
Long-Term Impact of Childhood Adversity on the Gut Microbiome of Nursing Students

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Participant Recruitment
2.2. Stool Collection and Analyses
2.3. Blood Collection and Analysis
2.4. Demographic and Collected Health Endpoints
2.5. Childhood Adversity
2.6. Depression Questionnaires
2.7. Statistical Analysis
3. Results
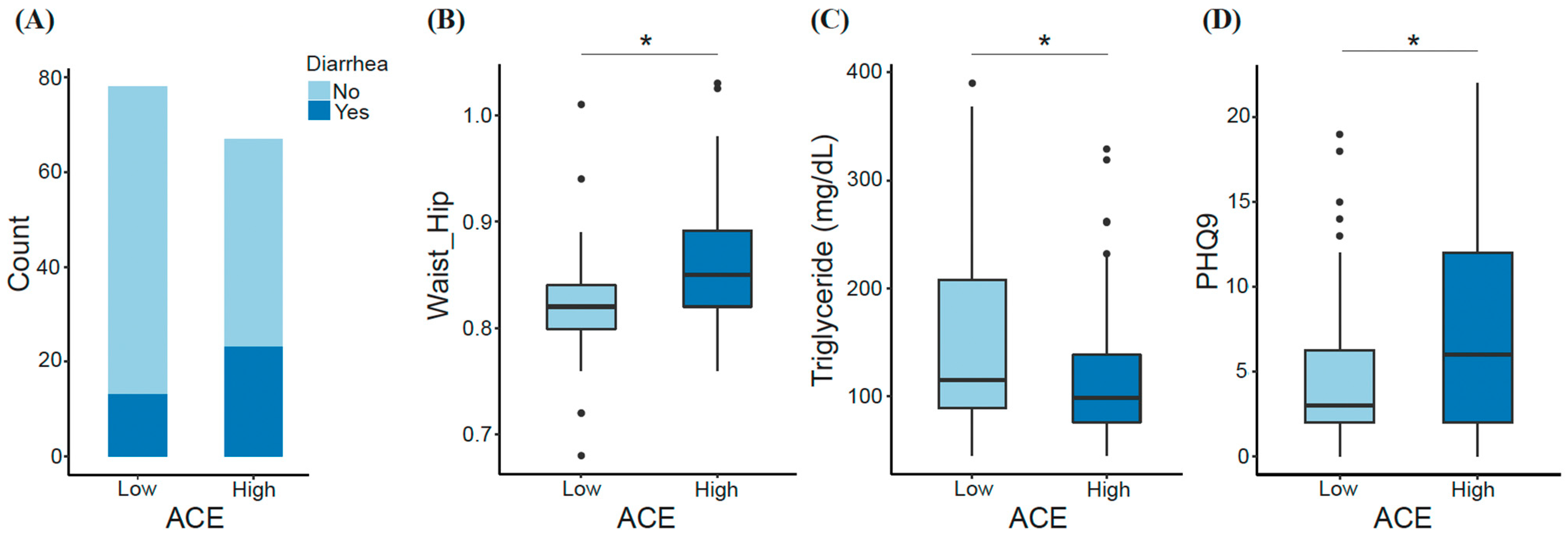
3.1. Demographic Characteristics and Features Differentiating ACE Groups
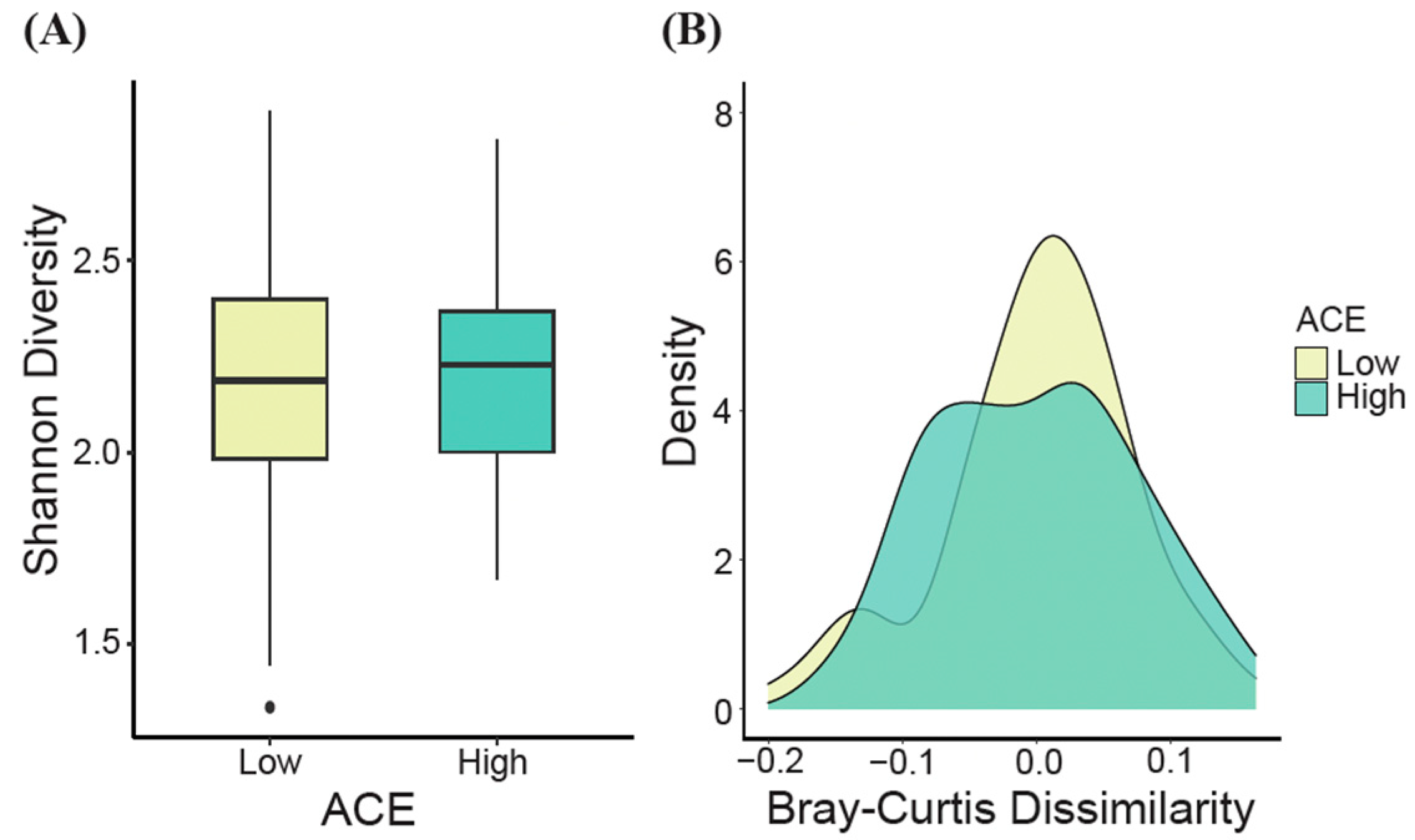
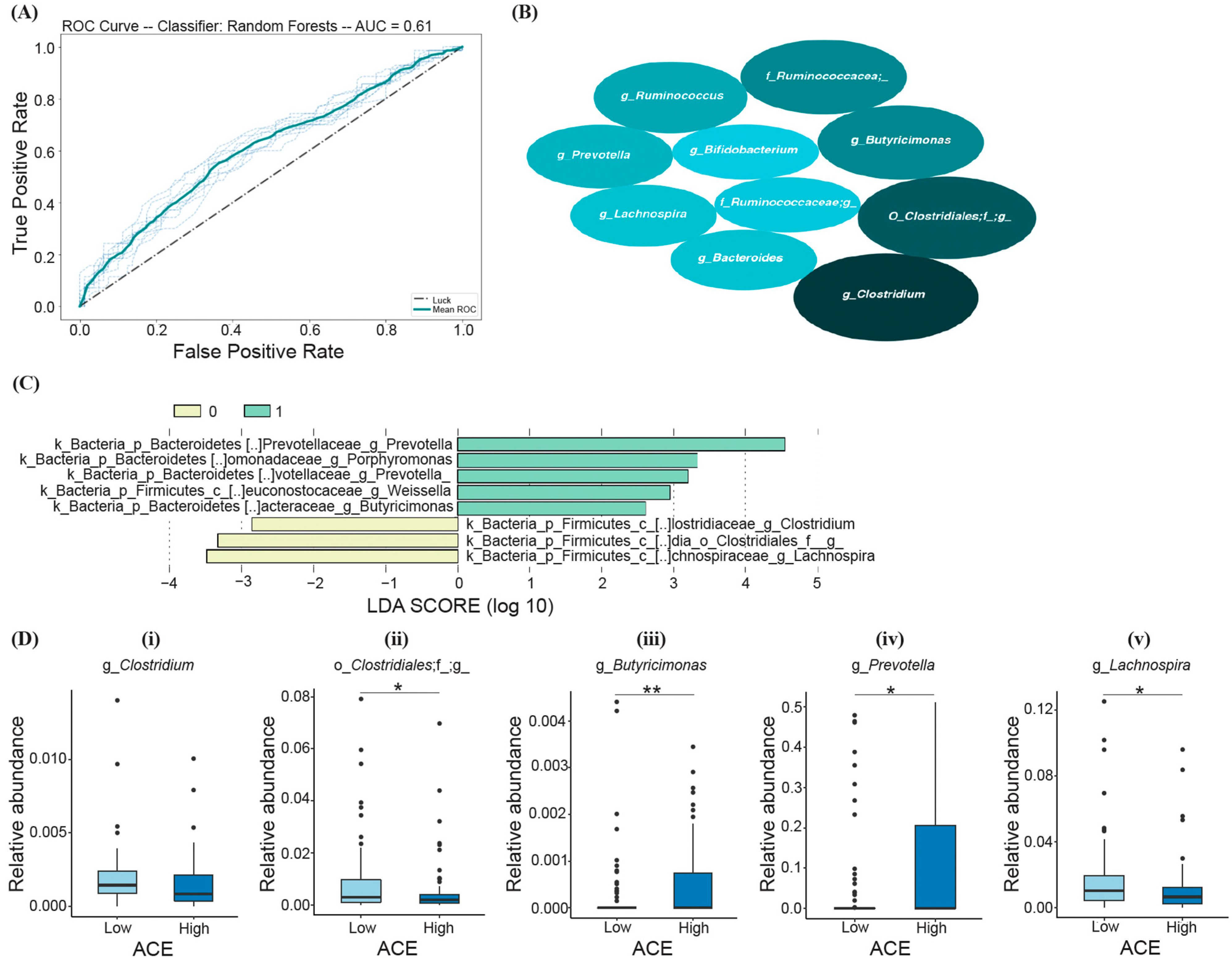
3.2. Gut Microbiome Diversity Analysis
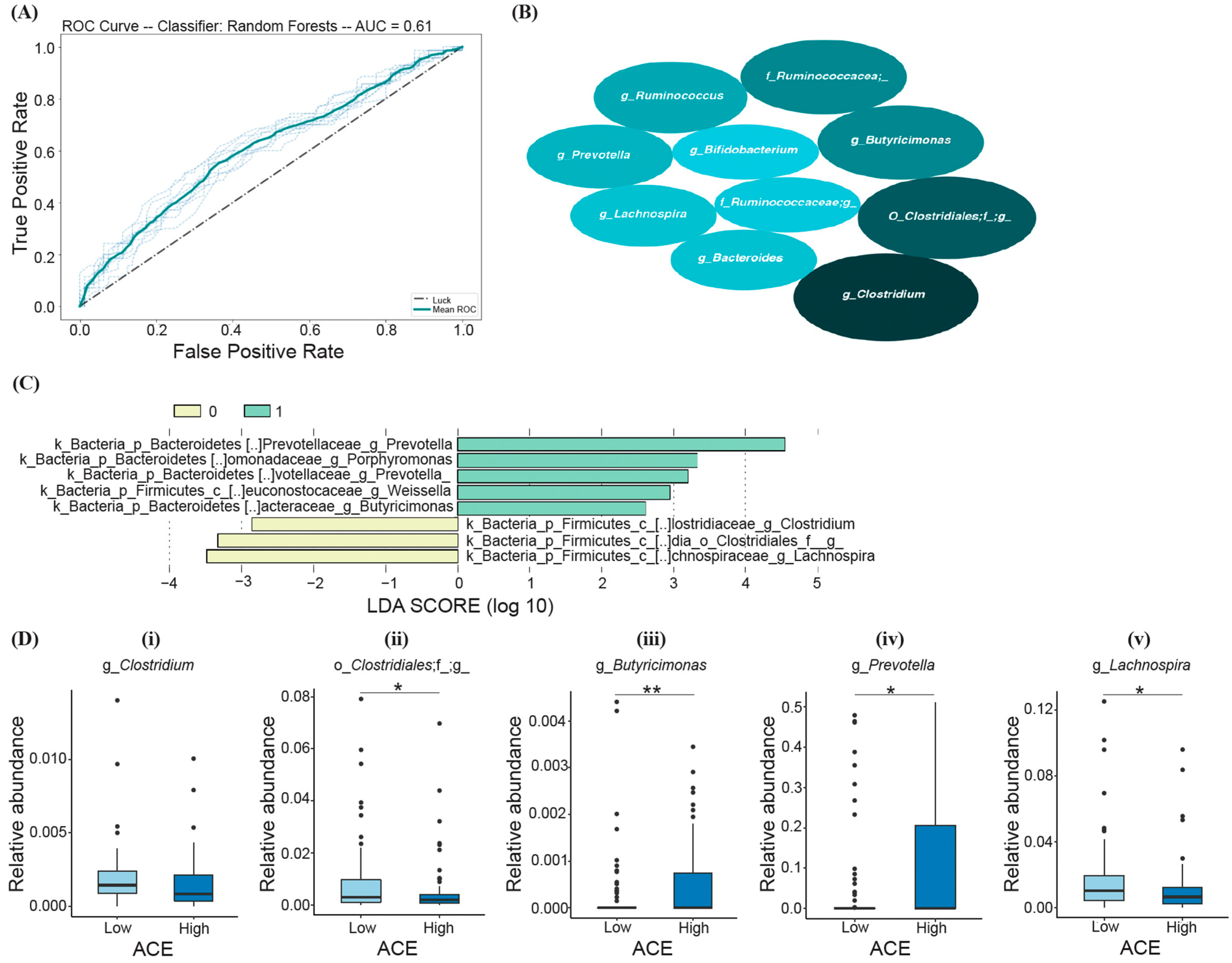
3.3. Bacteria Differentiating ACE Groups
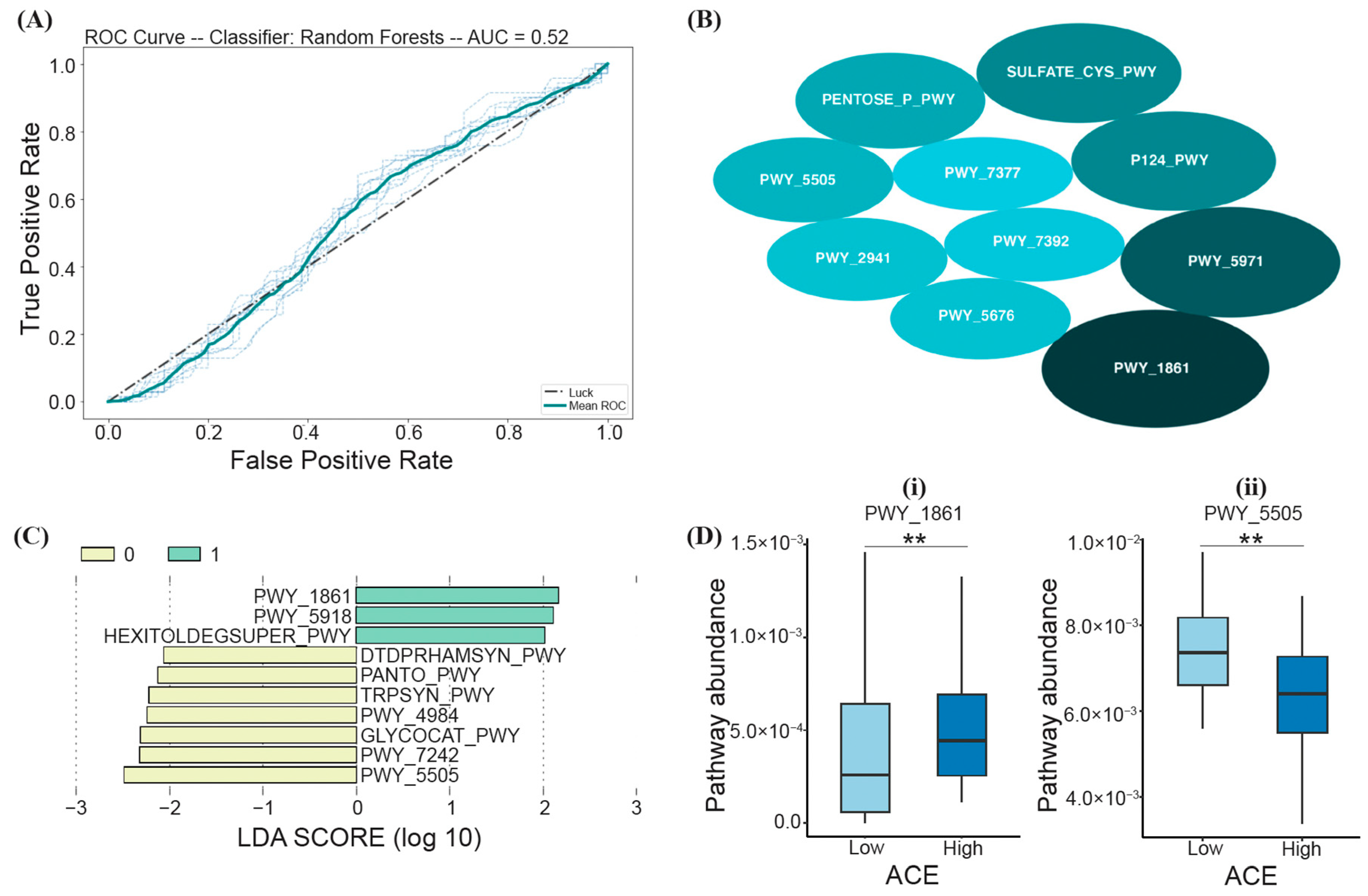
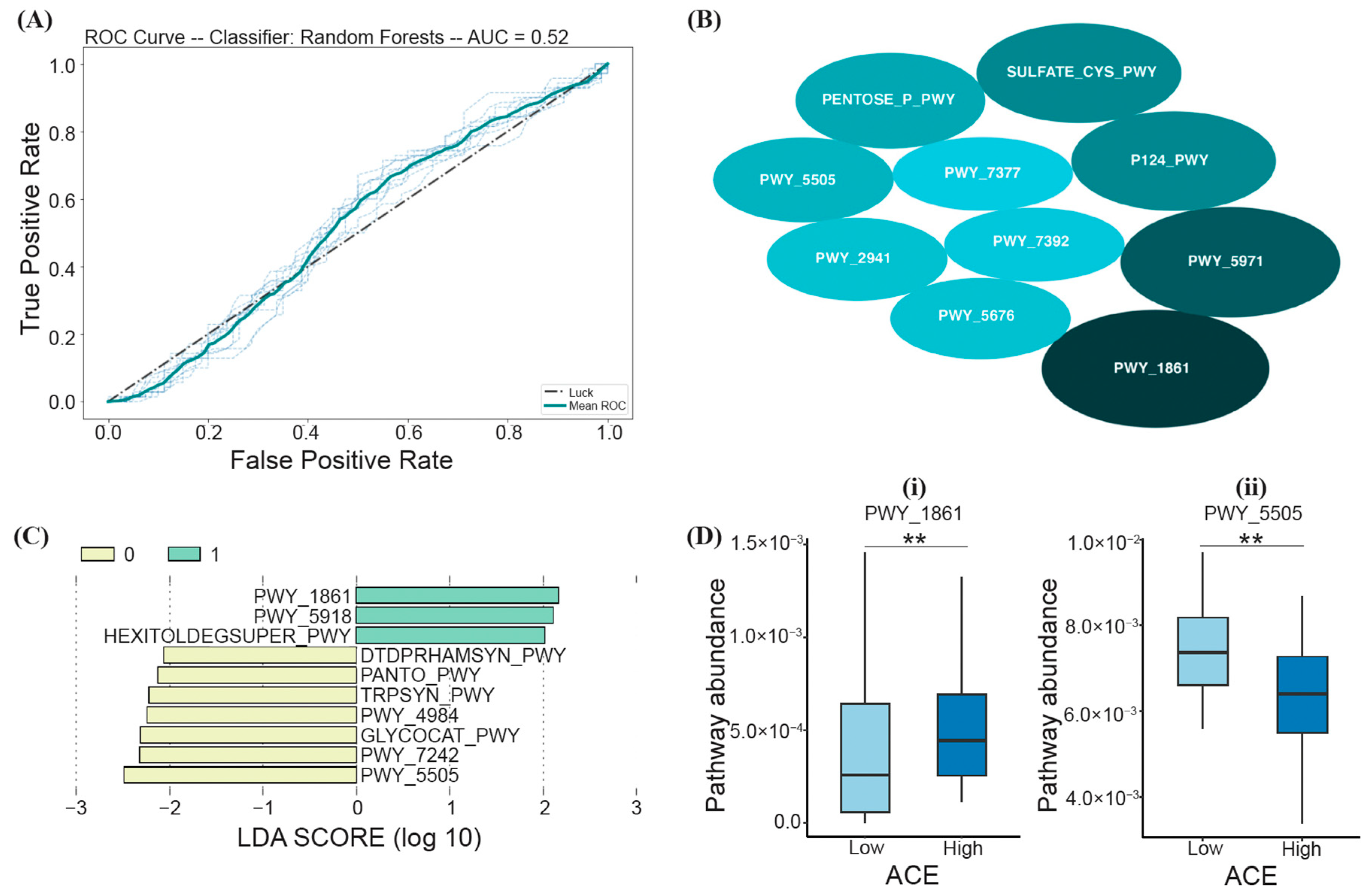
3.4. Functional Differences between ACE Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Felitti, V.J.; Anda, R.F.; Nordenberg, D.; Williamson, D.F.; Spitz, A.M.; Edwards, V.; Koss, M.P.; Marks, J.S. Relationship of