
Phenotypic Differences and Physiological Responses of Salt Resistance of Walnut with Four Rootstock Types

 ,
,
Abstract
:1. Research Background
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Determination of Growth Indicators
2.3. Determination of Physiological and Biochemical Indicators
2.3.1. Determination of the Relative Electrical Conductivity
2.3.2. Determination of the Chlorophyll Content
2.3.3. Determination of Photosynthetic Parameters
2.3.4. Determination of the Malondialdehyde Content
2.3.5. Determination of the Osmotic Adjustment Substances
Determination of the Proline Content
Determination of the Soluble Sugar Content
2.3.6. Determination of the Activity of Antioxidant Enzymes
2.4. Statistical Analysis
3. Results
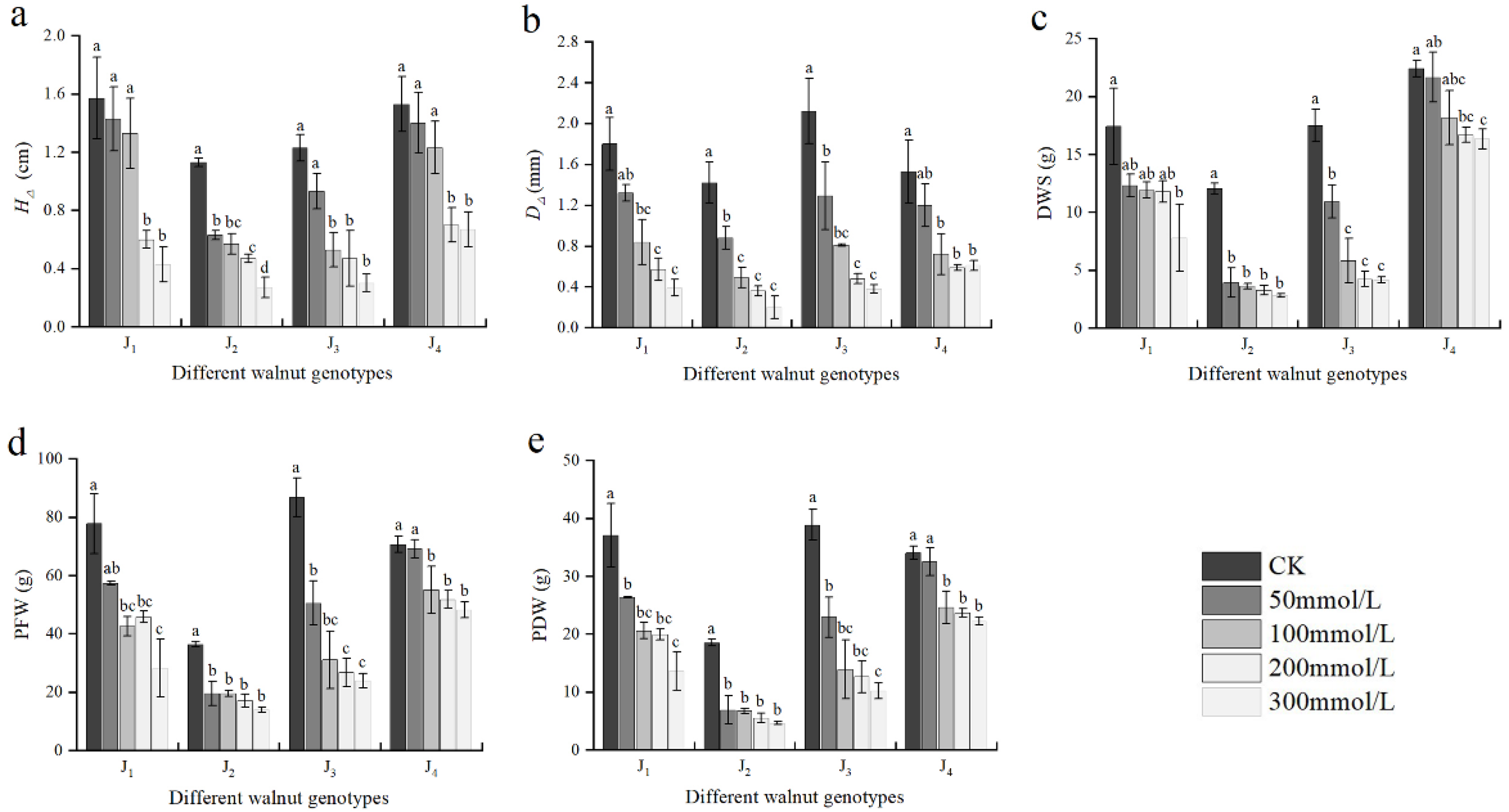
3.1. Effects of Salt Stress on External Morphological Characteristics of Seedlings
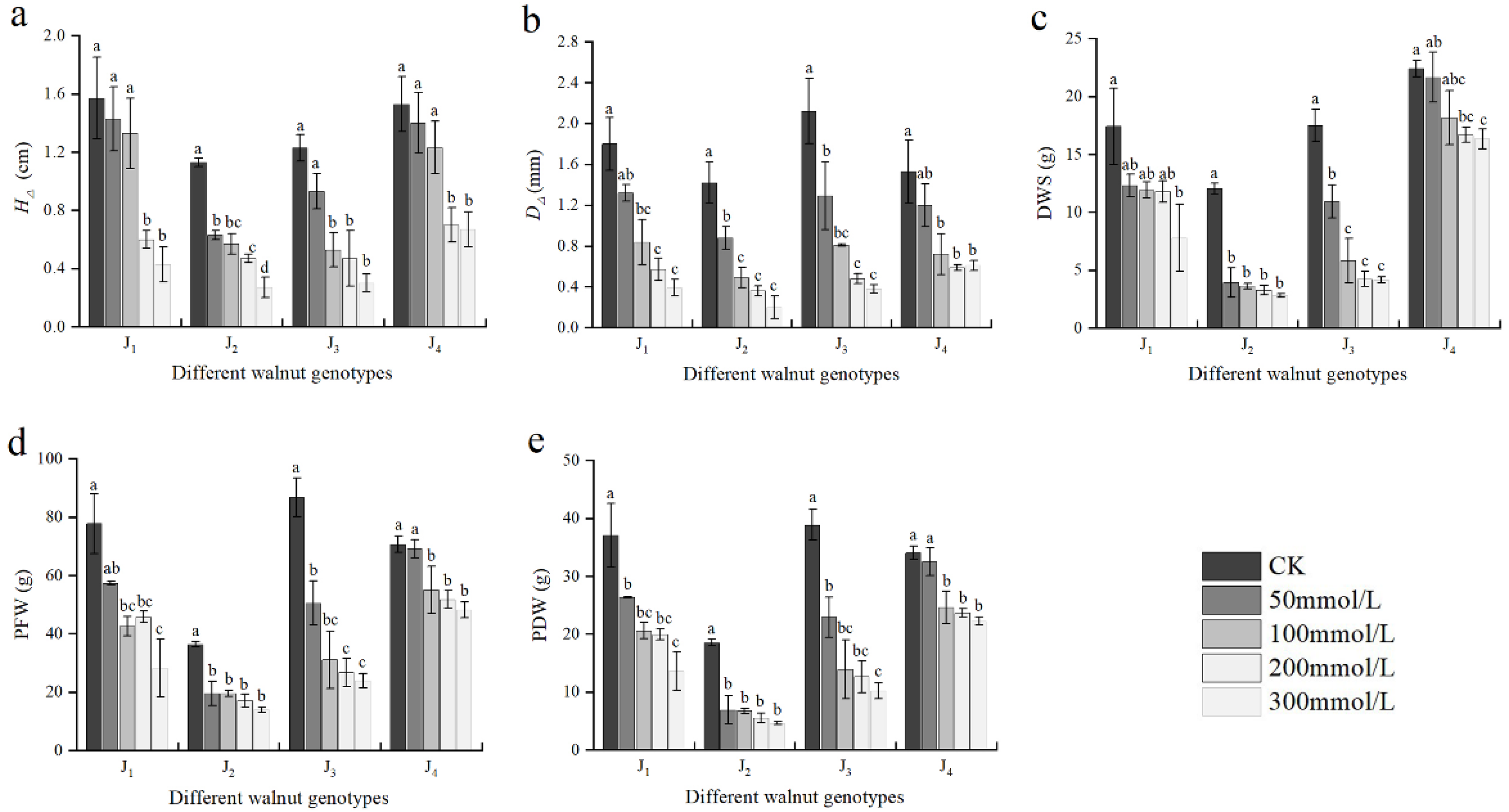
3.2. Effect of Salt Stress on Plant Growth
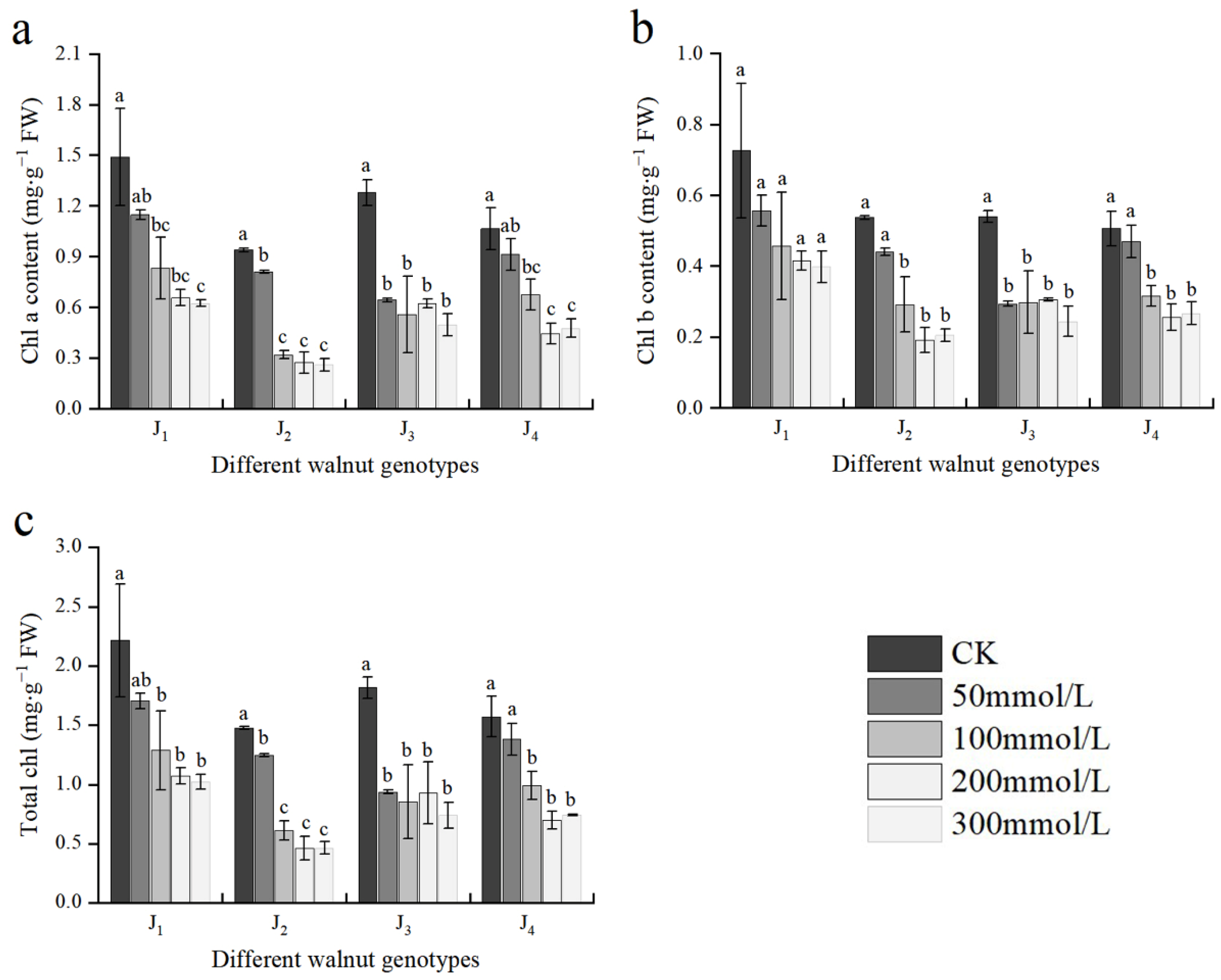
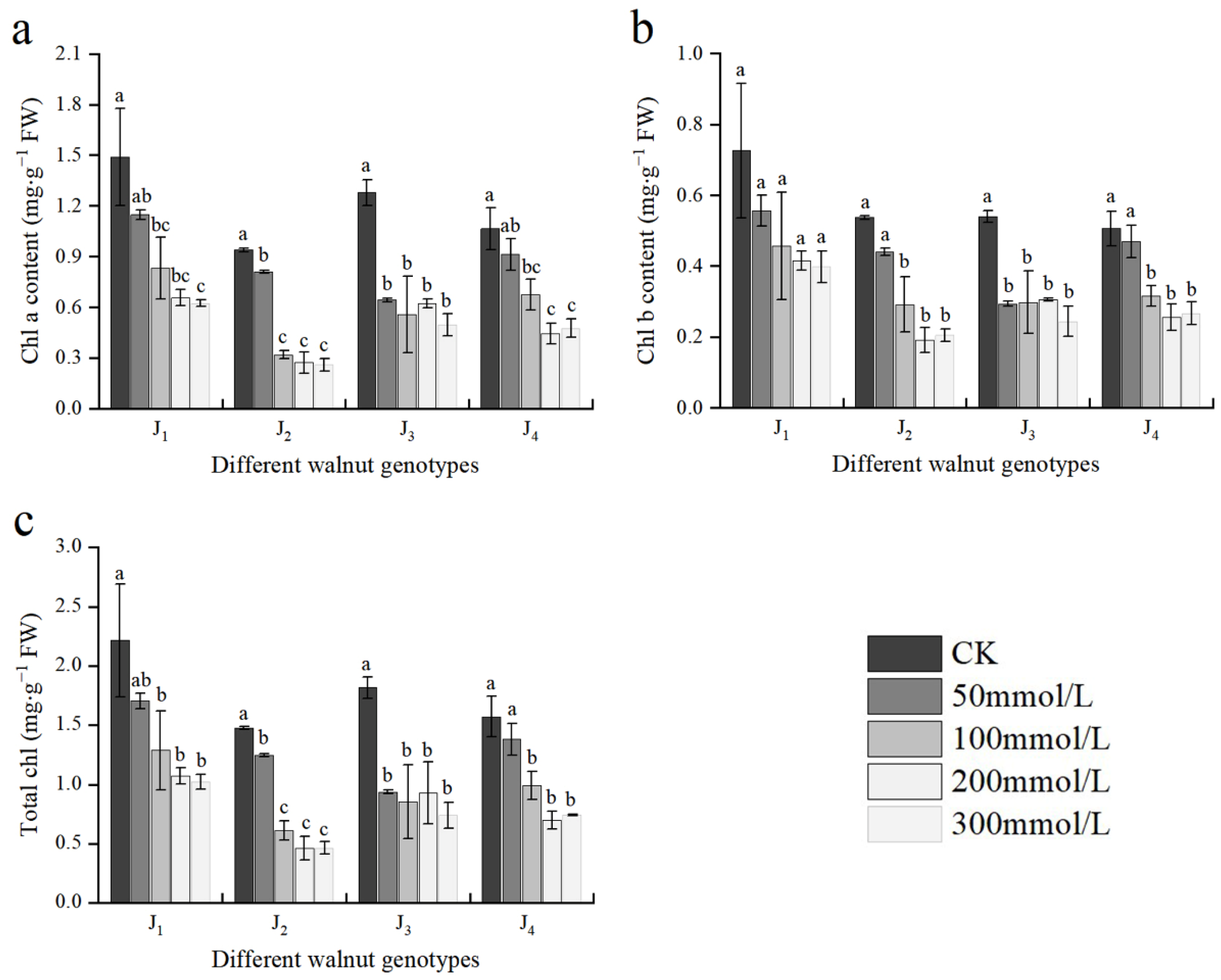
3.3. Effect of Salt Stress on Chlorophyll Content
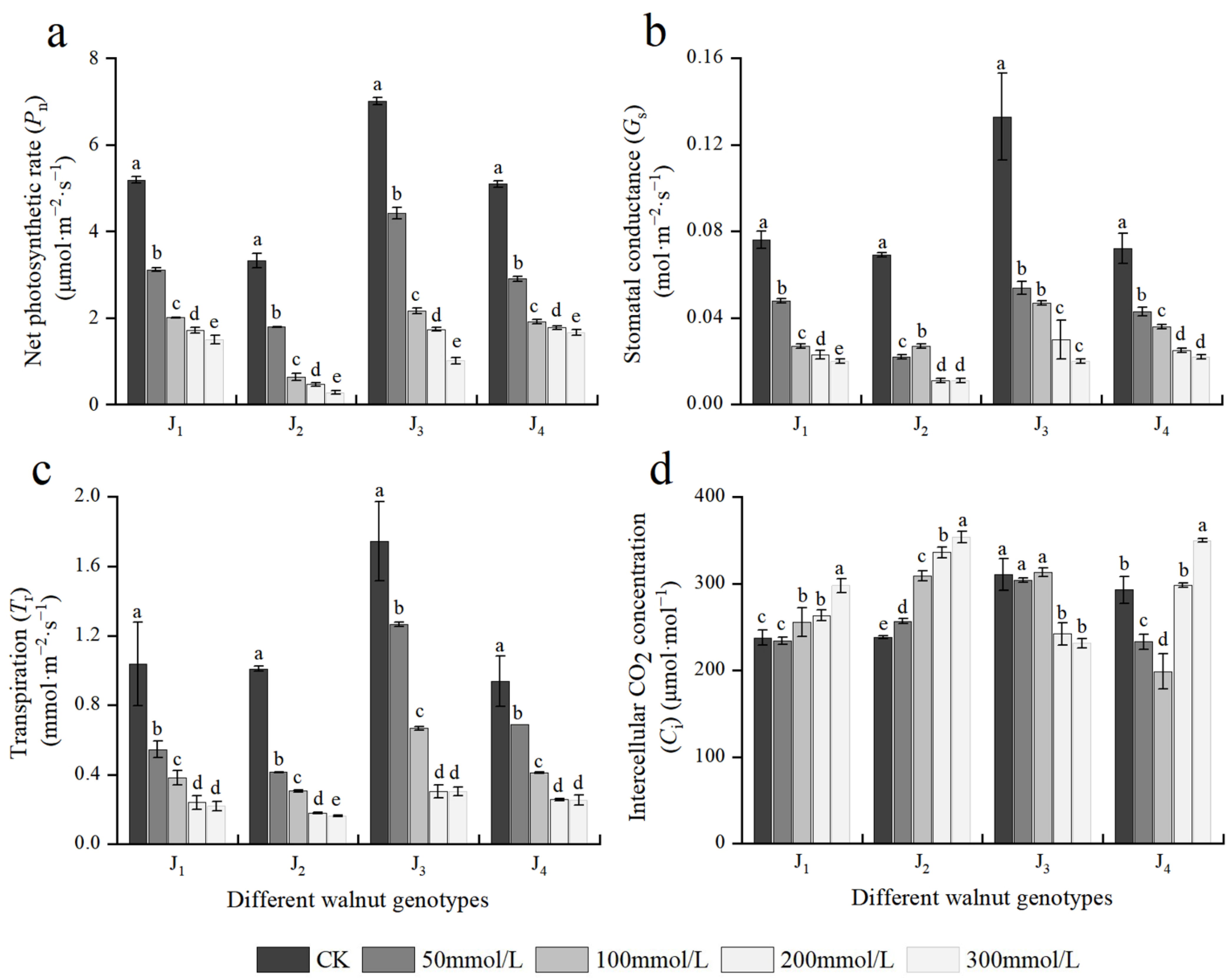
3.4. Effects of Salt Stress on Photosynthetic Gas Exchange Parameters
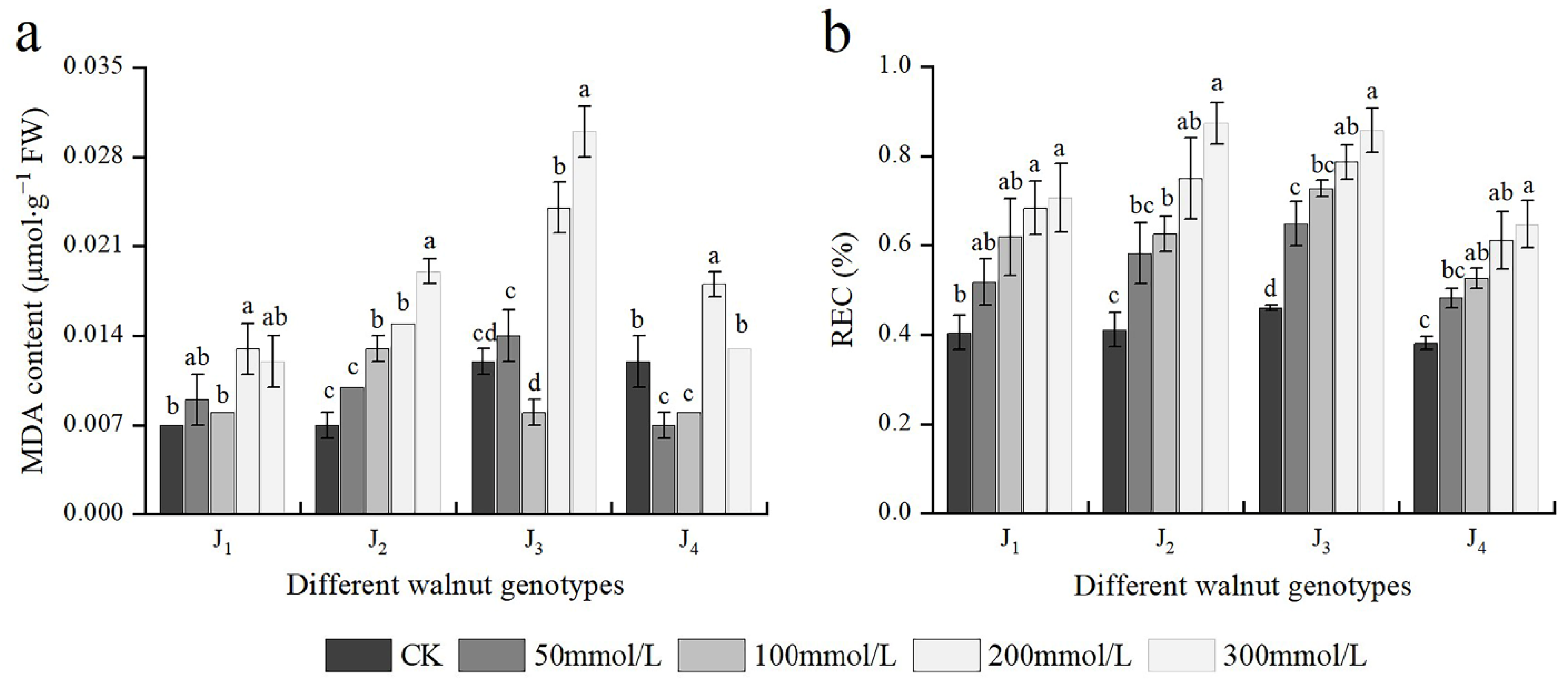
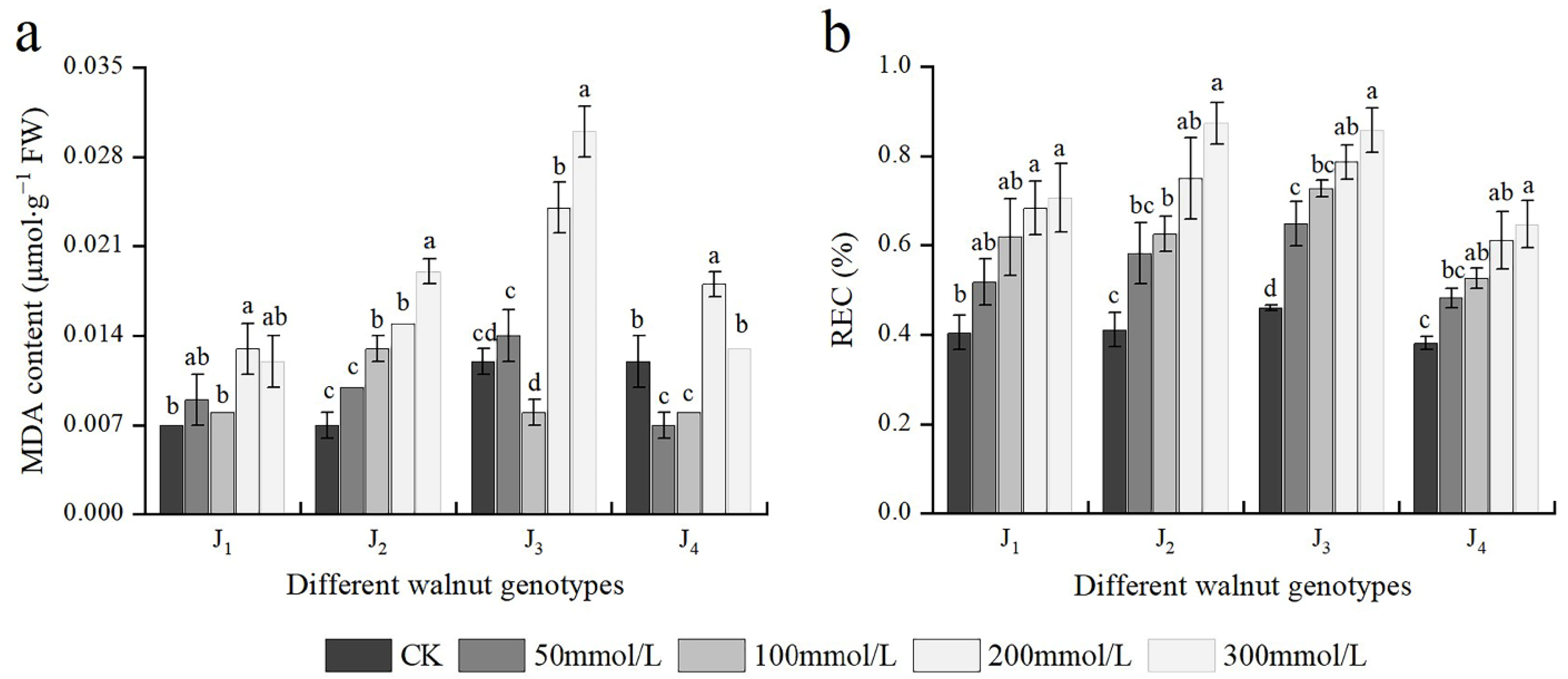
3.5. Effects of Salt Stress on MDA Content and REC
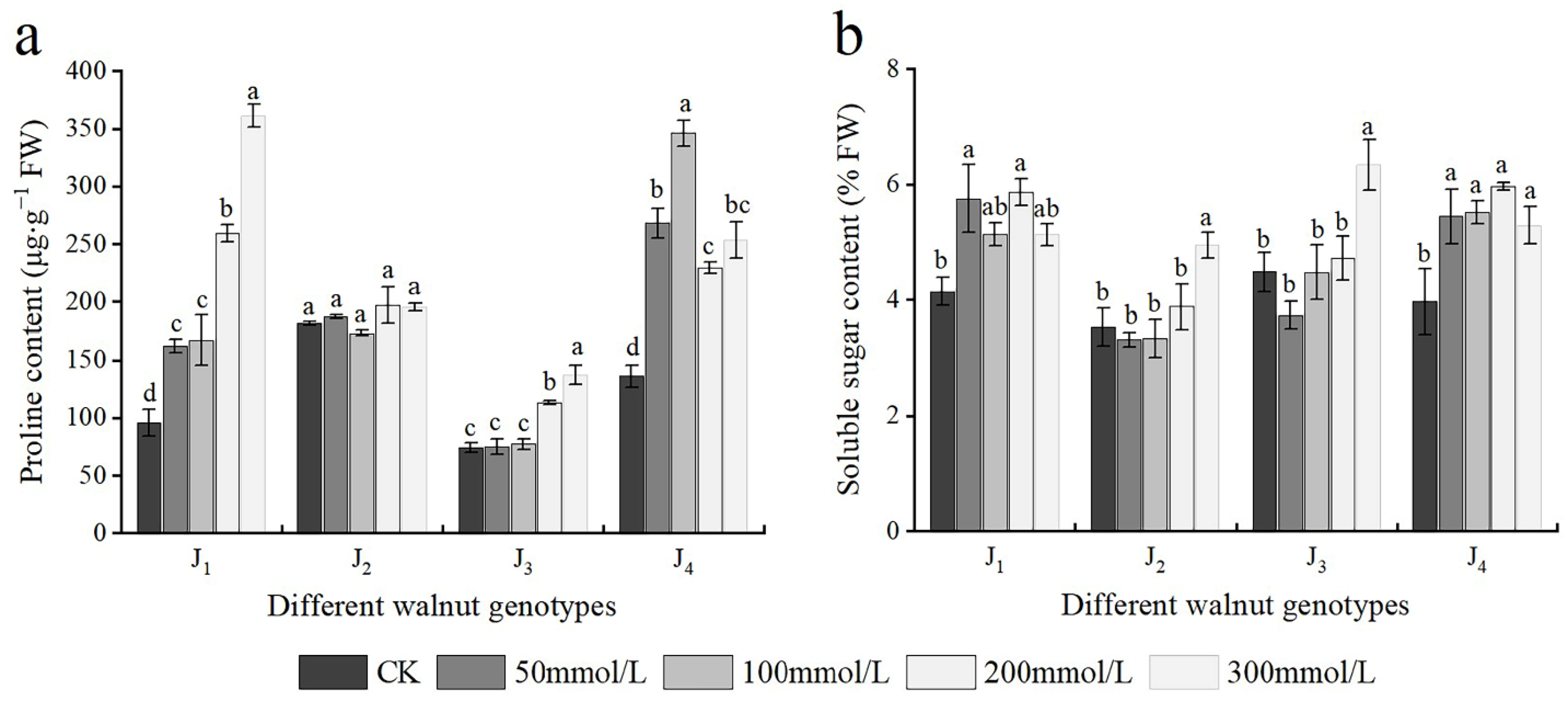
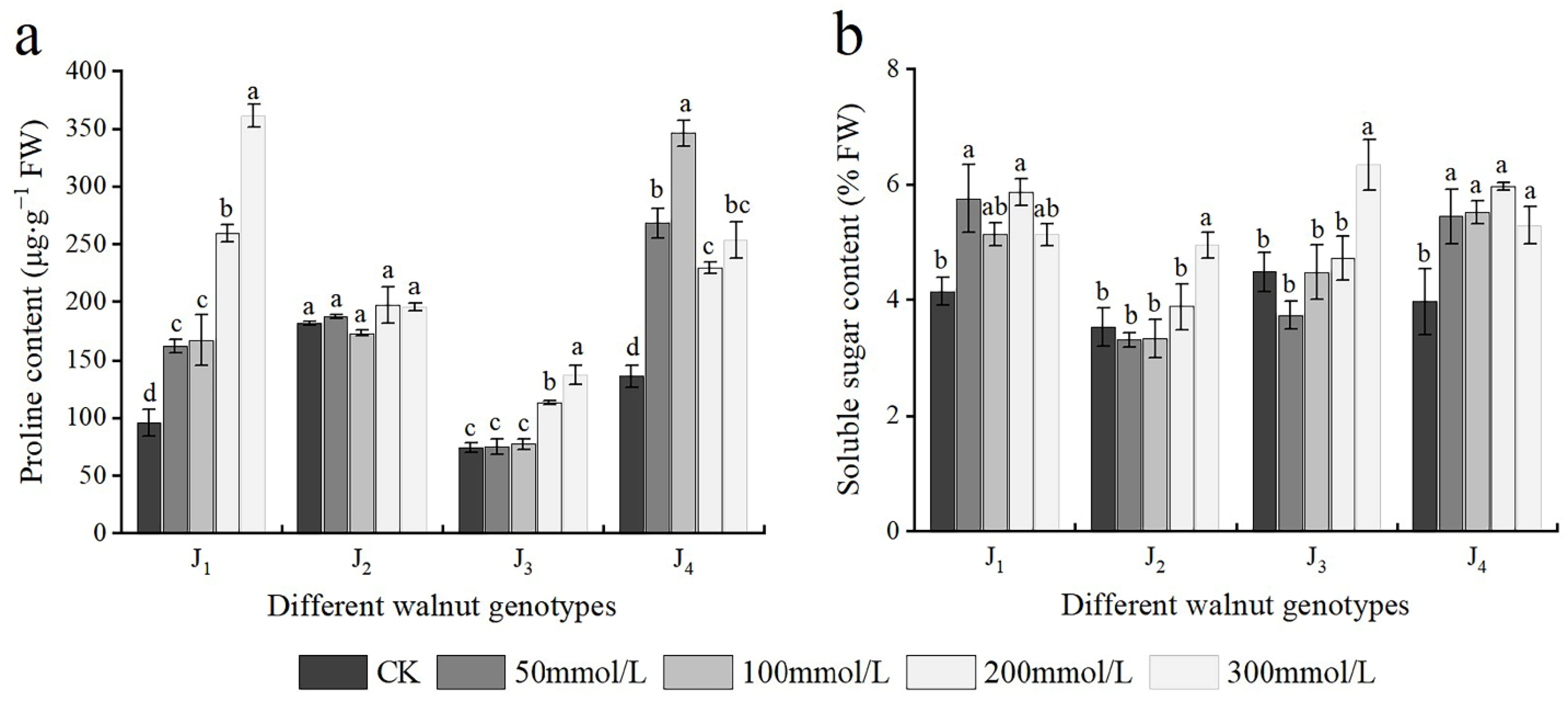
3.6. Effects of Salt Stress on Osmotic Adjustment Substances
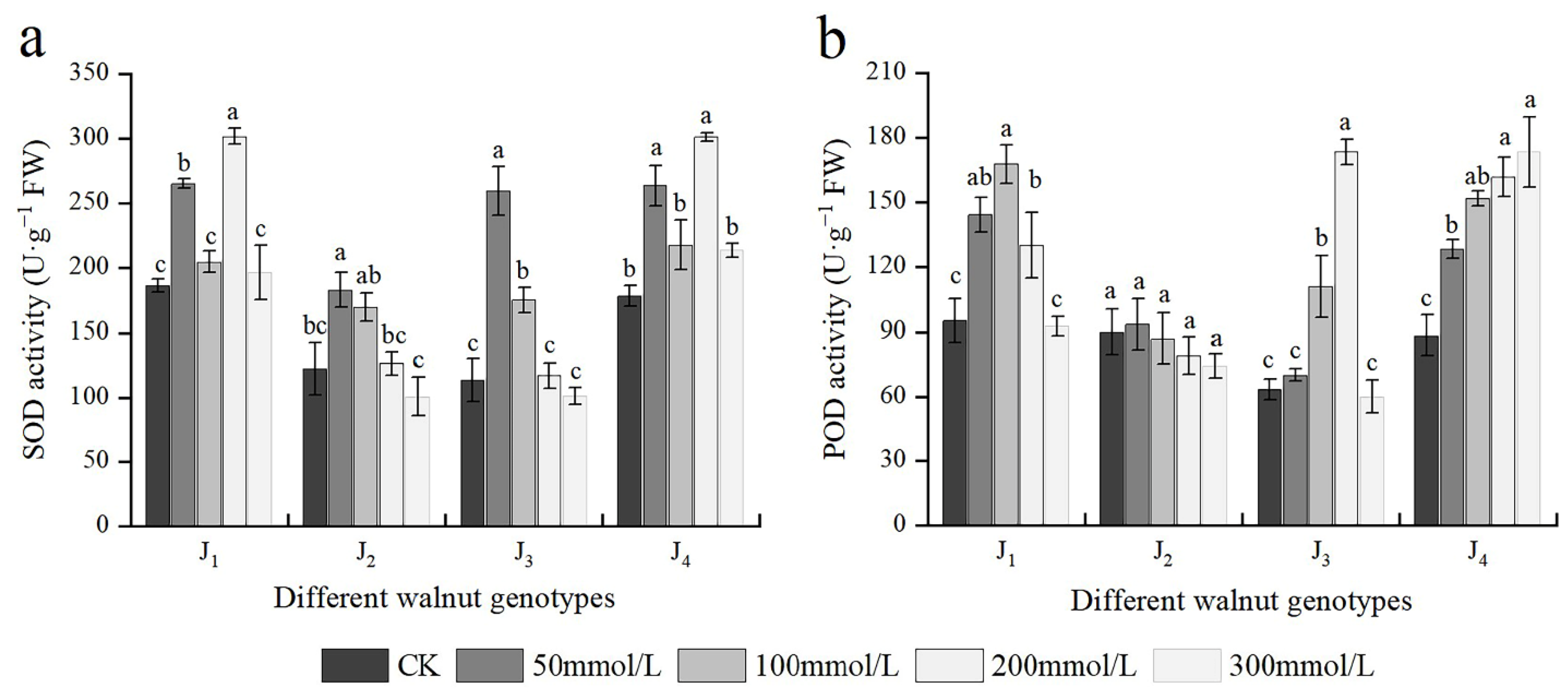
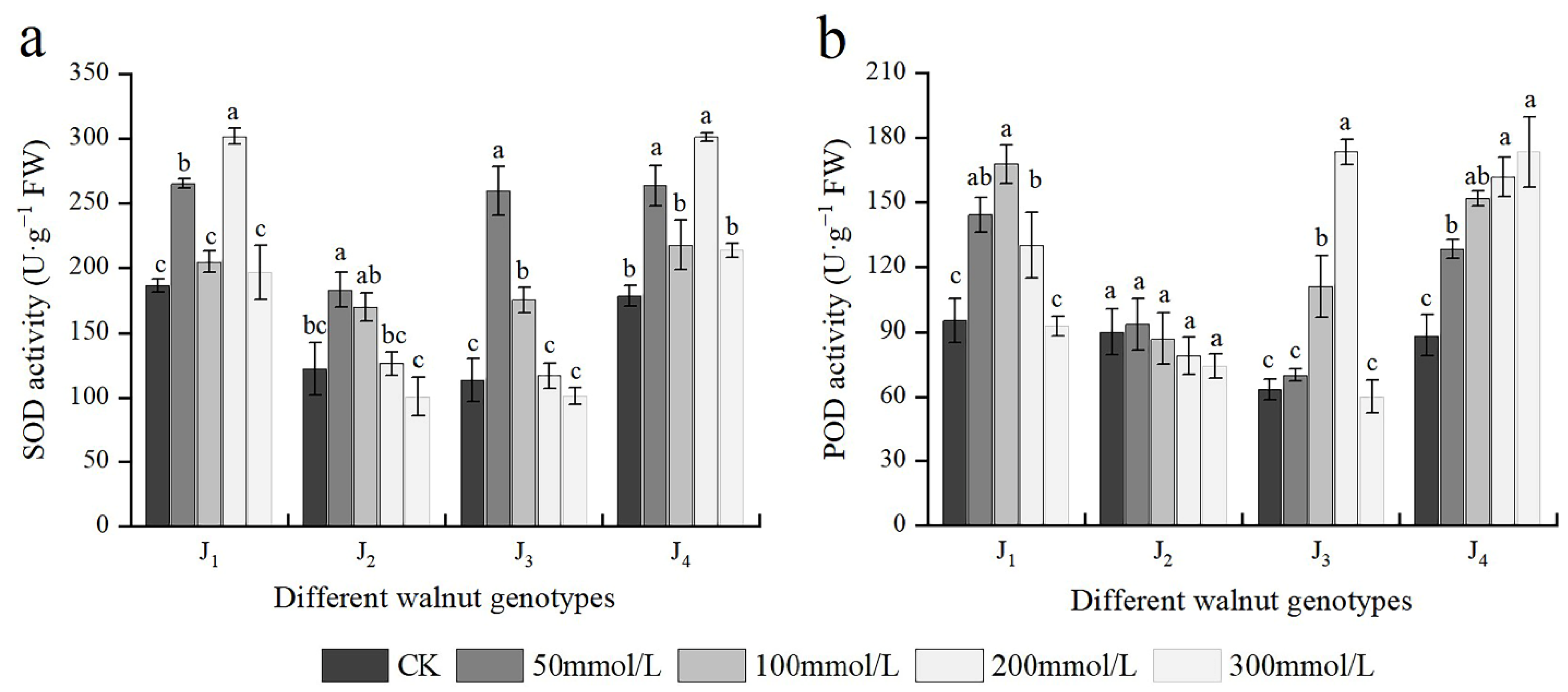
3.7. Effects of Salt Stress on Antioxidant Enzyme Activities
3.8. Comprehensive Evaluation of Salt Tolerance
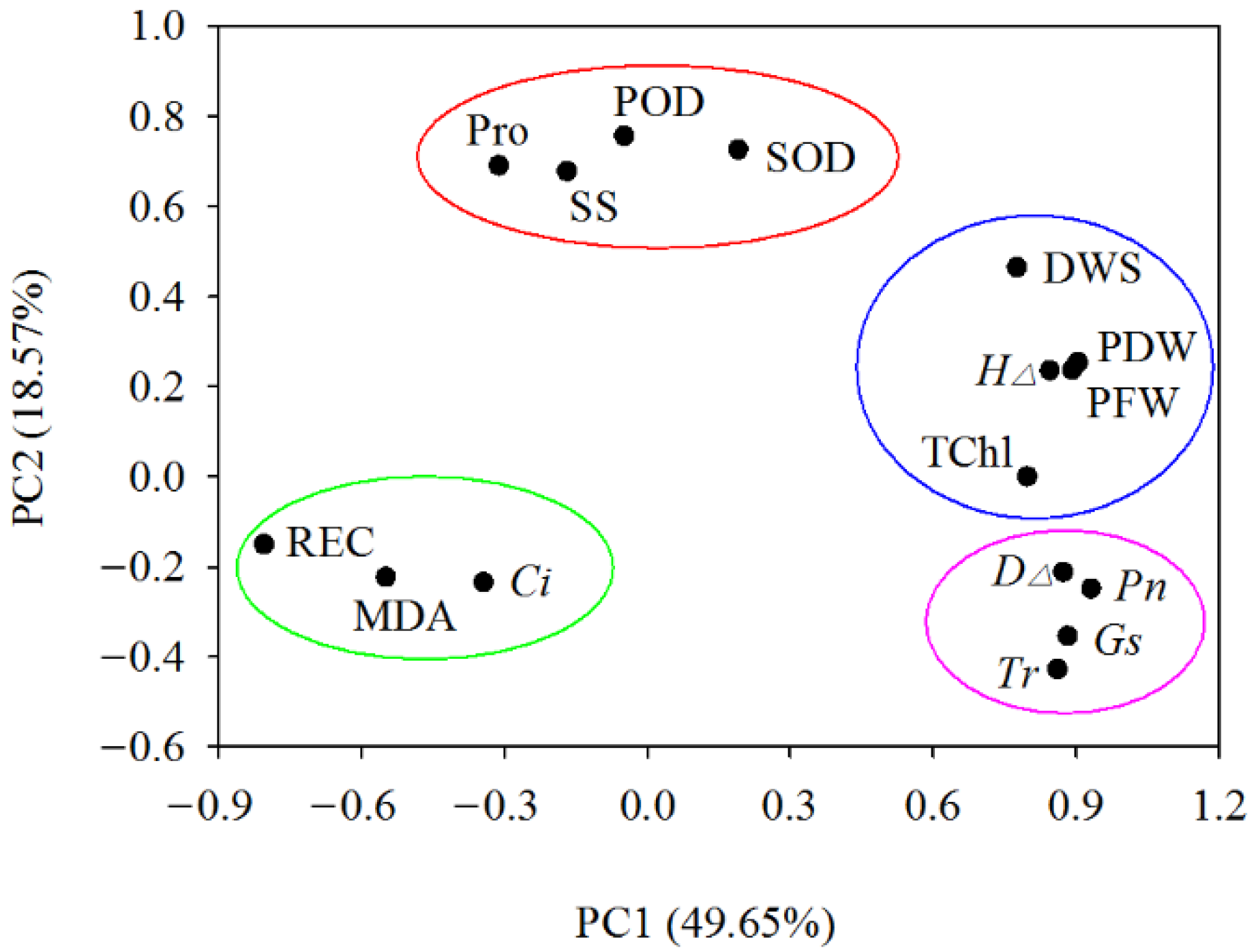
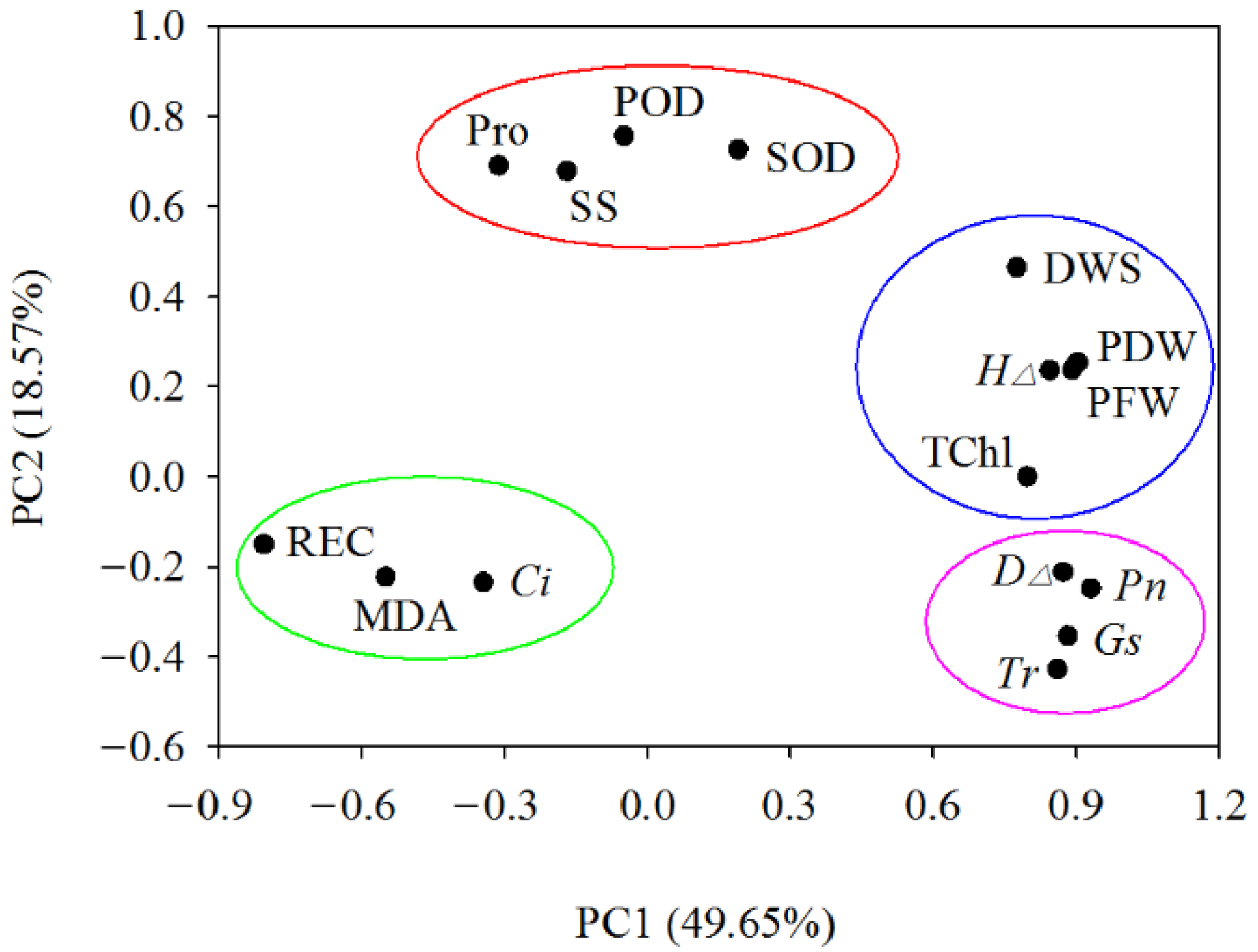
3.8.1. Principal Component Analysis (PCA)
3.8.2. Membership Function Analysis
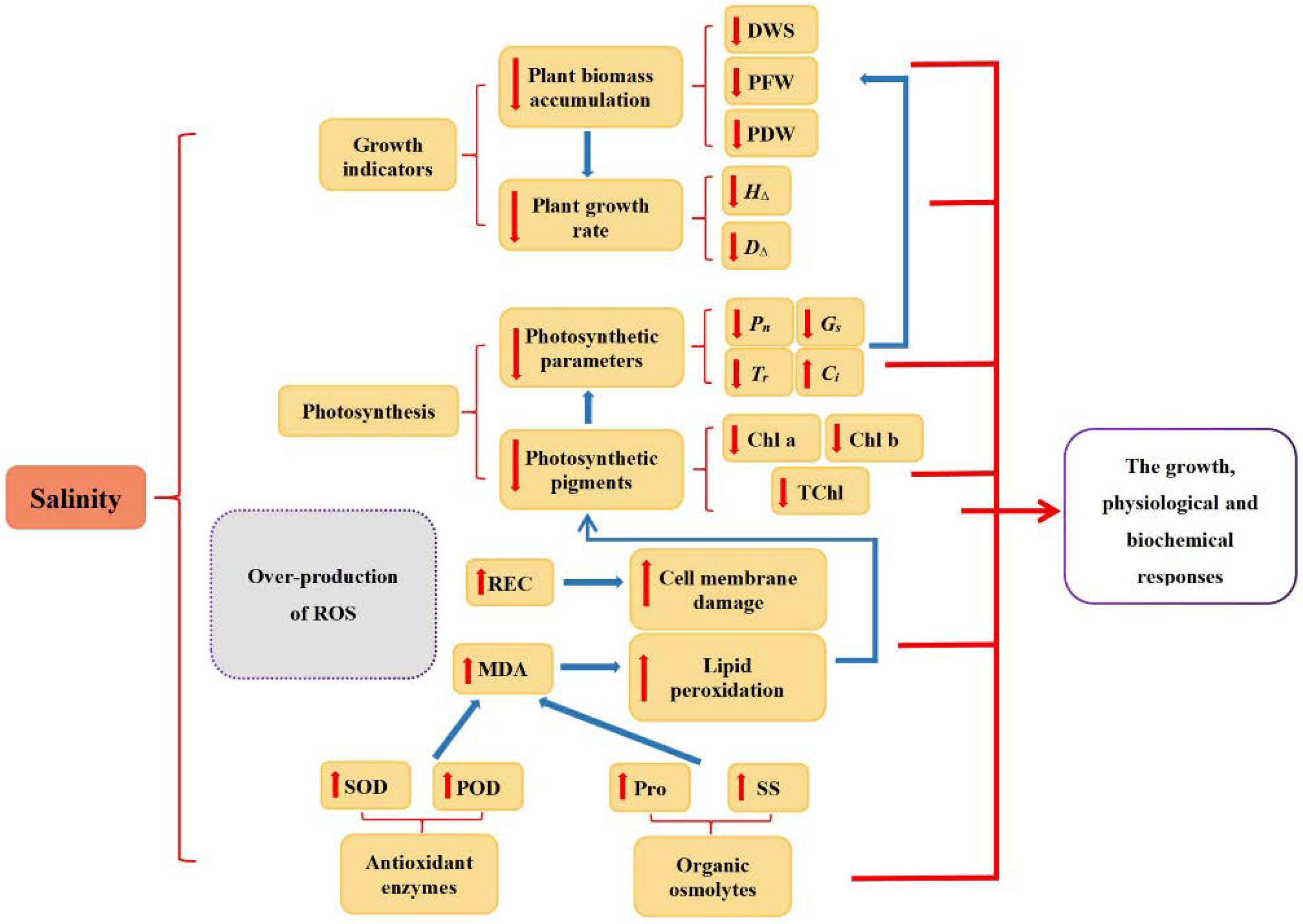
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, X.; Yang, Y.; Wei, H.; Yuan, Z. Soil microbiome-mediated salinity tolerance in poplar plantlets is source-dependent. Chemosphere 2021, 272, 129600. [Google Scholar] [CrossRef] [PubMed]
- Ci, L.J.; Yang, X.H. Desertification and Its Control in China; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 9783642018695. [Google Scholar]
