Investigation of insulin resistance in the popularly used four rat models of type-2 diabetes. Biomed. Pharmacother. 2018, 101, 155–161. [Google Scholar] [CrossRef]
- Poulose, N.; Raju, R. Sirtuin regulation in aging and injury. Biochim. Biophys. Acta 2015, 1852, 2442–2455. [Google Scholar] [CrossRef]
- Markus, R.P.; Cecon, E.; Pires-Lapa, M.A. Immune-pineal axis: Nuclear factor κB (NF-kB) mediates the shift in the melatonin source from pinealocytes to immune competent cells. Int. J. Mol. Sci. 2013, 14, 10979–10997. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, W.; Song, G.; Pang, S.; Peng, Z.; Li, Y.; Wang, P. High-Fructose Diet Increases Inflammatory Cytokines and Alters Gut Microbiota Composition in Rats. Mediators Inflamm. 2020, 2020, 6672636. [Google Scholar] [CrossRef]
- Duan, Y.; Zeng, L.; Zheng, C.; Song, B.; Li, F.; Kong, X.; Xu, K. Inflammatory Links Between High Fat Diets and Diseases. Front. Immunol. 2018, 9, 2649. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, D.M.; Gu, T.T.; Ding, X.Q.; Fan, C.Y.; Zhu, Q.; Shi, Y.W.; Hong, Y.; Kong, L.D. Morin reduces hepatic inflammation-associated lipid accumulation in high fructose-fed rats via inhibiting sphingosine kinase 1/sphingosine 1-phosphate signalling pathway. Biochem. Pharmacol. 2013, 86, 1791–1804. [Google Scholar] [CrossRef]
- Vieira, E.; Mirizio, G.G.; Barin, G.R.; de Andrade, R.V.; Nimer, N.F.S.; La Sala, L. Clock Genes, Inflammation and the Immune System-Implications for Diabetes, Obesity and Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 9743. [Google Scholar] [CrossRef]
- Ojetola, A.A.; Asiwe, J.N.; Adeyemi, W.J.; Ogundipe, D.J.; Fasanmade, A.A. Dietary Supplementation with D-Ribose-L-Cysteine Prevents Hepatic Stress and Pro-Inflammatory Responses in Male Wistar Rats Fed a High-Fructose High-Fat Diet. Pathophysiology 2022, 29, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Boiko, D.I.; Shkodina, A.D.; Hasan, M.M.; Bardhan, M.; Kazmi, S.K.; Chopra, H.; Bhutra, P.; Baig, A.A.; Skrypnikov, A.M. Melatonergic Receptors (Mt1/Mt2) as a Potential Additional Target of Novel Drugs for Depression. Neurochem. Res. 2022, 47, 2909–2924. [Google Scholar] [CrossRef] [PubMed]
- Koushki, M.; Dashatan, N.A.; Meshkani, R. Effect of Resveratrol Supplementation on Inflammatory Markers: A Systematic Review and Meta-analysis of Randomized Controlled Trials. Clin. Ther. 2018, 40, 1180–1192.e5. [Google Scholar] [CrossRef] [PubMed]
- Kopec, A.; Piatkowska, E.; Leszczynska, T.; Koronowicz, A. Effect of long term administration of resveratrol on lipid concentration in selected organs and liver’s histology in rats fed high fructose diet. J. Funct. Foods 2013, 5, 299–305. [Google Scholar] [CrossRef]
- Reda, D.; Elshopakey, G.E.; Mahgoub, H.A.; Risha, E.F.; Khan, A.A.; Rajab, B.S.; El-Boshy, M.E.; Abdelhamid, F.M. Effects of Resveratrol Against Induced Metabolic Syndrome in Rats: Role of Oxidative Stress, Inflammation, and Insulin Resistance. Evid. Based Complement. Alternat. Med. 2022, 2022, 3362005. [Google Scholar] [CrossRef] [PubMed]
- Kuršvietienė, L.; Stanevičienė, I.; Mongirdienė, A.; Bernatonienė, J. Multiplicity of effects and health benefits of resveratrol. Medicina 2016, 52, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Javkhedkar, A.A.; Quiroz, Y.; Rodriguez-Iturbe, B.; Vaziri, N.D.; Lokhandwala, M.F.; Banday, A.A. Resveratrol restored Nrf2 function, reduced renal inflammation, and mitigated hypertension in spontaneously hypertensive rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R840–R846. [Google Scholar] [CrossRef]
- Kuznetsova, T.Y.; Solovyova, N.V.; Solovyov, V.V.; Kostenko, V.O. Antioxidant activity of melatonin and glutathione interacting with hydroxyl and superoxide anion radicals. Ukr. Biochem. J. 2017, 89, 22–30. [Google Scholar] [CrossRef]