childhood abuse and household dysfunction to many of the leading causes of death in adults. The Adverse Childhood Experiences (ACE) Study. Am. J. Prev. Med. 1998, 14, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Cronholm, P.F.; Forke, C.M.; Wade, R.; Bair-Merritt, M.H.; Davis, M.; Harkins-Schwarz, M.; Pachter, L.M.; Fein, J.A. Adverse childhood experiences: Expanding the concept of adversity. Am. J. Prev. Med. 2015, 49, 354–361. [Google Scholar] [CrossRef]
- Wade, R.; Cronholm, P.F.; Fein, J.A.; Forke, C.M.; Davis, M.B.; Harkins-Schwarz, M.; Pachter, L.M.; Bair-Merritt, M.H. Household and community-level adverse childhood experiences and adult health outcomes in a diverse urban population. Child Abus. Negl. 2016, 52, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Anda, R.F.; Butchart, A.; Felitti, V.J.; Brown, D.W. Building a framework for global surveillance of the public health implications of adverse childhood experiences. Am. J. Prev. Med. 2010, 39, 93–98. [Google Scholar] [CrossRef] [PubMed]
- LaNoue, M.; Graeber, D.A.; Helitzer, D.L.; Fawcett, J. Negative affect predicts adults’ ratings of the current, but not childhood, impact of adverse childhood events. Community Ment. Health J. 2013, 49, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Anda, R.F.; Dube, S.R.; Giles, W.H.; Felitti, V.J. The relationship of exposure to childhood sexual abuse to other forms of abuse, neglect, and household dysfunction during childhood. Child Abuse Negl. 2003, 27, 625–639. [Google Scholar] [CrossRef] [PubMed]
- LaNoue, M.; Graeber, D.; de Hernandez, B.U.; Warner, T.D.; Helitzer, D.L. Direct and indirect effects of childhood adversity on adult depression. Community Ment. Health J. 2012, 48, 187–192. [Google Scholar] [CrossRef]
- Dong, M.; Giles, W.H.; Felitti, V.J.; Dube, S.R.; Williams, J.E.; Chapman, D.P.; Anda, R.F. Insights into causal pathways for ischemic heart disease: Adverse childhood experiences study. Circulation 2004, 110, 1761–1766. [Google Scholar] [CrossRef]
