

Foraging Burrow Site Selection and Diet of Chinese Pangolins, Chandragiri Municipality, Nepal


1
Central Department of Zoology, Institute of Science and Technology, Tribhuvan University, Kirtipur, Kathmandu 44618, Nepal
2
Nepal Zoological Society, Kirtipur, Kathmandu 44618, Nepal
3
Department of Fisheries and Wildlife, Michigan State University, East Lansing, MI 48824, USA
*
Author to whom correspondence should be addressed.
Animals 2022, 12(19), 2518; https://doi.org/10.3390/ani12192518
Submission received: 21 August 2022
/
Revised: 18 September 2022
/
Accepted: 19 September 2022
/
Published: 21 September 2022
(This article belongs to the Section Ecology and Conservation)
Abstract
:Simple Summary
As one of only eight species of pangolin, the Chinese pangolin (Manis pentadactyla) of Nepal inhabits forests, agricultural lands, and grasslands. Its population is declining due to hunting and habitat loss, and it is listed as critically endangered. Accurate information on its habitat and diet can aid in the development of site-specific management plans. Habitat characteristics such as forest canopy cover, slopes, and distance to agricultural lands and the nearest ant nests are important factors influencing the occurrence of the Chinese pangolin. Fecal analysis revealed that the ant species Aphaenogaster symthiesii, Camponotus sp., Monomorium sp., and Pheidole sp. were the dominant prey in the Chinese pangolin’s diet. This study provides baseline information to aid Chinese pangolin conservation in Nepal.
Abstract
The Chinese pangolin (Manis pentadactyla) is a myrmecophagous, nocturnal mammal species that occurs in forests, agricultural lands, and grasslands. It is critically endangered due to illegal hunting and habitat loss. Characterizing the Chinese pangolin’s habitat and diet could improve our knowledge of the conditions necessary for species persistence; however, limited information is available. We investigated the habitat and diet of Chinese pangolins in the Chandragiri Municipality, Kathmandu, Nepal from November 2021–March 2022. We identified foraging burrows within plots established along 20 transects, collected scats opportunistically at these burrows, and used a generalized linear model to assess the site-level habitat characteristics related to burrow occurrence. We recorded 88 foraging burrows which occurred in forests with 50–75% canopy closure at 1500–1700 m elevation with 20–40° slopes. The probability of detecting a Chinese pangolin foraging burrow was greater with the increasing slope gradient and decreased with increasing distance to agricultural lands and ant nests or termite mounds. The analysis of 10 scats revealed that Aphaenogaster symthiesii, Camponotus sp., Monomorium sp., and Pheidole sp. were the dominant ant prey species; no termites were detected. Baseline data from this study could be used for ex-situ conservation and the captive breeding of Chinese pangolins as well as aiding site-specific management plans in Nepal.
1. Introduction
The foraging site selection and the diet of species provide insights into their ecology and habitat use [1]. The knowledge of foraging ecology and diet is essential for developing conservation action and management plans for species’ long-term persistence [2,3] and their impacts on the prey species [4]. This knowledge is particularly important for species that are endangered or are of conservation concern.
The Chinese pangolin (Manis pentadactyla, Linnaeus 1758) is one of the four Asian pangolin species and occurs in Nepal, India, China, Taiwan, Bangladesh, Bhutan, Myanmar, Lao PDR, Thailand, and Vietnam [5]. The Chinese pangolin occurs in primary and secondary tropical forests [6], grasslands, agricultural areas, and some degraded habitats [7,8,9,10,11]. Chinese pangolins tend to inhabit broad-leaved forests because of the greater abundance of termites [12]. However, their occurrence can also be influenced by elevation, slope gradient, canopy cover, and the distance to water and human activity [9,13,14,15]. Chinese pangolins are nocturnal and have adapted for digging burrows which are used for hunting prey, shelter, and avoiding predators [12,16].
The Chinese pangolin is considered vulnerable to human threats due to its low reproductive rate, poor self-defensive mechanisms, and narrow habitat requirements [12]. It is classified as Critically Endangered by the International Union for Conservation of Nature (IUCN) Red List of Threatened Species [5]. The Chinese pangolin is listed under Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora [17] and protected in Nepal under the National Parks and Wildlife Conservation (NPWC) Act 1973 [18]. Human threats to the Chinese pangolin include illegal hunting for its flesh and scales [5] as well as habitat alteration and degradation [7,11,19]. Pangolins are among the few myrmecophagous mammal species [20], with specialized anatomical and morphological adaptations for foraging primarily on termites and ants [21,22,23,24]. They opt for specific ant and termite species rather than foraging on the most abundant species [23,25,26]. The Chinese pangolin selects ants over termites as prey, feeding on over 70 species of ant and 4 termite species [27]. Dominant ant prey species include Pheidologeton yanoi, Pheidole nodus, Polyrachis fervens, Crematogaster schimmeri, Camponotus monju, and Pseudolasius binghami, and the dominant termite prey species is Odontotermes formosanus. Chinese pangolins also appear to help control the invasive ant Anoplolepis gracilipes [28]. Therefore, the conservation of the Chinese pangolin could help maintain ecosystems by regulating insect populations [5,19] and provide ecosystem services by improving soil quality and mitigating the crop damage caused by termites and ants [8,29].