- Libutti, A.; Cammerino, A.; Monteleone, M. Risk assessment of soil salinization due to tomato cultivation in Mediterranean climate conditions. Water 2018, 10, 1503. [Google Scholar] [CrossRef] [Green Version]
- Kulczyk-Skrzeszewska, M.; Kieliszewska-Rokicka, B. Influence of drought and salt stress on the growth of young Populus nigra ‘Italica’ plants and associated mycorrhizal fungi and non-mycorrhizal fungal endophytes. New For. 2021, 1–16. [Google Scholar] [CrossRef]
- Toro, G.; Pimentel, P.; Salvatierra, A. Effective categorization of tolerance to salt stress through clustering Prunus rootstocks according to their physiological performances. Horticulturae 2021, 7, 542. [Google Scholar] [CrossRef]
- Liu, C.; Yan, M.; Huang, X.; Yuan, Z. Effects of salt stress on growth and physiological characteristics of pomegranate (Punica granatum L.) cuttings. Pak. J. Bot. 2018, 50, 457–464. [Google Scholar]
- Capula-Rodríguez, R.; Valdez-Aguilar, L.A.; Cartmill, D.L.; Cartmill, A.D.; Alia-Tejacal, I. Supplementary Calcium and Potassium Improve the Response of Tomato (Solanum lycopersicum L.) to Simultaneous Alkalinity, Salinity, and Boron Stress. Commun. Soil Sci. Plant Anal. 2016, 47, 505–511. [Google Scholar] [CrossRef]
- Chen, W.; Cui, P.; Sun, H.; Guo, W.; Yang, C.; Jin, H.; Fang, B.; Shi, D. Comparative effects of salt and alkali stresses on organic acid accumulation and ionic balance of seabuckthorn (Hippophae rhamnoides L.). Ind. Crops Prod. 2009, 30, 351–358. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Abid, M.; Zhang, Y.J.; Li, Z.; Bai, D.F.; Zhong, Y.P.; Fang, J.B. Effect of Salt stress on growth, physiological and biochemical characters of Four kiwifruit genotypes. Sci. Hortic. 2020, 271, 109473. [Google Scholar] [CrossRef]
- Zhong, Y.P.; Qi, X.J.; Chen, J.Y.; Li, Z.; Bai, D.F.; Wei, C.G.; Fang, J.B. Growth and physiological responses of four kiwifruit genotypes to salt stress and resistance evaluation. J. Integr. Agric. 2019, 18, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Fu, Y.; Hu, D.; Yu, J.; Liu, H. Effect of green, yellow and purple radiation on biomass, photosynthesis, morphology and soluble sugar content of leafy lettuce via spectral wavebands “knock out”. Sci. Hortic. 2018, 236, 10–17. [Google Scholar] [CrossRef]