- Brown, D.W.; Anda, R.F.; Felitti, V.J.; Edwards, V.J.; Malarcher, A.M.; Croft, J.B.; Giles, W.H. Adverse childhood experiences are associated with the risk of lung cancer: A prospective cohort study. BMC Public Health 2010, 10, 311. [Google Scholar] [CrossRef]
- van der Meer, L.; van Duijn, E.; Wolterbeek, R.; Tibben, A. Adverse childhood experiences of persons at risk for huntington’s disease or BRCA1/2 hereditary breast/ovarian cancer. Clin. Genet. 2012, 81, 18–23. [Google Scholar] [CrossRef]
- Jewkes, R.K.; Dunkle, K.; Nduna, M.; Jama, P.; Puren, A. Associations between childhood adversity and depression, substance abuse and HIV and HSV2 incident infections in rural South African youth. Child Abuse Negl. 2010, 34, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.P.; Whitfield, C.L.; Felitti, V.J.; Dube, S.R.; Edwards, V.J.; Anda, R.F. Adverse childhood experiences and the risk of depressive disorders in adulthood. J. Affect. Disord. 2004, 82, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Dube, S.R.; Fairweather, D.; Pearson, W.S.; Felitti, V.J.; Anda, R.F.; Croft, J.B. Cumulative childhood stress and autoimmune diseases in adults. Psychosom. Med. 2009, 71, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Hillis, S.D.; Anda, R.F.; Felitti, V.J.; Nordenberg, D.; Marchbanks, P.A. Adverse childhood experiences and sexually transmitted diseases in men and women: A retrospective study. Virus Res. 2000, 106, e11. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Anda, R.F. Adverse childhood experiences: Origins of behaviors that sustain the HIV epidemic. AIDS 2009, 23, 2231–2233. [Google Scholar] [CrossRef]
- McLaughlin, K.A.; Greif Green, J.; Gruber, M.J.; Sampson, N.A.; Zaslavsky, A.M.; Kessler, R.C. Childhood adversities and first onset of psychiatric disorders in a national sample of US adolescents. Arch. Gen. Psychiatry 2012, 69, 1151–1160. [Google Scholar] [CrossRef]