Because of their nocturnality, adaptations for burrowing [30,31], and their tendency to forage 5–6 km from their resident burrows each night [32], the foraging behavior and ecology of pangolins are difficult to observe in the field. Their specialist diet also makes them difficult to maintain in captivity for observational studies [33]. Consequently, there is limited knowledge regarding the foraging ecology and diet of the Chinese pangolin. We characterized the foraging burrow site selection of the Chinese pangolin and identified prey remains from scats associated with these burrows from the Chandragiri Municipality, Nepal, to further our understanding of pangolin ecology and improve its conservation.
2. Materials and Methods
2.1. Study Area
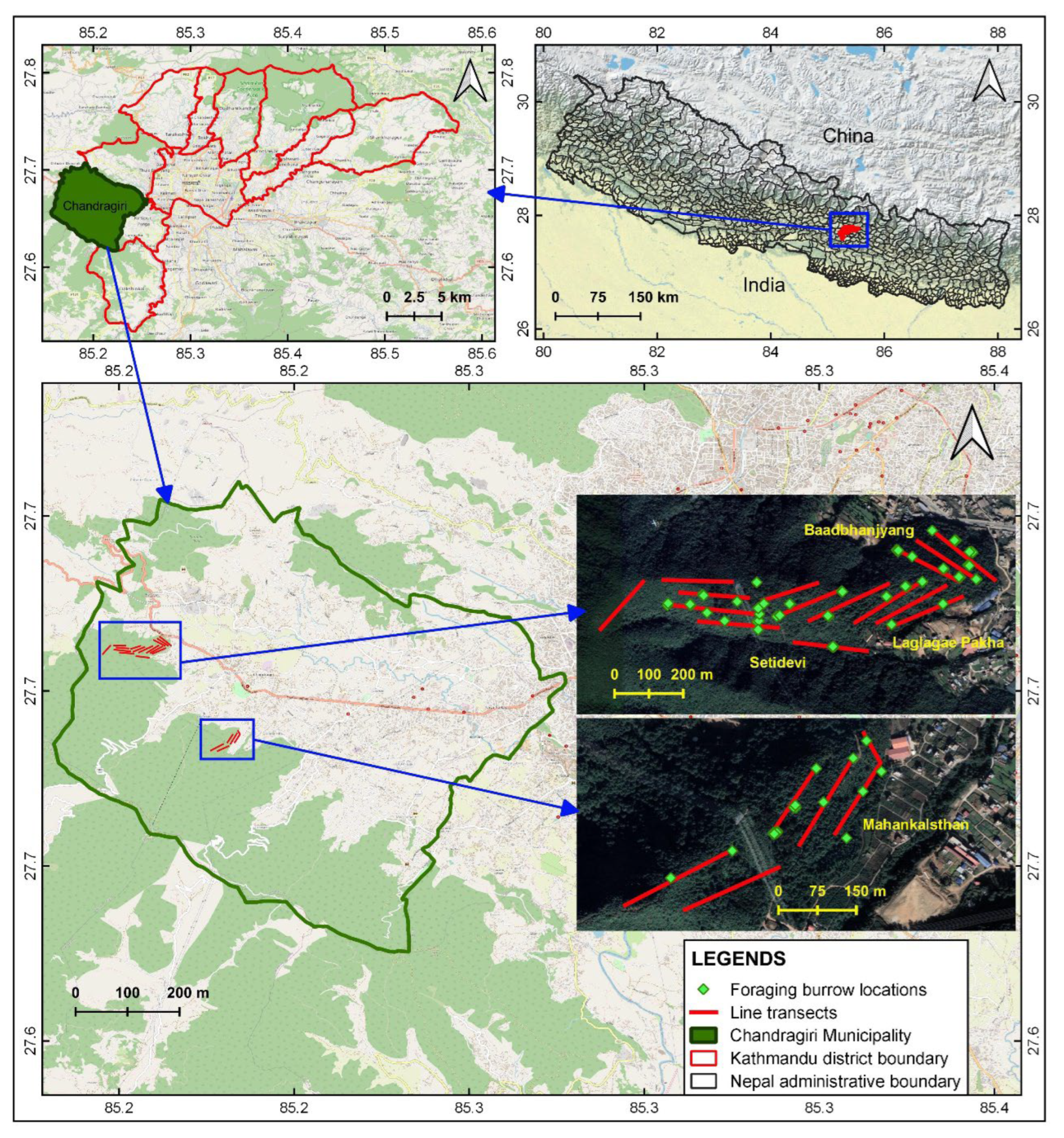
The Chandragiri Municipality (27°43′36.49″–27°32′45.03″ N, 85°16′39.51″–85°11′8.68″ E) is in southwest Kathmandu District, Bagmati Province, Nepal (Figure 1) and comprises 43.9 km2. It has a human population of 136,928 (3118 people/km2; [34]). It has predominantly hilly terrain with elevations from 1310 to 2551 m above sea level. It contains 23 community forests covering 1171 ha. The vegetation is mixed forest and includes the Nepalese alder (Alnus nepalensis), needlewood (Schima wallichii), chinkapin (Castanopsis tribuloides), pine (Pinus roxburghii), oak (Quercus spp.), rhododendron (Rhododendron arboretum), Himalayan ash (Fraxinus floribunda), and marking nut (Semicarpus anacardium). Major mammal species include the large Indian civet (Viverra zibetha), yellow-throated marten (Martes flavigula), jungle cat (Felis chaus), golden jackal (Canis aureus), Chinese pangolin (Manis pentadactyla), hoary-bellied squirrel (Callosciurus pygerythrus), leopard cat (Prionailurus bengalensis), leopard (Panthera pardus), and wild boar (Sus scrofa) [35].