- Li, Q.; Lv, L.R.; Teng, Y.J.; Si, L.B.; Ma, T.; Yang, Y.L. Apoplastic hydrogen peroxide and superoxide anion exhibited different regulatory functions in salt-induced oxidative stress in wheat leaves. Biol. Plant. 2018, 62, 750–762. [Google Scholar] [CrossRef]
- Liu, A.; Hu, Z.; Bi, A.; Fan, J.; Gitau, M.M.; Amombo, E.; Chen, L.; Fu, J. Photosynthesis, antioxidant system and gene expression of bermudagrass in response to low temperature and salt stress. Ecotoxicology 2016, 25, 1445–1457. [Google Scholar] [CrossRef]
- Shahid, M.A.; Balal, R.M.; Khan, N.; Simón-Grao, S.; Alfosea-Simón, M.; Cámara-Zapata, J.M.; Mattson, N.S.; Garcia-Sanchez, F. Rootstocks influence the salt tolerance of Kinnow mandarin trees by altering the antioxidant defense system, osmolyte concentration, and toxic ion accumulation. Sci. Hortic. 2019, 250, 1–11. [Google Scholar] [CrossRef]
- Cha-Um, S.; Kirdmanee, K. Effect of salt stress on proline accumulation, photosynthetic ability and growth characters in two maize cultivars. Pak. J. Bot. 2009, 41, 87–98. [Google Scholar]
- Zhang, W.; Zwiazek, J.J. Effects of root medium pH on root water transport and apoplastic pH in red-osier dogwood (Cornus sericea) and paper birch (Betula papyrifera) seedlings. Plant Biol. 2016, 18, 1001–1007. [Google Scholar] [CrossRef]
- Zhu, S.; Nong, J.; Luo, G.; Li, Q.; Wang, F.; Jiang, D.; Zhao, X. Varied tolerance and different responses of five citrus rootstocks to acid stress by principle component analysis and orthogonal analysis. Sci. Hortic. 2021, 278, 109853. [Google Scholar] [CrossRef]
- Karimi, S.; Karami, H.; Vahdati, K.; Mokhtassi-Bidgoli, A. Antioxidative responses to short-term salinity stress induce drought tolerance in walnut. Sci. Hortic. 2020, 267, 109322. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, J.; Pei, D.; Yu, L. Combined effects of water stress and salinity on growth, physiological, and biochemical traits in two walnut genotypes. Physiol. Plant. 2021, 172, 176–187. [Google Scholar] [CrossRef]
- Song, D.; Pan, K.; Zhang, A.; Wu, X.; Tariq, A.; Chen, W.; Li, Z.; Sun, F.; Sun, X.; Olatunji, O.A.; et al. Optimization of growth and production parameters of walnut (Juglans regia) saplings with response surface methodology. Sci. Rep. 2018, 8, 9992. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Mei, L.; Jiang, H.; Hu, G.; Wang, Y. Evaluation of Carya cathayensis resistance to Botryosphaeria trunk canker using grafting on pecan. Sci. Hortic. 2019, 248, 184–188. [Google Scholar] [CrossRef]
