Beer and its Non-Alcoholic Compounds: Role in Pancreatic Exocrine Secretion, Alcoholic Pancreatitis and Pancreatic Carcinoma

Abstract
:1. Introduction
2. Results and Discussion
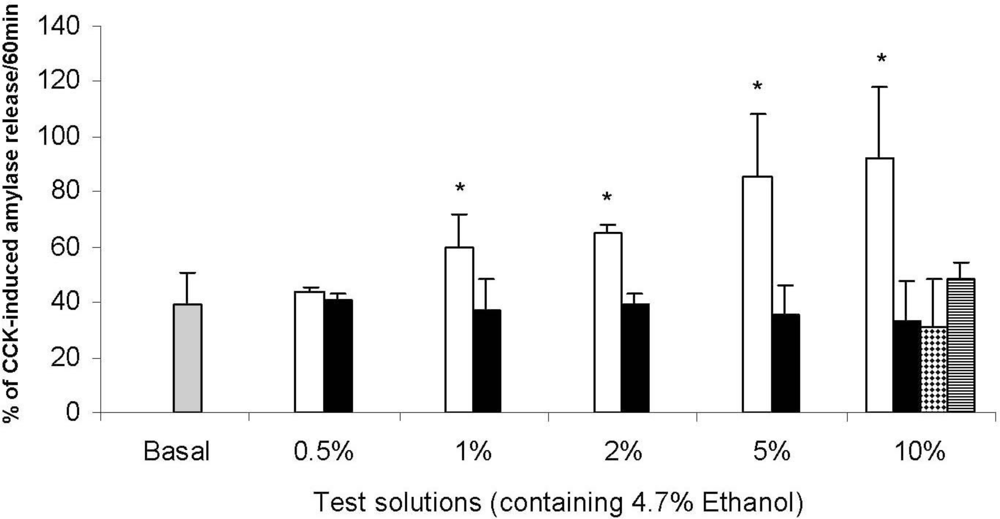
2.1. Beer and Pancreatic Secretion
2.2. Beer and Pancreatitis
2.3. Non-Alcoholic Constituents of Beer and Pancreatitis
2.4. Beer and Carcinogenesis
3. Conclusion
Acknowledgments
References and Notes
- Niebergall-Roth, E; Harder, H; Singer, MV. A review: acute and chronic effects of ethanol and alcoholic beverages on the pancreatic exocrine secretion in vivo and in vitro. Alcohol Clin. Exp. Res 1998, 22, 1570–1583. [Google Scholar]
- Schneider, A; Haas, SL; Singer, MV. Comprehensive Handbook of Alcohol Related Pathology; Preedy, VR, Watson, RR, Eds.; Elsevier Ltd: Amsterdam, The Netherland, 2005; Volume 2, pp. 577–597. [Google Scholar]
- Chari, ST; Harder, H; Teyssen, S; Knodel, C; Riepl, RL; Singer, MV. Effect of beer, yeast-fermented glucose, and ethanol on pancreatic enzyme secretion in healthy human subjects. Dig. Dis. Sci 1996, 41, 1216–1224. [Google Scholar]
- Hajnal, F; Flores, MC; Radley, S; Valenzuela, JE. Effect of alcohol and alcoholic beverages on meal-stimulated pancreatic secretion in humans. Gastroenterology 1990, 98, 191–196. [Google Scholar]
- Hajnal, F; Flores, MC; Valenzuela, JE. Effect of alcohol and alcoholic beverages on nonstimulated pancreatic secretion in humans. Pancreas 1989, 4, 486–491. [Google Scholar]
- Singer, MV; Niebergall-Roth, E. Secretion from acinar cells of the exocrine pancreas: role of enteropancreatic reflexes and cholecystokinin. Cell Biol. Int 2009, 33, 1–9. [Google Scholar]
- Leung, PS. The physiology of a local renin-angiotensin system in the pancreas. J. Physiol 2007, 580, 31–37. [Google Scholar]
- Jaworek, J; Nawrot-Porabka, K; Leja-Szpak, A; Bonior, J; Szklarczyk, J; Kot, M; Konturek, SJ; Pawlik, WW. Melatonin as modulator of pancreatic enzyme secretion and pancreatoprotector. J. Physiol. Pharmacol 2007, 58, 65–80. [Google Scholar]
- Singh, J; Pariente, JA; Salido, GM. The physiological role of histamine in the exocrine pancreas. Inflamm. Res 1997, 46, 159–165. [Google Scholar]
- Chey, WY; Chang, T. Neural hormonal regulation of exocrine pancreatic secretion. Pancreatology 2001, 1, 320–335. [Google Scholar]
- Gerloff, A; Singer, MV; Feick, P. Beer-induced pancreatic enzyme secretion: characterization of some signaling pathways and of the responsible nonalcoholic compounds. Alcohol Clin. Exp. Res 2009, 33, 1638–1645. [Google Scholar]
- Gerloff, A; Singer, MV; Feick, P. Beer but not wine, hard liquors, or pure ethanol stimulates amylase secretion of rat pancreatic acinar cells in vitro. Alcohol Clin. Exp. Res 2009, 33, 1545–1554. [Google Scholar]
- Dietrich, JB. AR4-2J cells: a model to study polypeptide hormone receptors. Biosci. Rep 1996, 16, 273–288. [Google Scholar]
- Spanier, BW; Dijkgraaf, MG; Bruno, MJ. Epidemiology, aetiology and outcome of acute and chronic pancreatitis: An update. Best Pract. Res. Clin. Gastroenterol 2008, 22, 45–63. [Google Scholar]
- Chowdhury, P; Gupta, P. Pathophysiology of alcoholic pancreatitis: an overview. World J. Gastroenterol 2006, 12, 7421–7427. [Google Scholar]
- Ammann, RW. The natural history of alcoholic chronic pancreatitis. Intern. Med 2001, 40, 368–375. [Google Scholar]