2.2. Methods
We conducted our field survey from November 2021–March 2022 within the Mahankal, Setidevi, Laglagae, and Baadbhanjyang community forests in the Chandragiri Municipality. Within each forest we established five transects (20 total) about 300 m long and established five, 10 × 10-m plots on each transect at 50-m intervals (Figure 1). We established a total of 100 survey plots on the 20 transects. In each plot, we recorded the foraging burrow occurrence, collected the fecal samples observed, and measured eight habitat covariates. We counted the number of ant nests and termite mounds within each plot and measured the distance from the plot center to the nearest nest or mound. We estimated the slope gradient using a clinometer at the plot center. We estimated canopy coverage by averaging values obtained at the plot center and the four corners using a spherical densiometer. We counted the number of trees (>5-cm diameter at breast height and >1.5-m tall) in each plot. We then measured the distance from the plot center to the nearest road, water source (e.g., stream, pond), and agricultural land; distances <25 m were measured using a tape measure, with greater distances estimated using Google Earth Pro.
We collected Chinese pangolin fecal samples opportunistically from burrow openings to estimate diet composition, placing each in a plastic bag with desiccant before analyses. We identified fecal samples from the Chinese pangolin by visual observation and by detecting the presence of chitin fragments in the feces [36].
In the laboratory, we placed samples in 70% ethanol and separated items manually, and then used a stereomicroscope for identification. We identified prey to the lowest taxonomic level using keys for ants [37], termites [38], and other invertebrates, as well as a reference collection obtained during our surveys in the study area.
We used a generalized linear mixed model to identify factors affecting the occurrence of the Chinese pangolin in the Chandragiri Municipality in 2021. Factors include elevation (m), slope gradient (°), forest canopy cover (%), distance to a water source (m), roads (m), settlements (m), agricultural land (m), ant nests (m), no. of ant nests, and tree abundance. Elevation and distance to settlement were highly correlated variables (|r| > 0.7) with distance to agricultural land, and we retained the latter for analysis (Figure 2). We rescaled continuous variables from 0 to 1 before analyses. We used R program for analyses [39]. Means are reported with a +1 standard error (SE).
3. Results
We identified 88 foraging burrows in the 38 surveyed plots; 30 (34%) in the Mahankalsthan, 15 (17%) in the Setidevi, 24 (27%) in the Baadbhanjyang, and 19 (22%) in the Laglagae Pakha community forests. We located burrows 1450 to 1800 m above sea level [(average 1585 ± 11.6 m (SE)] with most burrows (81.8%, n = 81) within 1500–1700 m elevation (Table 1). Most burrows (46.5%, n = 46) were recorded on 30–40° slopes (Figure 3). Most burrows (61.4%, n = 54) were under 50–75% forest canopy coverage (Figure 4). The average distance to the nearest road from the center of the plot was 28.1 ± 0.9 m (range = 1 to 272.3 m), and most burrows (88.6%, n = 78) were <100 m from the nearest road. Most burrows (55.6%, n = 55) were <100 m from the nearest water source. The average distance to agricultural lands was 224 ± 28.2 m (range = 55–1025 m) with most burrows (78.8%, n = 78) recorded within 400 m of agricultural lands (Figure 5). The average distance to the nearest ant nest or termite mound from the center of plots was 5.4 ± 1.3 m (range = 0.6–38 m) with most burrows (72.7%, n = 72) within 20 m (Figure 6).
The probability of detecting a Chinese pangolin foraging burrow was greater with increasing slope and decreased with increasing distance to agricultural lands and ant nests or termite mounds (Table 2). No other variables measured were significant.
4. Discussion
The occurrence of Chinese pangolin foraging burrows was influenced by slope gradient, distance to agricultural lands, and distance to ant nests. The presence of burrows on slopes of 30–40° suggests moderately steep slopes are more suitable for foraging. Steeper slopes (30–60°) could maintain stable temperatures inside burrows and ensure the availability of termites [40], whereas the absence of burrows in slopes >60° could be due to reduced food availability and accessibility [12,40,41]. Moderately steeper slopes could also ensure the stability and integrity of burrows by reducing erosion and facilitate their excavation [40].
We found that canopy cover influenced pangolin burrow presence; that most burrows occurred in forests with 50–75% canopy coverage suggests that Chinese pangolins opt for intermediate to higher levels of canopy cover, as reported previously [9]. Chinese pangolins have poor defenses against predators [42], and a greater canopy cover could reduce predation risk [43]. Chinese pangolins have a poor capacity to adapt to changing temperatures, and denser canopy cover may also buffer against fluctuations in ambient temperature [40]. Additionally, ants and termites are more abundant and diverse in areas with denser forest canopies [44,45]. Finally, forests with denser canopies have less understory vegetation which could facilitate pangolin movements [46].
