Effects of Zearalenone on Production Performance, Egg Quality, Ovarian Function and Gut Microbiota of Laying Hens


Abstract
:1. Introduction
2. Results
2.1. Effects of ZEN on the Production Performance of Laying Hens
2.2. Effects of ZEN on the Egg Quality of Laying Hens
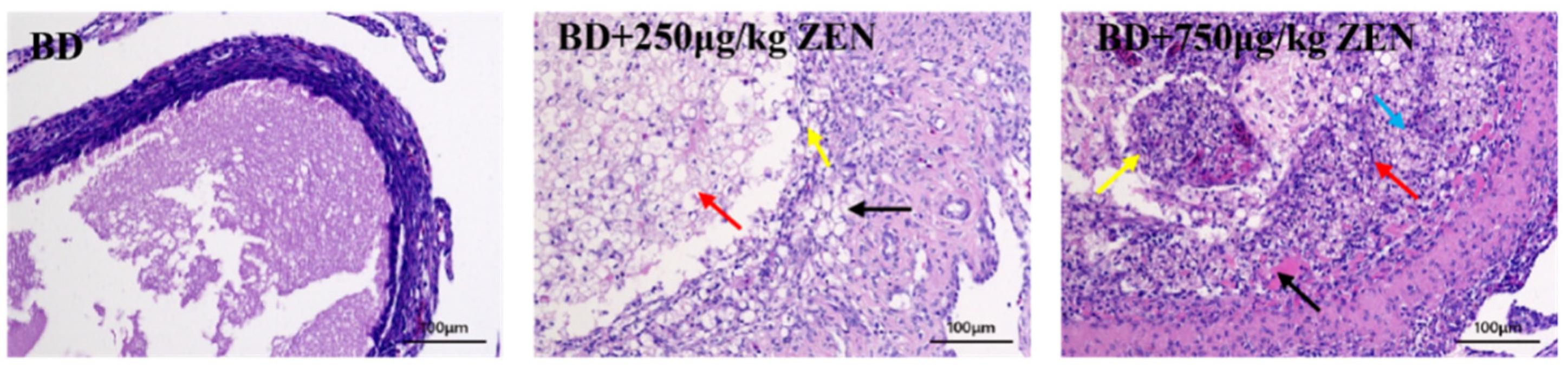
2.3. Effects of ZEN on the Ovarian Function of Laying Hens
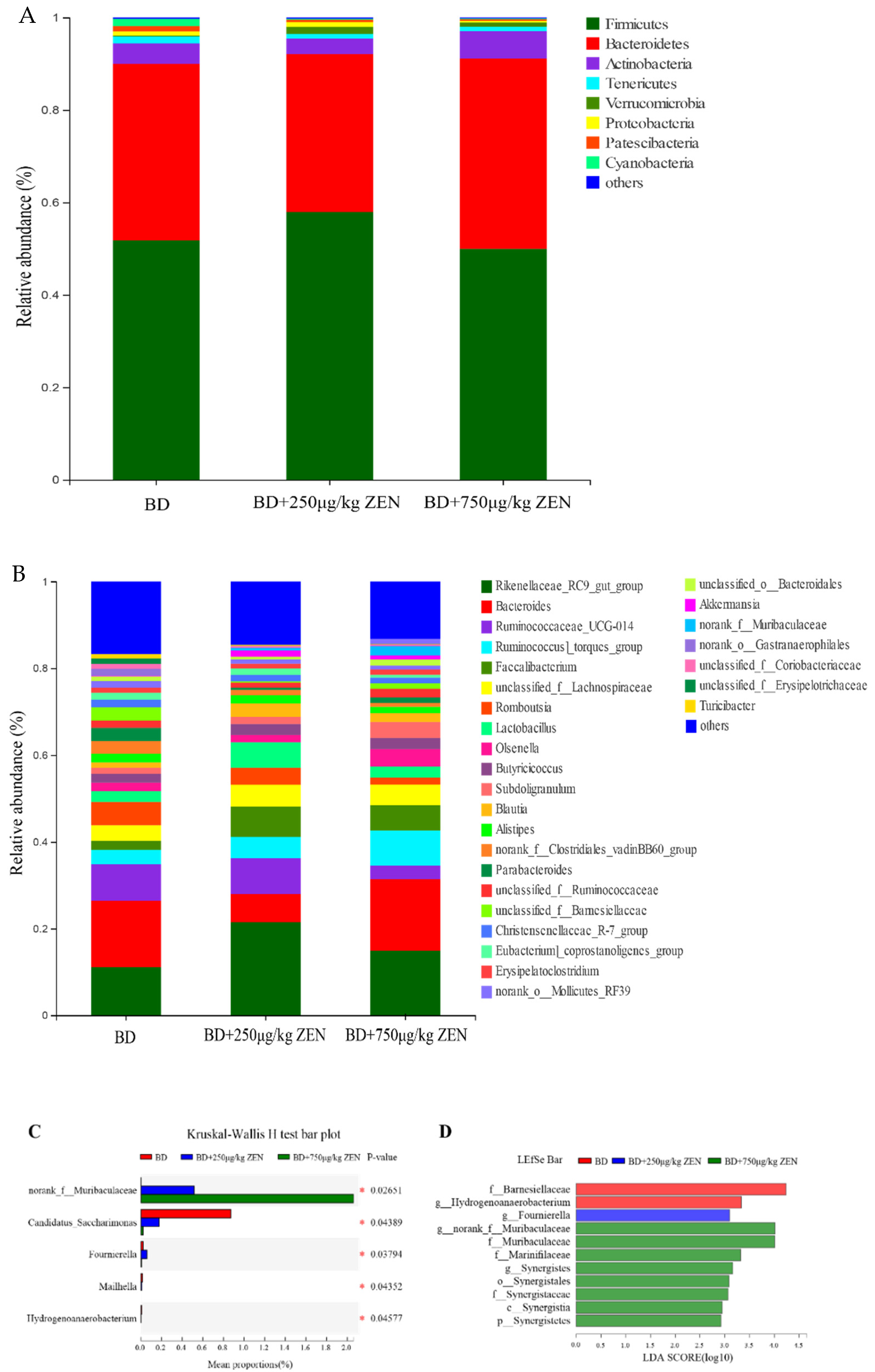
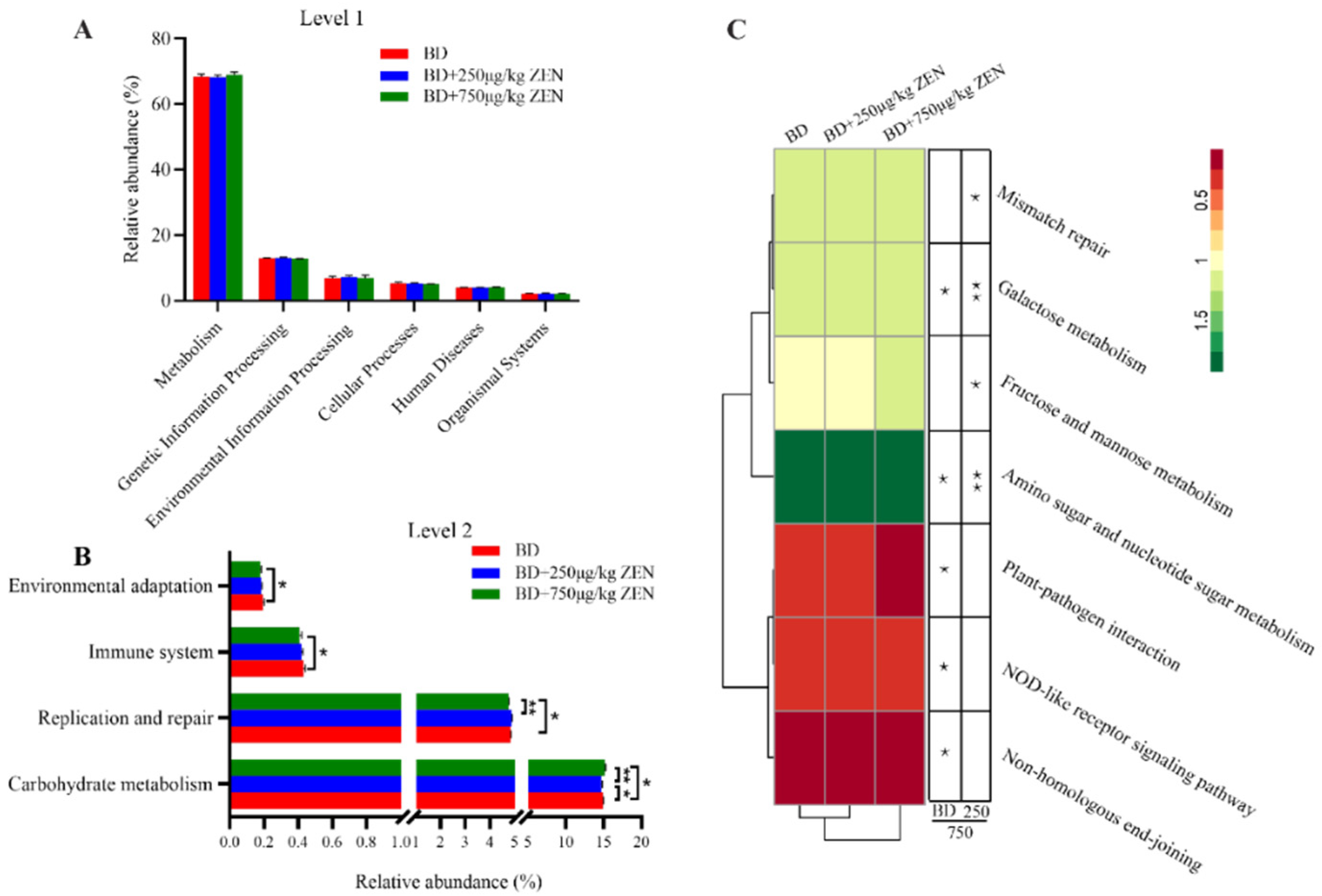
2.4. Effects of ZEN on the Gut Microbiota of Laying Hens
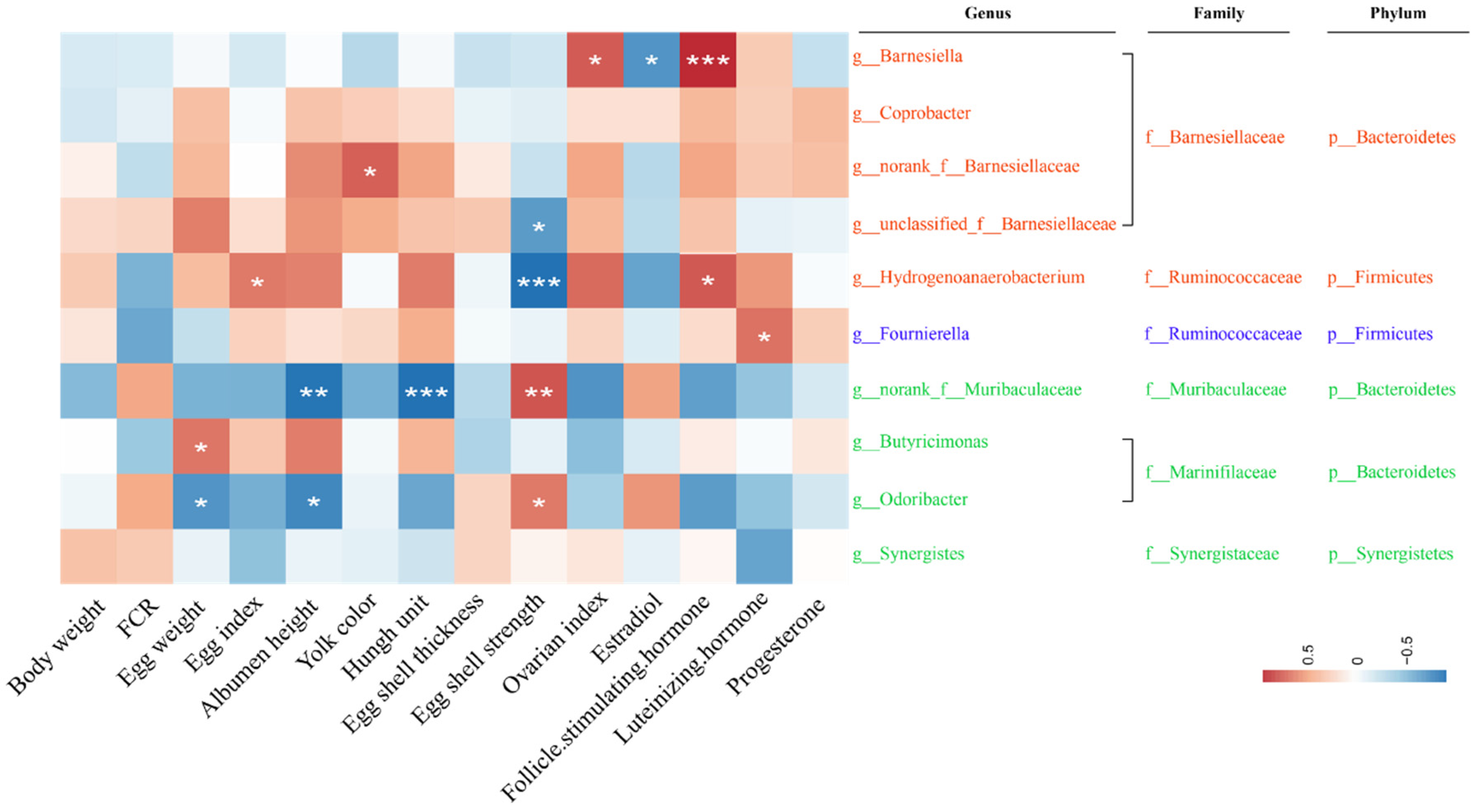
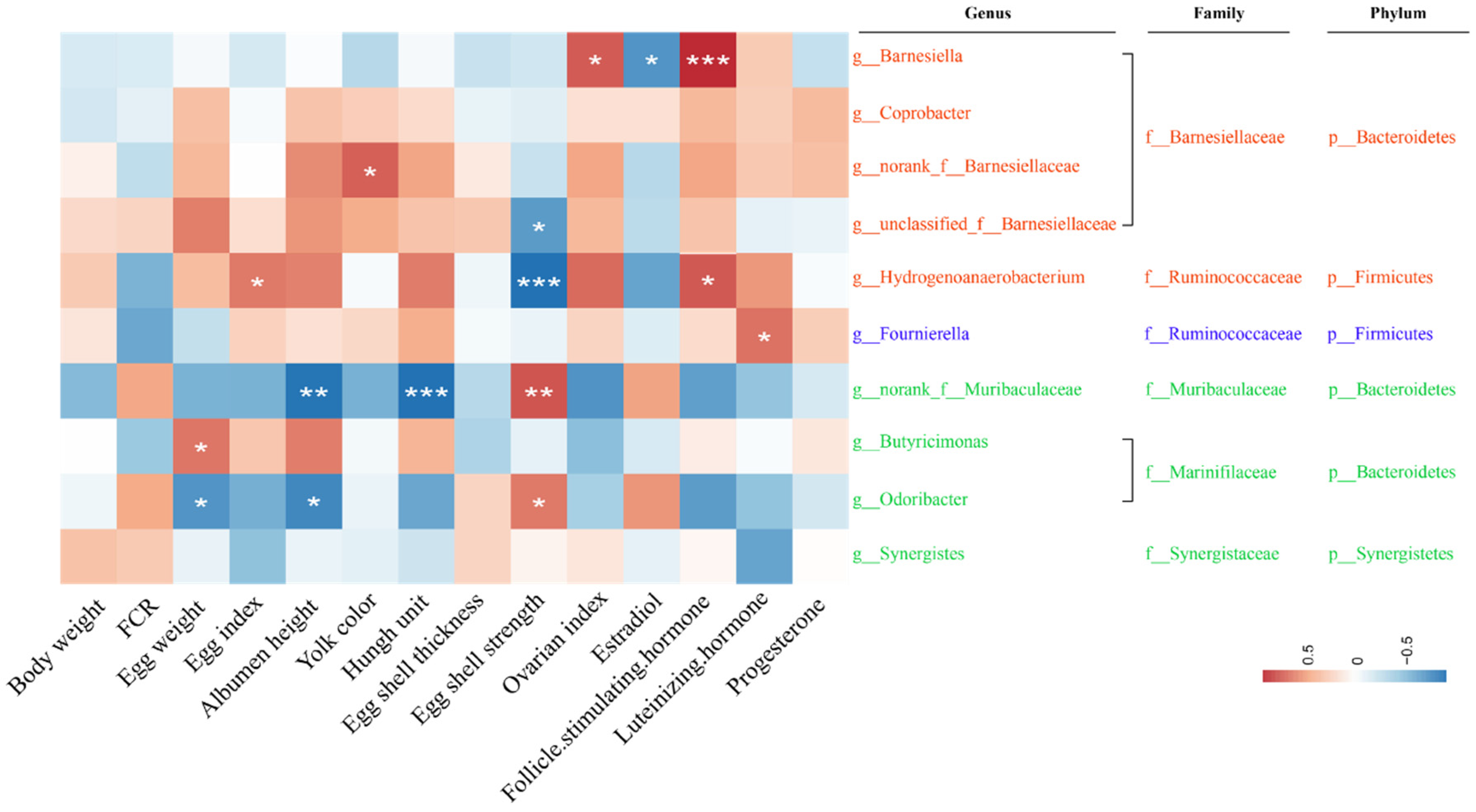
2.5. Correlation between Gut Microbiota and Reproductive Performance and Ovarian Function Faliure of Laying Hens
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals, Experimental Design and Sample Collection
5.2. Production Performance and Organ Index Analysis
5.3. Egg Quality Analysis
5.4. Serum Reproductive Hormones Level Analysis
5.5. Morphology Analysis
5.6. S rRNA Sequencing and Bioinformatics Analyses
5.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, Y.; Tan, Y.; Yan, Z.; Liao, Y.; Chen, J.; De Boevre, M.; De Saeger, S.; Wu, A. Antagonistic and Detoxification Potentials of Trichoderma Isolates for Control of Zearalenone (ZEN) Producing Fusarium graminearum. Front. Microbiol. 2017, 8, 2710. [Google Scholar] [CrossRef] [PubMed]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Monbaliu, S.; Van Poucke, C.; Detavernier, C.; Dumoulin, F.; Van De Velde, M.; Schoeters, E.; Van Dyck, S.; Averkieva, O.; Van Peteghem, C.; De Saeger, S. Occurrence of mycotoxins in feed as analyzed by a multi-mycotoxin LC-MS/MS method. J. Agric. Food Chem. 2010, 58, 66–71. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Zheng, H.; He, C.; Zhang, H. T-2 toxin, zearalenone and fumonisin B1 in feedstuffs from China. Food Addit. Contam. Part B 2013, 6, 116–122. [Google Scholar] [CrossRef]
- Kemboi, D.C.; Ochieng, P.E.; Antonissen, G.; Croubels, S.; Scippo, M.L.; Okoth, S.; Kangethe, E.K.; Faas, J.; Doupovec, B.; Lindahl, J.F.; et al. Multi-Mycotoxin Occurrence in Dairy Cattle and Poultry Feeds and Feed Ingredients from Machakos Town, Kenya. Toxins 2020, 12, 762. [Google Scholar] [CrossRef] [PubMed]
- Salah-Abbès, J.B.; Abbès, S.; Ouanes, Z.; Houas, Z.; Abdel-Wahhab, M.A.; Bacha, H.; Oueslati, R. Tunisian radish extract (Raphanus sativus) enhances the antioxidant status and protects against oxidative stress induced by zearalenone in Balb/c mice. J. Appl. Toxicol. 2008, 28, 6–14. [Google Scholar] [CrossRef]