- Meng, B.N.; Zhang, J.P.; Pei, D.; Xu, H.Z.; Wang, S.M.; Guo, Z.M. Effects of different rootstocks on photosynthesis characteristics in walnut. Nonwood For. Res. 2013, 2. [Google Scholar]
- Hao, L.C.; Wang, Y.Y.; Qi, G.H.; Zhang, X.M.; Li, B.G. Effect of rootstocks on the quality of nut of ‘Lyuling’ walnut. North. Hortic. 2015, 10, 29–32. [Google Scholar]
- Ebrahimi, A.; Zarei, A.; McKenna, J.R.; Bujdoso, G.; Woeste, K.E. Genetic diversity of Persian walnut (Juglans regia) in the cold-temperate zone of the United States and Europe. Sci. Hortic. 2017, 220, 36–41. [Google Scholar] [CrossRef]
- Aslamarz, A.A.; Vahdati, K.; Hassani, D.; Rahemi, M.; Mohammadi, N.; Leslie, C.J. Cold hardiness and its relationship with proline content in Persian walnut. Eur. J. Hortic. Sci. 2011, 76, 84–90. [Google Scholar]
- Yang, G.; Zhang, W.; Liu, Z.; Yi-Maer, A.Y.; Zhai, M.; Xu, Z. Both JrWRKY2 and JrWRKY7 of Juglans regia mediate responses to abiotic stresses and abscisic acid through formation of homodimers and interaction. Plant Biol. 2017, 19, 268–278. [Google Scholar] [CrossRef]
- Esfandiari Ghalati, R.; Shamili, M.; Homaei, A. Effect of putrescine on biochemical and physiological characteristics of guava (Psidium guajava L.) seedlings under salt stress. Sci. Hortic. 2020, 261, 108961. [Google Scholar] [CrossRef]
- Wang, J.; Qiao, Q.; Tao, J. The Physiological Response of Three Narcissus pseudonarcissus under NaCl Stress. Am. J. Plant Sci. 2019, 10, 447–461. [Google Scholar] [CrossRef] [Green Version]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Pan, J.; Zhang, M.; Xing, X.; Zhou, Y.; Liu, Y.; Li, D.; Li, D. ZmMKK4, a novel group C mitogen-activated protein kinase kinase in maize (Zea mays), confers salt and cold tolerance in transgenic Arabidopsis. Plant Cell Environ. 2011, 34, 1291–1303. [Google Scholar] [CrossRef]
- Khalid, M.F.; Hussain, S.; Anjum, M.A.; Ahmad, S.; Ali, M.A.; Ejaz, S.; Morillon, R. Better salinity tolerance in tetraploid vs. diploid volkamer lemon seedlings is associated with robust antioxidant and osmotic adjustment mechanisms. J. Plant Physiol. 2020, 244, 153071. [Google Scholar] [CrossRef]