The occurrence of Chinese pangolin foraging burrows was greater nearer to agricultural areas, but the distance to water and roads did not influence burrow occurrence. Agricultural lands are abundant with ants and termites due to the presence of plant debris and animal dung [47]. However, anthropogenic activities such as the collection of fuel wood and fodder, livestock grazing, and the use of pesticides can cause disturbance and decreased prey availability for pangolins [15]. Moreover, pangolin activity near agricultural or other areas of human activity could increase their risk of being hunted [48]. The lack of relationship between the distance to water and burrow occurrence could be a consequence of the prevalence of water in the study area. Pangolins use water sources frequently and ants, termites, and other insects often select moist habitats [49,50]. We suggest that the abundance of water sources in our study area alleviated the need for the spatial selection of this resource.
The greater presence of Chinese pangolin foraging burrows near ant or termite mounds is undoubtedly a consequence of their myrmecophagous diet [12,43]. Burrows appear to be excavated near food sources [HPS personal observation] which would reduce energy expenditure when foraging [51]. Chinese pangolins feed almost exclusively on ants and termites with occasional feeding on other invertebrates. The presence of only four ant species (A. symthiesii, Camponotus sp., Monomorium sp., and Pheidole sp.) in the fecal samples despite that 15 ant species were collected from the study area as references suggest that the Chinese pangolin may exhibit selection among ant species. Several reasons could explain the species consumed: (1) pangolins may feed on the most available species; (2) pangolins may opt for larger (>5 mm body length) species. Larger ants could increase foraging efficiency and provide more energy and nutrients than smaller-sized prey [26]; and (3) pangolins may prioritize easy-to-capture prey [25,28]. Ant species recovered from scats in this study nest just below the soil surface or in decomposed logs which can be readily obtained by pangolins, particularly juveniles [28].
The selection of the ant species consumed might also be due to the chemical and mechanical defenses of the prey species; pangolins avoid ant and termite species with well-developed defense systems [52]. Ant species in the subfamily Ponerinae have strong defenses against predators [28], which could explain the avoidance of Ponerinae species such as Brachyponera and Ectomomyrmex by the pangolins. Previous studies suggest that pangolins prefer ants over termites [23,25,26,27] for reasons including difficulty accessing termites from mounds [41], which may partially explain why termites were not detected in Chinese pangolin fecal samples in this study. However, the complete digestion of termites that are relatively soft-bodied compared to ants could have limited our ability to detect them [53].
5. Conclusions
The foraging habitat selection of Chinese pangolins was influenced by slope and apparent food availability. Chinese pangolins may exhibit prey selection, but larger studies quantifying prey use and availability are required. Our study provides baseline data on the foraging habitat use and diet that could benefit ex-situ conservation efforts as well as captive breeding programs for Chinese pangolins. Furthermore, this information can be used to aid site-specific management plans in Nepal to protect or improve habitat suitability for Chinese pangolins.
Author Contributions
Conceptualization, S.T. and H.P.S.; Data collection, S.T.; Supervised, H.P.S.; Data Analyzed, S.T. and H.P.S. writing—original draft preparation, S.T. and H.P.S. writing—review and editing, S.T., H.P.S. and J.L.B. All authors have read and agreed to the published version of the manuscript.
Funding
The research was financially supported by the National Youth Council, Nepal Government, Ministry of Youth and Sports. The first author is grateful to the National Youth Council for this research grant.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank the Department of Forests and Soil Conservation (Permit ID: 236) and Chandragiri Municipality (Permit ID: 787) for permission to conduct this research. We thank the Central Department of Zoology, Tribhuvan University for laboratory facilities. We thank Indra Prasad Subedi for ant species identification. We thank Krishna Rana, Kuber Bahadur Singh, Lila Gurung, Nita Kumari Somai and Parvati Gharti for field assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zweifel-Schielly, B.; Leuenberger, Y.; Kreuzer, M.; Suter, W. A herbivore’s food landscape: Seasonal dynamics and nutritional implications of diet selection by a red deer population in contrasting A lpine habitats. J. Zool. 2012, 286, 68–80. [Google Scholar] [CrossRef]
- Challender, D. Asian Pangolins: How behavioural research can contribute to their conservation. In Proceedings of the Workshop on trade and conservation of pangolins native to South and Southeast Asia, Singapore, 30 June–2 July 2008; p. 95. [Google Scholar]
- Redford, K.H. Dietary specialization and variation in two mammalian myrmecophages (variation in mammalian myrmecophagy). Rev. Chil. Hist. Nat. 1986, 59, 201–208. [Google Scholar]
- Klare, U.; Kamler, J.F.; Macdonald, D.W. A comparison and critique of different scat-analysis methods for determining carnivore diet. Mammal Rev. 2011, 41, 294–312. [Google Scholar] [CrossRef]
- Challender, D.; Wu, S.; Kaspal, P.; Khatiwada, A.; Ghose, A.; Sun, N.C.-M.; Mohapatra, R.; Suwal, T.L. Manis pentadactyla (Errata Version Published in 2020). The IUCN Red List of Threatened Species 2019: E. T12764A168392151. Available online: https://dx.doi.org/10.2305/IUCN.UK.2019-3.RLTS.T12764A168392151.en (accessed on 1 March 2022).