- Zheng, W.; Feng, N.; Wang, Y.; Noll, L.; Xu, S.; Liu, X.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; et al. Effects of zearalenone and its derivatives on the synthesis and secretion of mammalian sex steroid hormones: A review. Food Chem. Toxicol. 2019, 126, 262–276. [Google Scholar] [CrossRef]
- Yang, D.; Jiang, X.; Sun, J.; Li, X.; Li, X.; Jiao, R.; Peng, Z.; Li, Y.; Bai, W. Toxic effects of zearalenone on gametogenesis and embryonic development: A molecular point of review. Food Chem. Toxicol. 2018, 119, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Andersen, C.L.; Hu, L.; Wang, Z.; Li, Y.; Nagy, T.; Ye, X. Dietary exposure to mycotoxin zearalenone (ZEA) during post-implantation adversely affects placental development in mice. Reprod. Toxicol. 2019, 85, 42–50. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Y.; Gu, J.; Yuan, Y.; Liu, X.; Zheng, W.; Huang, Q.; Liu, Z.; Bian, J. Zearalenone inhibits testosterone biosynthesis in mouse Leydig cells via the crosstalk of estrogen receptor signaling and orphan nuclear receptor Nur77 expression. Toxicol. Vitr. 2014, 28, 647–656. [Google Scholar] [CrossRef]
- Massart, F.; Saggese, G. Oestrogenic mycotoxin exposures and precocious pubertal development. Int. J. Androl. 2010, 33, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Zatecka, E.; Ded, L.; Elzeinova, F.; Kubatova, A.; Dorosh, A.; Margaryan, H.; Dostalova, P.; Korenkova, V.; Hoskova, K.; Peknicova, J. Effect of zearalenone on reproductive parameters and expression of selected testicular genes in mice. Reprod. Toxicol. (Elmsford N. Y.) 2014, 45, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, Ł.; Gajęcka, M.; Lisieska-Żołnierczyk, S.; Dąbrowski, M.; Gajęcki, M.T. The Effect of Different Doses of Zearalenone in Feed on the Bioavailability of Zearalenone and Alpha-Zearalenol, and the Concentrations of Estradiol and Testosterone in the Peripheral Blood of Pre-Pubertal Gilts. Toxins 2020, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, F.; Caloni, F.; Schreiber, N.B.; Cortinovis, C.; Spicer, L.J. In vitro effects of deoxynivalenol and zearalenone major metabolites alone and combined, on cell proliferation, steroid production and gene expression in bovine small-follicle granulosa cells. Toxicon 2016, 109, 70–83. [Google Scholar] [CrossRef]