- Muñoz-Muñoz, J.L.; García-Molina, F.; García-Ruiz, P.A.; Arribas, E.; Tudela, J.; García-Cánovas, F.; Rodríguez-López, J.N. Enzymatic and chemical oxidation of trihydroxylated phenols. Food Chem. 2009, 113, 435–444. [Google Scholar] [CrossRef]
- Urbinati, G.; Nota, P.; Frattarelli, A.; Lucioli, S.; Forni, C.; Caboni, E. Morpho-physiological responses of sea buckthorn (Hippophae rhamnoides) to NaCl stress. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2019, 154, 827–834. [Google Scholar] [CrossRef]
- Mao, P.; Zhang, Y.; Cao, B.; Guo, L.; Shao, H.; Cao, Z.; Jiang, Q.; Wang, X. Effects of salt stress on eco-physiological characteristics in Robinia pseudoacacia based on salt-soil rhizosphere. Sci. Total Environ. 2016, 568, 118–123. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Diaz-Vivancos, P.; Alvarez, S.; Fernandez-Garcia, N.; Sanchez-Blanco, M.J.; Hernandez, J.A. NaCl-induced physiological and biochemical adaptative mechanisms in the ornamental Myrtus communis L. plants. J. Plant Physiol. 2015, 183, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.J.; Uddin, M.J.; Hossain, M.A.; Henry, R.; Begum, M.K.; Sohel, M.A.T.; Mou, M.A.; Ahn, J.; Cheong, E.J.; Lim, Y.S. Exogenous putrescine attenuates the negative impact of drought stress by modulating physio-biochemical traits and gene expression in sugar beet (Beta vulgaris L.). PLoS ONE 2022, 17, e0262099. [Google Scholar] [CrossRef]
- Regni, L.; Del Pino, A.M.; Mousavi, S.; Palmerini, C.A.; Baldoni, L.; Mariotti, R.; Mairech, H.; Gardi, T.; D’Amato, R.; Proietti, P. Behavior of four olive cultivars during salt stress. Front. Plant Sci. 2019, 10, 867. [Google Scholar] [CrossRef]
- Moya, J.L.; Tadeo, F.R.; Gómez-Cadenas, A.; Primo-Millo, E.; Talón, M. Transmissible salt tolerance traits identified through reciprocal grafts between sensitive Carrizo and tolerant Cleopatra citrus genotypes. J. Plant Physiol. 2002, 159, 991–998. [Google Scholar] [CrossRef]
- Khoshbakht, D.; Ramin, A.A.; Baninasab, B. Effects of sodium chloride stress on gas exchange, chlorophyll content and nutrient concentrations of nine citrus rootstocks. Photosynthetica 2015, 53, 241–249. [Google Scholar] [CrossRef]