- Schneider, A; Whitcomb, DC; Singer, MV. Animal models in alcoholic pancreatitis—what can we learn? Pancreatology 2002, 2, 189–203. [Google Scholar]
- Apte, MV; Pirola, RC; Wilson, JS. Individual susceptibility to alcoholic pancreatitis. J. Gastroenterol. Hepatol 2008, 23, S63–S68. [Google Scholar]
- Apte, MV; Pirola, RC; Wilson, JS. Molecular mechanisms of alcoholic pancreatitis. Dig. Dis 2005, 23, 232–240. [Google Scholar]
- Haber, P; Wilson, J; Apte, M; Korsten, M; Pirola, R. Individual susceptibility to alcoholic pancreatitis: still an enigma. J. Lab. Clin. Med 1995, 125, 305–312. [Google Scholar]
- Wilson, JS; Bernstein, L; McDonald, C; Tait, A; McNeil, D; Pirola, RC. Diet and drinking habits in relation to the development of alcoholic pancreatitis. Gut 1985, 26, 882–887. [Google Scholar]
- Nakamura, T; Ishikawa, A; Sekiguchi, S; Kuroda, M; Imazeki, H; Maruyama, K; Higuchi, S. Spirits Increase the Risk for Chronic Pancreatitis More Than Beer and Sake in 111 Japanese Alcoholics. Gastroenterology 2002, 122, A509. [Google Scholar]
- Nakamura, Y; Kobayashi, Y; Ishikawa, A; Maruyama, K; Higuchi, S. Severe chronic pancreatitis and severe liver cirrhosis have different frequencies and are independent risk factors in male Japanese alcoholics. J. Gastroenterol 2004, 39, 879–887. [Google Scholar]
- Kristiansen, L; Gronbaek, M; Becker, U; Tolstrup, JS. Risk of pancreatitis according to alcohol drinking habits: a population-based cohort study. Am. J. Epidemiol 2008, 168, 932–937. [Google Scholar]
- Piendl, A. Inhaltsstoffe des Bieres, dargestellt am Beispiel des Pilsener Lagerbieres. In Alkohol und Alkoholfolgekrankheiten, 2nd ed; Singer, MV, Teyssen, S, Eds.; Springer Verlag: Berlin, Germany, 2005; pp. 65–69. [Google Scholar]
- Szabolcs, A; Varga, IS; Varga, C; Berko, A; Kaszaki, J; Letoha, T; Tiszlavicz, L; Sari, R; Lonovics, J; Takacs, T. Beneficial effect of resveratrol on cholecystokinin-induced experimental pancreatitis. Eur. J. Pharmacol 2006, 532, 187–193. [Google Scholar]
- Ma, ZH; Ma, QY; Wang, LC; Sha, HC; Wu, SL; Zhang, M. Effect of resveratrol on peritoneal macrophages in rats with severe acute pancreatitis. Inflamm. Res 2005, 54, 522–527. [Google Scholar]
- Meng, Y; Ma, QY; Kou, XP; Xu, J. Effect of resveratrol on activation of nuclear factor kappa-B and inflammatory factors in rat model of acute pancreatitis. World J. Gastroenterol 2005, 11, 525–528. [Google Scholar]
- Wang, L; Ma, Q; Chen, X; Sha, H; Ma, Z. Effects of resveratrol on calcium regulation in rats with severe acute pancreatitis. Eur. J. Pharmacol 2008, 580, 271–276. [Google Scholar]
- Warzecha, Z; Dembinski, A; Ceranowicz, P; Konturek, S; Tomaszewska, R; Stachura, J; Nakamura, T; Konturek, PC. Inhibition of cyclooxygenase-2 reduces the protective effect of hepatocyte growth factor in experimental pancreatitis. Eur. J. Pharmacol 2004, 486, 107–119. [Google Scholar]
- Li, ZD; Ma, QY; Wang, CA. Effect of resveratrol on pancreatic oxygen free radicals in rats with severe acute pancreatitis. World J. Gastroenterol 2006, 12, 137–140. [Google Scholar]
- Lawinski, M; Sledzinski, Z; Kubasik-Juraniec, J; Spodnik, JH; Wozniak, M; Boguslawski, W. Does resveratrol prevent free radical-induced acute pancreatitis? Pancreas 2005, 31, 43–47. [Google Scholar]