- Silva, I.P.; Brito, D.C.C.; Silva, T.E.S.; Silva, R.F.; Guedes, M.I.F.; Silva, J.Y.G.; Rodrigues, A.P.R.; Santos, R.R.; Figueiredo, J.R. In vitro exposure of sheep ovarian tissue to the xenoestrogens zearalenone and enterolactone: Effects on preantral follicles. Theriogenology 2021, 174, 124–130. [Google Scholar] [CrossRef]
- Chang, J.; Wang, T.; Wang, P.; Yin, Q.; Liu, C.; Zhu, Q.; Lu, F.; Gao, T. Compound probiotics alleviating aflatoxin B1 and zearalenone toxic effects on broiler production performance and gut microbiota. Ecotoxicol. Environ. Saf. 2020, 194, 110420. [Google Scholar] [CrossRef]
- Wu, K.; Ren, C.; Gong, Y.; Gao, X.; Rajput, S.A.; Qi, D.; Wang, S. The insensitive mechanism of poultry to zearalenone: A review. Anim. Nutr. 2021, 7, 587–594. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, C.; Wang, J.; Celi, P.; Ding, X.; Bai, S.; Zeng, Q.; Mao, X.; Zhuo, Y.; Xu, S.; et al. Characterization of the Intestinal Microbiota of Broiler Breeders With Different Egg Laying Rate. Front. Vet. Sci. 2020, 7, 599337. [Google Scholar] [CrossRef]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen-gut microbiome axis: Physiological and clinical implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef]
- Liew, W.P.; Mohd-Redzwan, S. Mycotoxin: Its Impact on Gut Health and Microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Elmassry, M.M.; Zayed, A.; Farag, M.A. Gut homeostasis and microbiota under attack: Impact of the different types of food contaminants on gut health. Crit. Rev. Food Sci. Nutr. 2022, 62, 738–763. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2020, 60, 2710–2729. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef]
- Sypecka, Z.; Kelly, M.; Brereton, P. Deoxynivalenol and zearalenone residues in eggs of laying hens fed with a naturally contaminated diet: Effects on egg production and estimation of transmission rates from feed to eggs. J. Agric. Food Chem. 2004, 52, 5463–5471. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Ueberschär, K.H.; Halle, I.; Matthes, S.; Valenta, H.; Flachowsky, G. Effect of addition of a detoxifying agent to laying hen diets containing uncontaminated or Fusarium toxin-contaminated maize on performance of hens and on carryover of zearalenone. Poult. Sci. 2002, 81, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.R.; Smith, T.K. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on performance and metabolism of laying hens. Poult. Sci. 2004, 83, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Bari, M.S.; Cohen-Barnhouse, A.M.; Campbell, D.L.M. Early rearing enrichments influenced nest use and egg quality in free-range laying hens. Animal 2020, 14, 1249–1257. [Google Scholar] [CrossRef]