- Ahmad, P.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; John, R.; Egamberdieva, D.; Gucel, S. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L.) through antioxidative defense system. Front. Plant Sci. 2015, 6, 868. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.; Shao, H.; Shao, C.; Chen, P.; Zhao, S.; Brestic, M.; Chen, X. Physiological adaptive mechanisms of plants grown in saline soil and implications for sustainable saline agriculture in coastal zone. Acta Physiol. Plant. 2013, 35, 2867–2878. [Google Scholar] [CrossRef]
- Mazumdar, P.; Lau, S.E.; Singh, P.; Takhtgahi, H.M.; Harikrishna, J.A. Impact of sea-salt on morpho-physiological and biochemical responses in banana (Musa acuminata cv. Berangan). Physiol. Mol. Biol. Plants 2019, 25, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wang, J.; Yan, D.; Yuan, H.; Wang, Y.; He, Y.; Wang, X.; Li, Z.; Mei, J.; Hu, M.; et al. Exogenous spermidine improves salt tolerance of pecan-grafted seedlings via activating antioxidant system and inhibiting the enhancement of Na+/K+ ratio. Acta Physiol. Plant. 2020, 42, 83. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotoxicol. Environ. Saf. 2017, 137, 64–70. [Google Scholar] [CrossRef]
- Abdallah, M.M.S.; Abdelgawad, Z.A.; El-Bassiouny, H.M.S. Alleviation of the adverse effects of salinity stress using trehalose in two rice varieties. S. Afr. J. Bot. 2016, 103, 275–282. [Google Scholar] [CrossRef]
- Hatami, E.; Shokouhian, A.A.; Ghanbari, A.R.; Naseri, L.A. Alleviating salt stress in almond rootstocks using of humic acid. Sci. Hortic. 2018, 237, 296–302. [Google Scholar] [CrossRef]
- Klein, A.; Hüsselmann, L.; Keyster, M.; Ludidi, N. Exogenous nitric oxide limits salt-induced oxidative damage in maize by altering superoxide dismutase activity. S. Afr. J. Bot. 2018, 115, 44–49. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; Laouane, R.B.; Anli, M.; Boutasknit, A.; Wahbi, S.; Meddich, A. Use of mycorrhizal fungi in improving tolerance of the date palm (Phoenix dactylifera L.) seedlings to salt stress. Sci. Hortic. 2019, 253, 429–438. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, H.S.; Park, S.C.; Ji, C.Y.; Yang, J.W.; Lee, H.U.; Kwak, S.S. Downregulation of swpa4 peroxidase expression in transgenic sweetpotato plants decreases abiotic stress tolerance and reduces stress-related peroxidase expression. Plant Biotechnol. Rep. 2021, 15, 69–76. [Google Scholar] [CrossRef]