- Takabayashi, F; Harada, N; Hara, Y. The effects of green tea catechins (Polyphenon) on DL-ethionine-induced acute pancreatitis. Pancreas 1995, 11, 127–131. [Google Scholar]
- Takabayashi, F; Harada, N. Effects of green tea catechins (Polyphenon 100) on cerulein-induced acute pancreatitis in rats. Pancreas 1997, 14, 276–279. [Google Scholar]
- Takabayashi, F; Harada, N; Tahara, S; Kaneko, T; Hara, Y. Effect of green tea catechins on the amount of 8-hydroxydeoxyguanosine (8-OHdG) in pancreatic and hepatic DNA after a single administration of N-nitrosobis(2-oxopropyl)amine (BOP). Pancreas 1997, 15, 109–112. [Google Scholar]
- Takabayashi, F; Tahara, S; Kaneko, T; Harada, N. Effect of green tea catechins on oxidative DNA damage of hamster pancreas and liver induced by N-Nitrosobis(2-oxopropyl)amine and/or oxidized soybean oil. Biofactors 2004, 21, 335–337. [Google Scholar]
- Bhagat, L; Singh, VP; Hietaranta, AJ; Agrawal, S; Steer, ML; Saluja, AK. Heat shock protein 70 prevents secretagogue-induced cell injury in the pancreas by preventing intracellular trypsinogen activation. J. Clin. Invest 2000, 106, 81–89. [Google Scholar]
- Maxwell, S; Cruickshank, A; Thorpe, G. Red wine and antioxidant activity in serum. Lancet 1994, 344, 193–194. [Google Scholar]
- Apte, MV; Pirola, RC; Wilson, JS. Battle-scarred pancreas: Role of alcohol and pancreatic stellate cells in pancreatic fibrosis. J. Gastroenterol. Hepatol 2006, 21, S97–S101. [Google Scholar]
- Masamune, A; Satoh, M; Kikuta, K; Suzuki, N; Satoh, K; Shimosegawa, T. Ellagic acid blocks activation of pancreatic stellate cells. Biochem. Pharmacol 2005, 70, 869–878. [Google Scholar]
- Go, VL; Gukovskaya, A; Pandol, SJ. Alcohol and pancreatic cancer. Alcohol 2005, 35, 205–211. [Google Scholar]
- Genkinger, JM; Spiegelman, D; Anderson, KE; Bergkvist, L; Bernstein, L; van den Brandt, PA; English, DR; Freudenheim, JL; Fuchs, CS; Giles, GG; Giovannucci, E; Hankinson, SE; Horn-Ross, PL; Leitzmann, M; Mannisto, S; Marshall, JR; McCullough, ML; Miller, AB; Reding, DJ; Robien, K; Rohan, TE; Schatzkin, A; Stevens, VL; Stolzenberg-Solomon, RZ; Verhage, BA; Wolk, A; Ziegler, RG; Smith-Warner, SA. Alcohol intake and pancreatic cancer risk: a pooled analysis of fourteen cohort studies. Cancer Epidemiol. Biomarkers Prev 2009, 18, 765–776. [Google Scholar]
- Lowenfels, AB; Maisonneuve, P; Cavallini, G; Ammann, RW; Lankisch, PG; Andersen, JR; Dimagno, EP; Andren-Sandberg, A; Domellof, L. Pancreatitis and the risk of pancreatic cancer. International Pancreatitis Study Group. N. Engl. J. Med 1993, 328, 1433–1437. [Google Scholar]
- Nothlings, U; Murphy, SP; Wilkens, LR; Henderson, BE; Kolonel, LN. Flavonols and pancreatic cancer risk: the multiethnic cohort study. Am. J. Epidemiol 2007, 166, 924–931. [Google Scholar]
- Mouria, M; Gukovskaya, AS; Jung, Y; Buechler, P; Hines, OJ; Reber, HA; Pandol, SJ. Food-derived polyphenols inhibit pancreatic cancer growth through mitochondrial cytochrome C release and apoptosis. Int. J. Cancer 2002, 98, 761–769. [Google Scholar]
- Edderkaoui, M; Odinokova, I; Ohno, I; Gukovsky, I; Go, VL; Pandol, SJ; Gukovskaya, AS. Ellagic acid induces apoptosis through inhibition of nuclear factor kappa B in pancreatic cancer cells. World J. Gastroenterol 2008, 14, 3672–3680. [Google Scholar]