- Narinc, D.; Aygun, A.; Karaman, E.; Aksoy, T. Egg shell quality in Japanese quail: Characteristics, heritabilities and genetic and phenotypic relationships. Animal 2015, 9, 1091–1096. [Google Scholar] [CrossRef]
- Tomczyk, Ł.; Szablewski, T.; Stuper-Szablewska, K.; Nowaczewski, S.; Cegielska-Radziejewska, R. The influence of the conditions of acquisition and storage of table eggs on changes in their quality and the presence of mycobiota and Fusarium mycotoxins. Poult. Sci. 2019, 98, 2964–2971. [Google Scholar] [CrossRef]
- Zhao, L.; Feng, Y.; Wei, J.T.; Zhu, M.X.; Zhang, L.; Zhang, J.C.; Karrow, N.A.; Han, Y.M. Mitigation Effects of Bentonite and Yeast Cell Wall Binders on AFB(1), DON, and OTA Induced Changes in Laying Hen Performance, Egg Quality, and Health. Toxins 2021, 13, 156. [Google Scholar] [CrossRef]
- Jia, R.; Ma, Q.; Fan, Y.; Ji, C.; Zhang, J.; Liu, T.; Zhao, L. The toxic effects of combined aflatoxins and zearalenone in naturally contaminated diets on laying performance, egg quality and mycotoxins residues in eggs of layers and the protective effect of Bacillus subtilis biodegradation product. Food Chem. Toxicol. 2016, 90, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Ueberschär, K.H.; Halle, I.; Matthes, S.; Valenta, H.; Flachowsky, G. On the effects of Fusarium toxin contaminated maize in laying hen feeding. Mycotoxin Res. 2001, 17 (Suppl. 1), 62–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Applegate, T. Zearalenone (ZEN) in Livestock and Poultry: Dose, Toxicokinetics, Toxicity and Estrogenicity. Toxins 2020, 12, 377. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed. Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Ratriyanto, A.; Mosenthin, R. Osmoregulatory function of betaine in alleviating heat stress in poultry. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1634–1650. [Google Scholar] [CrossRef]
- Chen, X.X.; Yang, C.W.; Huang, L.B.; Niu, Q.S.; Jiang, S.Z.; Chi, F. Zearalenone Altered the Serum Hormones, Morphologic and Apoptotic Measurements of Genital Organs in Post-weaning Gilts. Asian-Australas. J. Anim. Sci. 2015, 28, 171–179. [Google Scholar] [CrossRef]