- Gökmen, E.; Ceyhan, E. Effects of drought stress on growth parameters, enzyme activities and proline content in chickpea genotypes. Bangladesh J. Bot. 2015, 44, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Chen, M.; Lei; Wang, L. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Oustric, J.; Morillon, R.; Ollitrault, P.; Herbette, S.; Luro, F.; Froelicher, Y.; Tur, I.; Dambier, D.; Giannettini, J.; Berti, L.; et al. Somatic hybridization between diploid Poncirus and Citrus improves natural chilling and light stress tolerances compared with equivalent doubled-diploid genotypes. Trees 2018, 32, 883–895. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl Concentration /(mmol/L) | Morphological Characteristic of Plants | |||
|---|---|---|---|---|
| J1 | J2 | J3 | J4 | |
| 0 (CK) | Normal color of leaves and growth of plant | Normal color of leaves and growth of plant | Normal color of leaves and growth of plant | Normal color of leaves and growth of plant |
| 50 | Few leaves became yellow and withered | Partial leaves became yellow, withered and curled | Few leaves became yellow, withered and curled | Partial leaf margin withered and curled at upper of plant |
| 100 | Partial leaves became yellow, withered and curled | More leaves obvious became yellow, withered and curled | More leaves became yellow, withered and curled | Partial leaves withered and curled at upper of plant |
| 200 | More leaves became yellow, withered and curled, few leaves fallen off | Most leaves withered and curled, and leaves obvious fallen off | Leaves withered and curled seriously, partial leaves fallen off | More leaves withered and curled, few leaves fallen off |
| 300 | More leaves obvious became yellow, withered and curled, partial leaves fallen off | Most leaves withered, curled and fallen off | More leaves obvious withered, curled and fallen off | More leaves obvious withered and curled, partial leaves fallen off |
| Parameters | Main Effects | Interaction | |
|---|---|---|---|
| Genotype(G) | Salinity(S) | G × S | |
| H∆ | 0.000 | 0.000 | 0.350 |
| D∆ | 0.013 | 0.000 | 0.797 |
| DWS | 0.000 | 0.000 | 0.127 |