- Ding, XZ; Adrian, TE. Resveratrol inhibits proliferation and induces apoptosis in human pancreatic cancer cells. Pancreas 2002, 25, e71–e76. [Google Scholar]
- Yamagishi, M; Natsume, M; Osakabe, N; Nakamura, H; Furukawa, F; Imazawa, T; Nishikawa, A; Hirose, M. Effects of cacao liquor proanthocyanidins on PhIP-induced mutagenesis in vitro, and in vivo mammary and pancreatic tumorigenesis in female Sprague-Dawley rats. Cancer Lett 2002, 185, 123–130. [Google Scholar]
- Sun, W; Wang, W; Kim, J; Keng, P; Yang, S; Zhang, H; Liu, C; Okunieff, P; Zhang, L. Anti-cancer effect of resveratrol is associated with induction of apoptosis via a mitochondrial pathway alignment. Adv. Exp. Med. Biol 2008, 614, 179–186. [Google Scholar]


{kind=link}
| Compound | Experimental Design | Effect on Pancreas | Species | Reference |
|---|---|---|---|---|
| Quercetin | in vivo 1.3 mg daily, initiated 10 days after tumor transplantation | Decrease of the primary tumor growth Prevention of metastatic lesion | Nude Mouse | [45] |
| in vitro <100 μM | Induction of apoptosis by mitochondrial depolarization, cytochrome c release and caspase activation | Rat (BSp73AS cell line) Human (Mia PACA-2 cell line) | [45] | |
| Ellagic acid | in vitro 10–50 mM | Induction of apoptosis by mitochondrial depolarization, cytochrome c release and caspase activation Decrease of NF-kB binding activity | Human (Mia PACA-2 and PANC-1 cell line) | [46] |
| Resveratrol | in vitro <100 μM | Induction of apoptosis by mitochondrial depolarization, cytochrome c release and caspase activation | Rat (BSp73AS cell line) Human (Mia PACA-2 cell line) | [45] |
| in vitro 100 μM | Inhibition of proliferation and induction of apoptosis Change of cell cycle distribution | Human (PANC-1 and AsPC-1 cell line) | [47] | |
| Proanthocyanidins | in vivo 0.025 and 0.25% fed during or after experimentally induced tumor growth | Inhibition of rat pancreatic carcinogenesis in the initiation stage | Rat (Sprague-Dawley) | [48] |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gerloff, A.; Singer, M.V.; Feick, P. Beer and its Non-Alcoholic Compounds: Role in Pancreatic Exocrine Secretion, Alcoholic Pancreatitis and Pancreatic Carcinoma. Int. J. Environ. Res. Public Health 2010, 7, 1093-1104. https://doi.org/10.3390/ijerph7031093
Gerloff A, Singer MV, Feick P. Beer and its Non-Alcoholic Compounds: Role in Pancreatic Exocrine Secretion, Alcoholic Pancreatitis and Pancreatic Carcinoma. International Journal of Environmental Research and Public Health. 2010; 7(3):1093-1104. https://doi.org/10.3390/ijerph7031093
Chicago/Turabian StyleGerloff, Andreas, Manfred V. Singer, and Peter Feick. 2010. "Beer and its Non-Alcoholic Compounds: Role in Pancreatic Exocrine Secretion, Alcoholic Pancreatitis and Pancreatic Carcinoma" International Journal of Environmental Research and Public Health 7, no. 3: 1093-1104. https://doi.org/10.3390/ijerph7031093
APA StyleGerloff, A., Singer, M. V., & Feick, P. (2010). Beer and its Non-Alcoholic Compounds: Role in Pancreatic Exocrine Secretion, Alcoholic Pancreatitis and Pancreatic Carcinoma. International Journal of Environmental Research and Public Health, 7(3), 1093-1104. https://doi.org/10.3390/ijerph7031093