- Chakraborty, S.; Ramakrishna, S.; Chakraborty, R. Pangolins (Mammlia: Pholiodota) of India. Zool. Soc. India ENVIS Newsl. 2002, 9, 5–9. [Google Scholar]
- Gurung, J.B. A pangolin survey in Royal Nagarjung Forest in Kathmandu, Nepal. Tigerpaper 1996, 23, 29–32. [Google Scholar]
- Sharma, H.P.; Rimal, B.; Zhang, M.; Sharma, S.; Poudyal, L.P.; Maharjan, S.; Kunwar, R.; Kaspal, P.; Bhandari, N.; Baral, L. Potential distribution of the critically endangered Chinese Pangolin (Manis pentadactyla) in different land covers of Nepal: Implications for conservation. Sustainability 2020, 12, 1282. [Google Scholar] [CrossRef]
- Suwal, T.L.; Thapa, A.; Gurung, S.; Aryal, P.C.; Basnet, H.; Basnet, K.; Shah, K.B.; Thapa, S.; Koirala, S.; Dahal, S. Predicting the potential distribution and habitat variables associated with pangolins in Nepal. Glob. Ecol. Conserv. 2020, 23, e01049. [Google Scholar] [CrossRef]
- DNPWC; DoF. Pangolin Conservation Action Plan for Nepal (2018–2022); Department of National Parks and Wildlife Conservation and Department of Forests: Kathmandu, Nepal, 2018; pp. 1–29. [Google Scholar]
- Chao, J.; Tsao, E.; Traylor-Holzer, K.; Reed, D.; Leus, K. Formosan Pangolin Population and Habitat Viability Assessment: Final Report; IUCN/SSC Conservation Breeding Specialist Group: Apple Valley, MN, USA, 2005. [Google Scholar]
- Wu, S.; Ma, G.; Chen, H.; Xu, Z.; Li, Y.; Liu, N. A preliminary study on burrow ecology of Manis pentadactyla. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2004, 15, 401–407. [Google Scholar]
- Dorji, D.; Chong, J.L.; Dorji, T. Habitat preference and current distribution of Chinese Pangolin (Manis pentadactyla L. 1758) in Dorokha Dungkhag, Samtse, southern Bhutan. J. Threat. Taxa 2020, 12, 16424–16433. [Google Scholar] [CrossRef]
- Shrestha, A.; Bhattarai, S.; Shrestha, B.; Koju, N.P. Factors influencing the habitat choice of pangolins (Manis spp.) in low land of Nepal. Ecol. Evol. 2021, 11, 14689–14696. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, H.P.; Chaulagain, C.; Katuwal, H.B.; Belant, J.L. Estimating occupancy of Chinese pangolin (Manis pentadactyla) in a protected and non-protected area of Nepal. Ecol. Evol. 2020, 10, 4303–4313. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Wu, S.; Su, C.; Yang, L.; Zhang, F.; Ma, G. Air temperature changes in a burrow of Chinese pangolin, Manis pentadactyla, in winter. J. Vertebr. Biol. 2013, 62, 42–47. [Google Scholar] [CrossRef]
- CITES. Convention on International Trade in Endangered Species of Wild Fauna and Flora. Appendices I, II and III (22/06/2021). 2022, p. 78. Available online: https://cites.org/sites/default/files/eng/app/2021/E-Appendices-2021-06-22.pdf (accessed on 3 March 2022).
- Jnawali, S.; Baral, H.; Lee, S.; Acharya, K.; Upadhyay, G.; Pandey, M.; Griffiths, J. The status of Nepal mammals: The national red list series; Department of National Parks and Wildlife Conservation: Kathmandu, Nepal, 2011; pp. 1–266. [Google Scholar]
- Sharma, S.; Sharma, H.P.; Katuwal, H.B.; Belant, J.L. Knowledge of the Critically Endangered Chinese pangolin (Manis pentadactyla) by local people in Sindhupalchok, Nepal. Glob. Ecol. Conserv. 2020, 23, e01052. [Google Scholar] [CrossRef]
- Redford, K.H. Mammalian myrmecophagy: Feeding, foraging and food preference. Ph.D. Dissertation, Harvard University, Boston, MA, USA, 1983. [Google Scholar]
- Delsuc, F.; Metcalf, J.L.; Parfrey, L.W.; Song, S.J.; González, A.; Knight, R. Convergence of gut microbiomes in myrmecophagous mammals. Mol. Ecol. 2014, 23, 1301–1317. [Google Scholar] [CrossRef]
- Jacobsen, N.G.; Newbery, R.; De Wet, M.; Viljoen, P.; Pietersen, E. A contribution of the ecology of the Steppe pangolin Manis temminckii in the Transvaal. Z. Säugetierkunde 1991, 56, 94–100. [Google Scholar]