- Zhao, L.; Lei, Y.; Bao, Y.; Jia, R.; Ma, Q.; Zhang, J.; Chen, J.; Ji, C. Ameliorative effects of Bacillus subtilis ANSB01G on zearalenone toxicosis in pre-pubertal female gilts. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2015, 32, 617–625. [Google Scholar] [CrossRef]
- Gao, X.; Xiao, Z.H.; Liu, M.; Zhang, N.Y.; Khalil, M.M.; Gu, C.Q.; Qi, D.S.; Sun, L.H. Dietary Silymarin Supplementation Alleviates Zearalenone-Induced Hepatotoxicity and Reproductive Toxicity in Rats. J. Nutr. 2018, 148, 1209–1216. [Google Scholar] [CrossRef]
- Dong, M.; He, X.J.; Tulayakul, P.; Li, J.Y.; Dong, K.S.; Manabe, N.; Nakayama, H.; Kumagai, S. The toxic effects and fate of intravenously administered zearalenone in goats. Toxicon 2010, 55, 523–530. [Google Scholar] [CrossRef]
- Toyama, R.; Fujimori, T.; Nabeshima, Y.; Itoh, Y.; Tsuji, Y.; Osamura, R.Y.; Nabeshima, Y. Impaired regulation of gonadotropins leads to the atrophy of the female reproductive system in klotho-deficient mice. Endocrinology 2006, 147, 120–129. [Google Scholar] [CrossRef]
- Schulster, M.; Bernie, A.M.; Ramasamy, R. The role of estradiol in male reproductive function. Asian J. Androl. 2016, 18, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Wasser, S.K.; Fujimoto, V.Y.; Klein, N.A.; Moore, D.E.; Soules, M.R. Utility of follicle stimulating hormone (FSH), luteinizing hormone (LH), oestradiol and FSH:LH ratio in predicting reproductive age in normal women. Hum. Reprod. (Oxf. Engl.) 1997, 12, 1152–1155. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.F.; Zhang, N.Y.; Peng, Y.Z.; Qi, D.S. Interaction of zearalenone and soybean isoflavone on the development of reproductive organs, reproductive hormones and estrogen receptor expression in prepubertal gilts. Anim. Reprod. Sci. 2010, 122, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Yang, S.; Zhang, W.; Zhang, Y.; Li, P.; Guo, Y.; Wang, Y.; Chen, X.; He, J. The Influence of Selenium Yeast on Hematological, Biochemical and Reproductive Hormone Level Changes in Kunming Mice Following Acute Exposure to Zearalenone. Biol. Trace Elem. Res. 2016, 174, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Cui, J.; Yang, X.; Liu, S.; Han, S.; Chen, B. Effects of zearalenone on genital organ development, serum immunoglobulin, antioxidant capacity, sex hormones and liver function of prepubertal gilts. Toxicon 2021, 189, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.F.; Sprando, R.L.; Black, T.N.; Olejnik, N.; Eppley, R.M.; Alam, H.Z.; Rorie, J.; Ruggles, D.I. Effects of zearalenone on in utero development in rats. Food Chem. Toxicol. 