- Abadi, S.H.M.H.; Shirazi, A.; Alizadeh, A.M.; Changizi, V.; Najafi, M.; Khalighfard, S.; Nosrati, H. The Effect of Melatonin on Superoxide Dismutase and Glutathione Peroxidase Activity, and Malondialdehyde Levels in the Targeted and the Non-targetedLung and Heart Tissues after Irradiation in Xenograft Mice Colon Cancer. Curr. Mol. Pharmacol. 2018, 11, 326–335. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Y.; Song, Y.; Cheng, X.R.; Xia, S.; Rahman, M.R.; Shi, Y.; Le, G. Resveratrol restores the circadian rhythmic disorder of lipid metabolism induced by high-fat diet in mice. Biochem. Biophys. Res. Commun. 2015, 458, 86–91. [Google Scholar] [CrossRef]


{kind=link}
| Standard Rat Chow | High-Lipid Diet | ||
|---|---|---|---|
| Nutrients | g/kg Total | Nutrients | g/kg Total |
| Protein | 160 | Protein | 94 |
| Fat | 70 | Fat | 313 |
| Carbohydrate | 480 | Carbohydrate | 466 |
| Fibre | 68 | Fibre | 14 |
| Sodium | 2.7 | Sodium | 4 |
| Vitaminand mineral mix | 36 | Vitaminand mineral mix | 40 |
| Ingredients: | |||
| Refined wheat flour | 450 | ||
| Skimmed milk powder | 200 | ||
| Table margarine 82% fat | 200 | ||
| Starch | 100 | ||
| Peroxidised sunflower oil | 40 | ||
| Sodium chloride | 10 | ||
| Total calorie | 2720 kcal/kg | Total calorie | 4477 kcal/kg |
| Groups/Parameters | TNF-α (pg/mL) | CRP (ng/mL) |
|---|---|---|
| 1. Group 1 (Control), n = 7 | 34.0 ± 2.0 | 4.1 ± 0.1 |
| 2. Group 2 (HFHLD + RCL), n = 7 | 109.8 ± 6.0 ** | 12.8 ± 0.3 ** |
| 3. Group 3 (HFHLD + RCL + Resveratrol), n = 7 | 43.6 ± 4.9 ## | 5.3 ± 0.3 *,## |
| Groups/Parameters | MDA-TBA2 (µmol/L) | ||
|---|---|---|---|
| Before Incubation | After Incubation | Increment over Incubation Time | |
| 1. Group 1 (Control), n = 7 | 11.2 ± 0.9 | 25.4 ± 2.0 | 14.2 ± 2.4 |
| 2. Group 2 (HFHLD + RCL), n = 7 | 23.0 ± 0.6 ** | 48.3 ± 2.1 ** | 25.3 ± 1.9 * |
| 3. Group 3 (HFHLD + RCL + Resveratrol), n = 7 | 12.6 ± 0.9 ## | 27.7 ± 1.9 ## | 15.1 ± 2.2 # |
| Groups/Parameters | Glucose (mmol/L) | Insulin (μU/mL) | HOMA-IR |
|---|---|---|---|
| 1. Group 1 (Control), n = 7 | 4.94 ± 0.24 | 1.5 ± 0.2 | 0.32 ± 0.06 |
| 2. Group 2 (HFHLD + RCL), n = 7 | 6.89 ± 0.25 * | 5.5 ± 0.2 ** | 1.17 ± 0.04 ** |
| 3. Group 3 (HFHLD + RCL + Resveratrol), n = 7 | 4.45 ± 0.18 # | 2.1 ± 0.1 ## | 0.42 ± 0.03 ## |
| Groups/Parameters | Total CH (mmol/L) | HDL (mmol/L) | LDL (mmol/L) | VLDL (mmol/L) | TAG (mmol/L) |
|---|---|---|---|---|---|
| 1. Group 1 (Control), n = 7 | 2.39 ± 0.29 | 0.63 ± 0.04 | 1.47 ± 0.29 | 0.29 ± 0.02 | 0.65 ± 0.05 |
| 2. Group 2 (HFHLD + RCL), n = 7 | 2.62 ± 0.30 | 0.23 ± 0.02 ** | 1.43 ± 0.29 | 0.96 ± 0.04 ** | 2.10 ± 0.10 ** |
| 3. Group 3 (HFHLD + RCL + Resveratrol), n = 7 | 2.28 ± 0.22 | 0.48 ± 0.02 *,# | 1.29 ± 0.22 | 0.51 ± 0.03 **,## | 1.11 ± 0.06 **,## |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frenkel, Y.; Cherno, V.; Kostenko, H.; Chopra, H.; Gautam, R.K.; Kostenko, V. Dietary Supplementation with Resveratrol Attenuates Serum Melatonin Level, Pro-Inflammatory Response and Metabolic Disorder in Rats Fed High-Fructose High-Lipid Diet under Round-the-Clock Lighting. Pathophysiology 2023, 30, 37-47. https://doi.org/10.3390/pathophysiology30010005
Frenkel Y, Cherno V, Kostenko H, Chopra H, Gautam RK, Kostenko V. Dietary Supplementation with Resveratrol Attenuates Serum Melatonin Level, Pro-Inflammatory Response and Metabolic Disorder in Rats Fed High-Fructose High-Lipid Diet under Round-the-Clock Lighting. Pathophysiology. 2023; 30(1):37-47. https://doi.org/10.3390/pathophysiology30010005
Chicago/Turabian StyleFrenkel, Yurii, Valerii Cherno, Heorhii Kostenko, Hitesh Chopra, Rupesh K. Gautam, and Vitalii Kostenko. 2023. "Dietary Supplementation with Resveratrol Attenuates Serum Melatonin Level, Pro-Inflammatory Response and Metabolic Disorder in Rats Fed High-Fructose High-Lipid Diet under Round-the-Clock Lighting" Pathophysiology 30, no. 1: 37-47. https://doi.org/10.3390/pathophysiology30010005
APA StyleFrenkel, Y., Cherno, V., Kostenko, H., Chopra, H., Gautam, R. K., & Kostenko, V. (2023). Dietary Supplementation with Resveratrol Attenuates Serum Melatonin Level, Pro-Inflammatory Response and Metabolic Disorder in Rats Fed High-Fructose High-Lipid Diet under Round-the-Clock Lighting. Pathophysiology, 30(1), 37-47. https://doi.org/10.3390/pathophysiology30010005