- Pietersen, D.W.; Symes, C.T.; Woodborne, S.; McKechnie, A.E.; Jansen, R. Diet and prey selectivity of the specialist myrmecophage, Temminck’s ground pangolin. J. Zool. 2016, 298, 198–208. [Google Scholar] [CrossRef]
- Sun, N.C.M.; Lo, F.H.Y.; Chen, B.Y.; Yu, H.Y.; Liang, C.C.; Lin, C.C.; Chin, S.C.; Li, H.F. Digesta retention time and recovery rates of ants and termites in Chinese pangolins (Manis pentadactyla). Zoo Biol. 2020, 39, 168–175. [Google Scholar] [CrossRef]
- Panaino, W.; Parrini, F.; Phakoago, M.V.; Smith, D.; van Dyk, G.; Fuller, A. Do seasonal dietary shifts by Temminck’s pangolins compensate for winter resource scarcity in a semi-arid environment? J. Arid Environ. 2022, 197, 104676. [Google Scholar] [CrossRef]
- Swart, J.; Richardson, P.; Ferguson, J. Ecological factors affecting the feeding behaviour of pangolins (Manis temminckii). J. Zool. 1999, 247, 281–292. [Google Scholar] [CrossRef]
- Sun, N.C.-M.; Liang, C.-C.; Lin, C.-C.; Pei, K.J.-C.; Li, H.-F. Seasonal feeding ecology of an obligate myrmecophagous mammal, Chinese pangolin. In Proceedings of the Pacific Rim Termite Research Group Conference, Pacific Rim Termite Research Group, Taipei, Taiwan, 12–13 February 2020; pp. 112–118. [Google Scholar]
- Lee, R.H.; Cheung, K.; Fellowes, J.R.; Guénard, B. Insights into the Chinese pangolin’s (Manis pentadactyla) diet in a Peri-Urban habitat: A case study from Hong Kong. Trop. Conserv. Sci. 2017, 10, 1940082917709648. [Google Scholar] [CrossRef]
- Chao, J.-T.; Li, H.-F.; Lin, C.-C. The role of pangolins in ecosystems. In Pangolins; Elsevier: Amsterdam, The Netherlands, 2020; pp. 43–48. [Google Scholar]
- Bruce, T.; Kamta, R.; Mbobda, R.B.T.; Kanto, S.T.; Djibrilla, D.; Moses, I.; Deblauwe, V.; Njabo, K.; LeBreton, M.; Ndjassi, C. Locating giant ground pangolins (Smutsia gigantea) using camera traps on burrows in the Dja Biosphere Reserve, Cameroon. Trop. Conserv. Sci. 2018, 11, 1940082917749224. [Google Scholar] [CrossRef]
- Heath, M.; Coulson, I. Home range size and distribution in a wild population of Cape pangolins, Manis temminckii, in north-west Zimbabwe. Afr. J. Ecol. 1997, 35, 94–109. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, L. Pangolin’s habits and its resource protection. Chin. J. Zool. 1981, 16, 40–41. [Google Scholar]
- Yang, C.W.; Chen, S.; Chang, C.Y.; Lin, M.F.; Block, E.; Lorentsen, R.; Chin, J.S.; Dierenfeld, E.S. History and dietary husbandry of pangolins in captivity. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2007, 26, 223–230. [Google Scholar] [CrossRef]
- CBS. Central Bureau of Statistics. Preliminary Report of National Population 2021. 2021. Available online: https://censusnepal.cbs.gov.np/Home/Details?tpid=1&dcid=0f011f13-7ef6-42dd-9f03-c7d309d4fca3 (accessed on 15 June 2022).
- Katuwal, H.B.; Basent, H.; Sharma, H.P.; Koirala, S.; Khanal, B.; Neupane, K.R.; Thapa, K.B.; Panta, D.B.; Parajuli, K.; Lamichhane, S. Wildlife assessment of the Chandragiri hills, Kathmandu: Potentiality for ecotourism. Eur. J. Ecol. 2020, 6, 27–50. [Google Scholar] [CrossRef]
- Chame, M. Terrestrial mammal feces: A morphometric summary and description. Memórias Inst. Oswaldo Cruz 2003, 98, 71–94. [Google Scholar] [CrossRef]
- Nazarreta, R.; Buchori, D.; Hidayat, P.; Fardiansah, R.; Scheu, S.; Drescher, J. A Guide to the Ants of Jambi (Sumatra, Indonesia)–Identification Key to Common Ant Genera and Images of the EFForTS Collection. Version 4.0 Beta. Animal Ecology, Johann-Friedrich-Blumenbach Institute for Zoology and Anthropology; University of Göttingen: Göttingen, Germany, 2019. [Google Scholar]
- Kalleshwaraswamy, C.; Nagaraju, D.; Viraktamath, C. Illustrated identification key to common termite (Isoptera) genera of south India. Biosystematica 2013, 7, 11–21. [Google Scholar]
- Team, R.C. R: A Language and Environmentfor Statistical Computing; R Foundation forStatistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 15 June 2022).