2006, 44, 1455–1465. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Piotrowska, M.; Sliżewska, K.; Nowak, A.; Zielonka, L.; Zakowska, Z.; Gajęcka, M.; Gajęcki, M. The effect of experimental fusarium mycotoxicosis on microbiota diversity in porcine ascending colon contents. Toxins 2014, 6, 2064–2081. [Google Scholar] [CrossRef]
- Hussain, T.; Murtaza, G.; Kalhoro, D.H.; Kalhoro, M.S.; Metwally, E.; Chughtai, M.I.; Mazhar, M.U.; Khan, S.A. Relationship between gut microbiota and host-metabolism: Emphasis on hormones related to reproductive function. Anim. Nutr. 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, B.; Sun, Y.; Du, T.; Chen, Y.; Wang, X.; Li, H.; Nan, Y.; Zhang, G.; Zhou, E.M. Decreased egg production in laying hens associated with infection with genotype 3 avian hepatitis E virus strain from China. Vet. Microbiol. 2017, 203, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.R.; Smith, T.K.; Boermans, H.J.; Woodward, B. Effects of feed-borne Fusarium mycotoxins on hematology and immunology of laying hens. Poult. Sci. 2005, 84, 1841–1850. [Google Scholar] [CrossRef] [PubMed]
- Martins, H.M.; Guerra, M.M.; Bernardo, F. Zearalenone, deoxynivalenol and fumonisins in mixed-feed for laying hens. Mycotoxin Res. 2006, 22, 206–210. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, L.; Xu, Z.; Liu, X.; Chen, L.; Dai, J.; Karrow, N.A.; Sun, L. Occurrence of Aflatoxin B(1), deoxynivalenol and zearalenone in feeds in China during 2018–2020. J. Anim. Sci. Biotechnol. 2021, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Mokubedi, S.M.; Phoku, J.Z.; Changwa, R.N.; Gbashi, S.; Njobeh, P.B. Analysis of Mycotoxins Contamination in Poultry Feeds Manufactured in Selected Provinces of South Africa Using UHPLC-MS/MS. Toxins 2019, 11, 452. [Google Scholar] [CrossRef]
- Gou, Z.; Fan, Q.; Li, L.; Jiang, Z.; Lin, X.; Cui, X.; Wang, Y.; Zheng, C.; Jiang, S. Effects of dietary iron on reproductive performance of Chinese Yellow broiler breeder hens during the egg-laying period. Poult. Sci. 2020, 99, 3921–3929. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Li, X.; Xi, C.; Tang, B.; Wang, G.; Zhang, L.; Yuan, Z.; Zhao, H. Simultaneous determination of residues of six zeranols in eggs by high performance liquid chromatography with immunoaffinity cleanup column. Se Pu 2012, 30, 1021–1025. [Google Scholar] [CrossRef]