| PFW | 0.000 | 0.000 | 0.005 |
| PDW | 0.000 | 0.000 | 0.034 |
| Chl a | 0.000 | 0.000 | 0.389 |
| Chl b | 0.000 | 0.000 | 0.914 |
| TChl | 0.000 | 0.000 | 0.632 |
| Pn | 0.000 | 0.000 | 0.000 |
| Gs | 0.000 | 0.000 | 0.000 |
| Tr | 0.000 | 0.000 | 0.000 |
| Ci | 0.000 | 0.000 | 0.000 |
| MDA | 0.000 | 0.000 | 0.000 |
| REC | 0.000 | 0.000 | 0.900 |
| Pro | 0.000 | 0.000 | 0.000 |
| SS | 0.000 | 0.000 | 0.001 |
| SOD | 0.000 | 0.000 | 0.000 |
| POD | 0.000 | 0.000 | 0.000 |
| Index | ||||
|---|---|---|---|---|
| H∆ | 0.594 | 0.182 | 0.259 | 0.660 |
| D∆ | 0.558 | 0.205 | 0.541 | 0.555 |
| DWS | 0.484 | 0.064 | 0.260 | 0.461 |
| PFW | 0.614 | 0.060 | 0.883 | 0.840 |
| PDW | 0.665 | 0.056 | 0.488 | 0.840 |
| TChl | 0.755 | 0.276 | 0.294 | 0.844 |
| Pn | 0.719 | 0.037 | 0.861 | 0.715 |
| Gs | 0.447 | 0.063 | 0.830 | 0.534 |
| Tr | 0.277 | 0.067 | 0.888 | 0.370 |
| Ci | 0.301 | 0.626 | 0.546 | 0.483 |
| REC | 0.571 | 0.392 | 0.198 | 0.760 |
| MDA | 0.835 | 0.592 | 0.361 | 0.699 |
| Pro | 0.581 | 0.545 | 0.040 | 0.743 |
| SS | 0.651 | 0.252 | 0.540 | 0.662 |
| POD | 0.663 | 0.276 | 0.299 | 0.730 |
| SOD | 0.812 | 0.192 | 0.294 | 0.844 |
| Average | 0.595 | 0.236 | 0.452 | 0.668 |
| Rank | 2 | 4 | 3 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, X.; Tang, J.; Fan, W.; Li, B.; Bai, Y.; He, J.; Pei, D.; Zhang, J. Phenotypic Differences and Physiological Responses of Salt Resistance of Walnut with Four Rootstock Types. Plants 2022, 11, 1557. https://doi.org/10.3390/plants11121557
Ji X, Tang J, Fan W, Li B, Bai Y, He J, Pei D, Zhang J. Phenotypic Differences and Physiological Responses of Salt Resistance of Walnut with Four Rootstock Types. Plants. 2022; 11(12):1557. https://doi.org/10.3390/plants11121557
Chicago/Turabian StyleJi, Xinying, Jiali Tang, Wei Fan, Baoxin Li, Yongchao Bai, Junxing He, Dong Pei, and Junpei Zhang. 2022. "Phenotypic Differences and Physiological Responses of Salt Resistance of Walnut with Four Rootstock Types" Plants 11, no. 12: 1557. https://doi.org/10.3390/plants11121557
APA StyleJi, X., Tang, J., Fan, W., Li, B., Bai, Y., He, J., Pei, D., & Zhang, J. (2022). Phenotypic Differences and Physiological Responses of Salt Resistance of Walnut with Four Rootstock Types. Plants, 11(12), 1557. https://doi.org/10.3390/plants11121557