- Wu, S.; Liu, N.; Ma, G.; Xu, Z.; Chen, H. Habitat selection by Chinese pangolin (Manis pentadactyla) in winter in Dawuling Natural Reserve. Mammalia 2003, 67, 493–502. [Google Scholar] [CrossRef]
- Heath, M.E. Manis pentadactyla. Mamm. Species 1992, 414, 1–6. [Google Scholar] [CrossRef]
- Nowak, R.M.; Walker, E.P. Walker’s Mammals of the World, 6th ed.; Johns Hopkins University Press: Baltimore, MD, USA, 1999; Volume 1, pp. 1–1936. [Google Scholar]
- Maurice, M.E.; lionel Ebong, E.; Fuashi, N.A.; Godwill, I.I.; Zeh, A.F. The ecological impact on the distribution of pangolins in Deng-Deng National Park, Eastern Region, Cameroon. Glob. J. Ecol. 2019, 4, 8–14. [Google Scholar]
- Axelsson, E.; Andersson, J. A case study of termite mound occurrence in relation to forest edges and canopy cover within the Barandabhar forest corridor in Nepal. Int. J. Biodivers. Conserv. 2012, 4, 633–641. [Google Scholar]
- Corro, E.J.; Ahuatzin, D.A.; Jaimes, A.A.; Favila, M.E.; Ribeiro, M.C.; López-Acosta, J.C.; Dáttilo, W. Forest cover and landscape heterogeneity shape ant–plant co-occurrence networks in human-dominated tropical rainforests. Landsc. Ecol. 2019, 34, 93–104. [Google Scholar] [CrossRef]
- Karawita, H.; Perera, P.; Gunawardane, P.; Dayawansa, N. Habitat preference and den characterization of Indian Pangolin (Manis crassicaudata) in a tropical lowland forested landscape of southwest Sri Lanka. PLoS ONE 2018, 13, e0206082. [Google Scholar] [CrossRef]
- Richer, R.; Coulson, I.; Heath, M. Foraging behaviour and ecology of the Cape pangolin (Manis temminckii) in north-western Zimbabwe. Afr. J. Ecol. 1997, 35, 361–369. [Google Scholar] [CrossRef]
- Katuwal, H.B.; Sharma, H.P.; Parajuli, K. Anthropogenic impacts on the occurrence of the critically endangered Chinese pangolin (Manis pentadactyla) in Nepal. J. Mammal 2017, 98, 1667–1673. [Google Scholar] [CrossRef]
- Shrestha, S.; Bashyal, A.; Rijal, R.; Shrestha, J.; Shrestha, P.; Shrestha, N.; McGreevy, T.J., Jr.; Buffum, B.; Khanal, S.N. An Ecological Assessment of Critically Endangered Chinese Pangolin Manis pentadactyla (Mammalia: Pholidota: Manidae) in the Midhills Region of Nepal. Open J. Ecol. 2021, 11, 344–356. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Osbrink, W.L. Effect of soil type and moisture availability on the foraging behavior of the Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2010, 103, 799–807. [Google Scholar] [CrossRef]
- Heath, M.E.; Vanderlip, S.L. Biology, husbandry, and veterinary care of captive Chinese pangolins (Manis pentadactyla). Zoo Biol. 1988, 7, 293–312. [Google Scholar] [CrossRef]
- Redford, K.H. Feeding and food preference in captive and wild giant anteaters (Myrmecophaga tridactyla). J. Zool. 1985, 205, 559–572. [Google Scholar] [CrossRef]
- Katdare, B.; Bharti, H.; Narvekar, N.; Singh, A.C. Ant species consumed in diet as prey species by Indian pangolin (Manis crassicaudata) in northern Western Ghats of Maharashtra India. Int. J. Zool. Stud. 2021, 6, 36–40. [Google Scholar]
Figure 1.
Chinese pangolin study area with established line transects and foraging burrows in Chandragiri Municipality, Nepal.
Figure 1.
Chinese pangolin study area with established line transects and foraging burrows in Chandragiri Municipality, Nepal.

Figure 2.
Correlation matrix between predictive variables to estimate factors influencing Chinese pangolin foraging burrow occurrence in Chandragiri Municipality, Nepal. Variables Ant_nests = distance to nearest ant nest, Road = distance to nearest road, canopy_coverage = % forest canopy coverage, Tree_abundance = number of trees, Settlement = distance to nearest settlement, Agricultural_land = Distance to nearest agricultural land, Ant_nest_no = number of ant nests and water_source = distance to nearest water source.
Figure 2.
Correlation matrix between predictive variables to estimate factors influencing Chinese pangolin foraging burrow occurrence in Chandragiri Municipality, Nepal. Variables Ant_nests = distance to nearest ant nest, Road = distance to nearest road, canopy_coverage = % forest canopy coverage, Tree_abundance = number of trees, Settlement = distance to nearest settlement, Agricultural_land = Distance to nearest agricultural land, Ant_nest_no = number of ant nests and water_source = distance to nearest water source.

Figure 3.
Chinese pangolin foraging burrow distribution by slope gradient in Chandragiri Municipality, Nepal.
Figure 3.
Chinese pangolin foraging burrow distribution by slope gradient in Chandragiri Municipality, Nepal.

Figure 4.
Chinese pangolin foraging burrow distribution by forest canopy coverage in Chandragiri Municipality, Nepal.
Figure 4.
Chinese pangolin foraging burrow distribution by forest canopy coverage in Chandragiri Municipality, Nepal.

Figure 5.
Chinese pangolin foraging burrow distribution by distance to agricultural lands in Chandragiri Municipality, Nepal.
Figure 5.
Chinese pangolin foraging burrow distribution by distance to agricultural lands in Chandragiri Municipality, Nepal.