- Zhu, L.; Fu, J.; Xiao, X.; Wang, F.; Jin, M.; Fang, W.; Wang, Y. Faecal microbiota transplantation-mediated jejunal microbiota changes halt high-fat diet-induced obesity in mice via retarding intestinal fat absorption. Microb. Biotechnol. 2022, 15, 337–352. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week | Items | Supplemental Levels of ZEN in Diets, μg/kg | ||
|---|---|---|---|---|
| BD | BD + 250 μg/kg ZEN | BD + 750 μg/kg ZEN | ||
| Week 2 | Egg production (%) | 98.6 ± 2.0 | 98.6 ± 1.8 | 96.4 ± 3.2 |
| Feed intake (%) | 109.9 ± 1.3 | 110.2 ± 1.7 | 109.8 ± 1.3 | |
| FCR (g feed/g egg) | 1.86 ± 0.09 b | 1.88 ± 0.04 b | 1.99 ± 0.15 a | |
| Week 5 | Egg production (%) | 98.3 ± 1.6 | 98.5 ± 1.0 | 96.6 ± 2.0 |
| Feed intake (%) | 109.2 ± 1.1 | 109.4 ± 2.1 | 109.7 ± 0.8 | |
| FCR (g feed/g egg) | 1.83 ± 0.07 b | 1.86 ± 0.04 ab | 1.91 ± 0.08 a | |
| Week | Items | Supplemental Levels of ZEN in Diets, μg/kg | ||
|---|---|---|---|---|
| BD | BD + 250 μg/kg ZEN | BD + 750 μg/kg ZEN | ||
| Week 2 | Egg weight (g) | 62.0 ± 1.5 | 60.6 ± 1.7 | 60.6 ± 1.4 |
| Egg index | 1.26 ± 0.04 | 1.27 ± 0.03 | 1.28 ± 0.02 | |
| Albumen height (mm) | 7.15 ± 1.62 | 7.40 ± 1.07 | 6.56 ± 0.60 | |
| Yolk color | 7.58 ± 1.73 | 8.75 ± 1.22 | 8.45 ± 2.14 | |
| Haugh unit | 83.92 ± 9.3 | 86.01 ± 6.3 | 80.67 ± 4.3 | |
| Egg shell thickness (mm) | 0.31 ± 0.02 | 0.32 ± 0.01 | 0.32 ± 0.02 | |
| Egg shell strength (N) | 44.7 ± 5.5 | 44.3 ± 7.3 | 47.6 ± 2.8 | |
| Week 5 | Egg weight (g) | 61.0 ± 2.0 | 60.4 ± 2.0 | 60.6 ± 2.9 |
| Egg index | 1.26 ± 0.03 | 1.27 ± 0.03 | 1.27 ± 0.01 | |
| Albumen height (mm) | 6.66 ± 0.66 | 6.30 ± 1.10 | 5.85 ± 0.73 | |
| Yolk color | 8.11 ± 0.92 | 8.37 ± 0.62 | 9.26 ± 1.75 | |
| Haugh unit | 80.75 ± 4.0 | 76.84 ± 8.1 | 74.85 ± 5.3 | |
| Egg shell thickness (mm) | 0.33 ± 0.01 | 0.33 ± 0.01 | 0.32 ± 0.01 | |
| Egg shell strength (N) | 42.9 ± 5.9 | 42.0 ± 6.6 | 44.5 ± 4.5 | |
| Items | Treatment Level | ||
|---|---|---|---|
| BD | BD + 250 μg/kg ZEN | BD + 750 μg/kg ZEN | |
| ZEN | <LOD | <LOD | <LOD |
| α-ZOL | <LOD | <LOD | <LOD |
| β-ZOL | <LOD | <LOD | <LOD |
| Items | Supplemental Levels of ZEN in Diets, μg/kg | ||
|---|---|---|---|
| BD | BD + 250μg/kg ZEN | BD + 750 μg/kg ZEN | |
| Body weight (kg) | 1.96 ± 0.12 | 1.92 ± 0.15 | 1.89 ± 0.11 |
| Ovarian index | 0.26 ± 0.07 | 0.22 ± 0.03 | 0.21 ± 0.09 |
| Oviductal index | 3.29 ± 0.25 | 2.96 ± 0.19 | 3.10 ± 0.42 |
| Items | Supplemental Levels of ZEN in Diets, μg/kg | ||
|---|---|---|---|
| BD | BD + 250 μg/kg ZEN | BD + 750 μg/kg ZEN | |
| Estradiol (pg/mL) | 449.28 ± 44.90 | 440.72 ± 19.80 | 451.08 ± 43.26 |
| Follicle-stimulating hormone (IU/L) | 7.62 ± 0.31 a | 6.63 ± 0.33 b | 6.46 ± 0.25 b |
| Luteinizing hormone (ng/mL) | 151.85 ± 12.41 a | 145.84 ± 12.44 a | 129.35 ± 6.72 b |
| Progesterone (pg/mL) | 485.78 ± 113.40 a | 359.67 ± 64.04 b | 286.56 ± 57.40 b |
| Ingredients | % | Nutrition Component | Content (%) |
|---|---|---|---|
| Corn | 51.90 | Moisture | 9.68 |
| Soybean meal | 16.00 | Crude protein | 16.05 |
| Distillers Dried Grains with soluble (crude protein: 27%) | 11.72 | ME (MJ/kg) | 11.28 |
| Extruded soybean | 4.00 | Crude Ash | 14.24 |
| Corn germ meal | 2.50 | Crude fat | 5.52 |
| Soybean oil | 2.43 | Crude fiber | 4.28 |
| Shell powder | 3.00 | Calcium | 3.76 |
| Limestone | 6.55 | Phosphorus | 0.52 |
| Monocalcium phosphate | 0.72 | Lysine | 0.85 |
| Phytase | 0.02 | Methionine | 0.48 |
| Premix a | 1.10 | Arginine | 0.91 |
| Total | 99.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, T.; Li, J.; Wang, Y.; Li, M.; Yang, A.; Ren, C.; Qi, D.; Zhang, N. Effects of Zearalenone on Production Performance, Egg Quality, Ovarian Function and Gut Microbiota of Laying Hens. Toxins 2022, 14, 653. https://doi.org/10.3390/toxins14100653
Yuan T, Li J, Wang Y, Li M, Yang A, Ren C, Qi D, Zhang N. Effects of Zearalenone on Production Performance, Egg Quality, Ovarian Function and Gut Microbiota of Laying Hens. Toxins. 2022; 14(10):653. https://doi.org/10.3390/toxins14100653
Chicago/Turabian StyleYuan, Tao, Junyi Li, Yanan Wang, Meiling Li, Ao Yang, Chenxi Ren, Desheng Qi, and Niya Zhang. 2022. "Effects of Zearalenone on Production Performance, Egg Quality, Ovarian Function and Gut Microbiota of Laying Hens" Toxins 14, no. 10: 653. https://doi.org/10.3390/toxins14100653
APA StyleYuan, T., Li, J., Wang, Y., Li, M., Yang, A., Ren, C., Qi, D., & Zhang, N. (2022). Effects of Zearalenone on Production Performance, Egg Quality, Ovarian Function and Gut Microbiota of Laying Hens. Toxins, 14(10), 653. https://doi.org/10.3390/toxins14100653