Figure 6.
Chinese pangolin foraging burrow distribution by distance to ant nests in Chandragiri Municipality, Nepal.
Figure 6.
Chinese pangolin foraging burrow distribution by distance to ant nests in Chandragiri Municipality, Nepal.

Figure 7.
Ant body parts found in the fecal samples of the Chinese pangolin in Chandragiri Municipality, Nepal.
Figure 7.
Ant body parts found in the fecal samples of the Chinese pangolin in Chandragiri Municipality, Nepal.

Figure 8.
Insect body remains from fecal samples of Chinese pangolins in Chandragiri Municipality, Nepal.
Figure 8.
Insect body remains from fecal samples of Chinese pangolins in Chandragiri Municipality, Nepal.

Figure 9.
Major ant species present in the diet of Chinese pangolins in Chandragiri Municipality, Nepal.
Figure 9.
Major ant species present in the diet of Chinese pangolins in Chandragiri Municipality, Nepal.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Habitat variables for sites with (n = 38) and without (n = 62) Chinese pangolin foraging burrows, Chandragiri Municipality, Nepal.
Table 1.
Habitat variables for sites with (n = 38) and without (n = 62) Chinese pangolin foraging burrows, Chandragiri Municipality, Nepal.
| Variable | With Burrow | Without Burrow | ||||
|---|---|---|---|---|---|---|
| Mean | SE | Range | Mean | SE | Range | |
| Elevation (m) | 1585 | 11.6 | 1461–1737 | 1610 | 15.3 | 1454–1886 |
| Slope (°) | 28.1 | 0.9 | 16–40 | 24 | 1.1 | 8–40 |
| Forest canopy coverage (%) | 66.8 | 2.5 | 1.5–80.8 | 65.8 | 1.6 | 12.2–82 |
| Distance to road (m) | 40.8 | 8.8 | 2–272.3 | 18.5 | 6.4 | 1–239.7 |
| Distance to water (m) | 93.3 | 14.1 | 5.5–376.9 | 73.1 | 16.9 | 1.8–536.5 |
| Distance to settlement (m) | 284.3 | 26.2 | 50–939.8 | 233.7 | 35.4 | 47.3–656.6 |
| Distance to agricultural lands (m) | 245 | 28.2 | 57.8–694.6 | 329.2 | 37.3 | 63.8–1024.8 |
| Number of ant nests | 1.6 | 0.2 | 1–8 | 1 | 0.1 | 1–4 |
| Distance to ant nest (m) | 5.4 | 1.3 | 0.6–38 | 10 | 1.6 | 0.5–48 |
| Tree abundance | 13.1 | 1.03 | 1–29 | 12.5 | 0.7 | 3–27 |
Table 2.
Generalized linear mixed model estimates and 95% confidence limits describing the Chinese pangolin occurrence in Chandragiri Municipality, Nepal. Variables include slope gradient (°), forest canopy coverage (%), distance to water source (m), road (m), agricultural land (m), and ant nests (m), no. of ant nests, and tree abundance were included in model construction. Significant effects (p < 0.05) are in bold.
Table 2.
Generalized linear mixed model estimates and 95% confidence limits describing the Chinese pangolin occurrence in Chandragiri Municipality, Nepal. Variables include slope gradient (°), forest canopy coverage (%), distance to water source (m), road (m), agricultural land (m), and ant nests (m), no. of ant nests, and tree abundance were included in model construction. Significant effects (p < 0.05) are in bold.
| Variables | Estimate | Standard Error | Z-Score | p |
|---|---|---|---|---|
| (Intercept) | 0.928 | 1.297 | 0.716 | 0.474 |
| Slope gradient (°) | 2.612 | 1.117 | 2.338 | 0.019 |
| Forest canopy coverage (%) | −3.260 | 1.640 | −1.988 | 0.047 |
| Distance to road (m) | 1.483 | 1.198 | 1.238 | 0.216 |
| Distance to water source (m) | −0.291 | 1.384 | −0.210 | 0.834 |
| Distance to agricultural land (m) | −3.224 | 1.180 | −2.731 | 0.006 |
| Distance to ant nest (m) | −2.720 | 1.203 | −2.261 | 0.024 |
| Number of ant nests | 1.099 | 1.869 | 0.588 | 0.557 |
| Tree abundance | 2.385 | 1.504 | 1.586 | 0.113 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tamang, S.; Sharma, H.P.; Belant, J.L. Foraging Burrow Site Selection and Diet of Chinese Pangolins, Chandragiri Municipality, Nepal. Animals 2022, 12, 2518. https://doi.org/10.3390/ani12192518
AMA Style
Tamang S, Sharma HP, Belant JL. Foraging Burrow Site Selection and Diet of Chinese Pangolins, Chandragiri Municipality, Nepal. Animals. 2022; 12(19):2518. https://doi.org/10.3390/ani12192518
Chicago/Turabian StyleTamang, Sharmila, Hari Prasad Sharma, and Jerrold L. Belant. 2022. "Foraging Burrow Site Selection and Diet of Chinese Pangolins, Chandragiri Municipality, Nepal" Animals 12, no. 19: 2518. https://doi.org/10.3390/ani12192518
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.