- Hatch, V.; Swerbenski, H.; Gray, S.A.O. Family social support buffers the intergenerational association of maternal adverse childhood experiences and preschoolers’ externalizing behavior. Am. J. Orthopsychiatry 2020, 90, 489–501. [Google Scholar] [CrossRef]
- Gellis, L.A.; Lichstein, K.L.; Scarinci, I.C.; Durrence, H.H.; Taylor, D.J.; Bush, A.J.; Riedel, B.W. Socioeconomic status and insomnia. J. Abnorm. Psychol. 2005, 114, 111–118. [Google Scholar] [CrossRef]
- Janssen, I.; Boyce, W.F.; Simpson, K.; Pickett, W. Influence of individual- and area-level measures of socioeconomic status on obesity, unhealthy eating, and physical inactivity in Canadian adolescents. Am. J. Clin. Nutr. 2006, 83, 139–145. [Google Scholar] [CrossRef]
- RodrÍguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef]
- Cowan, C.S.M.; Dinan, T.G.; Cryan, J.F. Annual Research Review: Critical windows—The microbiota-gut-brain axis in neurocognitive development. J. Child Psychol. Psychiatry 2020, 61, 353–371. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Jašarević, E.; Howard, C.D.; Misic, A.M.; Beiting, D.P.; Bale, T.L. Stress during pregnancy alters temporal and spatial dynamics of the maternal and offspring microbiome in a sex-specific manner. Sci. Rep. 2017, 7, 44182. [Google Scholar] [CrossRef] [PubMed]
- Jašarević, E.; Howerton, C.L.; Howard, C.D.; Bale, T.L. Alterations in the vaginal microbiome by maternal stress are associated with metabolic reprogramming of the offspring gut and brain. Endocrinology 2015, 156, 3265–3276. [Google Scholar] [CrossRef] [PubMed]
- Flannery, J.E.; Stagaman, K.; Burns, A.R.; Hickey, R.J.; Roos, L.E.; Giuliano, R.J.; Fisher, P.A.; Sharpton, T.J. Gut feelings begin in childhood: The gut metagenome correlates with early environment, caregiving, and behavior. mBio 2020, 11, e02780-19. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, L.L.; Gawuga, C.E.; Tyrka, A.R.; Lee, J.K.; Anderson, G.M.; Price, L.H. Association between plasma IL-6 response to acute stress and early-life adversity in healthy adults. Neuropsychopharmacology 2010, 35, 2617–2623. [Google Scholar] [CrossRef]
- Danese, A.; Pariante, C.M.; Caspi, A.; Taylor, A.; Poulton, R. Childhood maltreatment predicts adult inflammation in a life-course study. Proc. Natl. Acad. Sci. USA 2007, 104, 1319–1324. [Google Scholar] [CrossRef]
- Fagundes, C.P.; Glaser, R.; Kiecolt-Glaser, J.K. Stressful early life experiences and immune dysregulation across the lifespan. Brain Behav. Immun. 2013, 27, 8–12. [Google Scholar] [CrossRef]
- Ehrlich, K.B.; Ross, K.M.; Chen, E.; Miller, G.E. Testing the biological embedding hypothesis: Is early life adversity associated with a later proinflammatory phenotype? Dev. Psychopathol. 2016, 28 Pt 2, 1273–1283. [Google Scholar] [CrossRef]
- Lovallo, W.R.; Farag, N.H.; Sorocco, K.H.; Cohoon, A.J.; Vincent, A.S. Lifetime adversity leads to blunted stress axis reactivity: Studies from the oklahoma family health patterns project. Biol. Psychiatry 2012, 71, 344–349. [Google Scholar] [CrossRef]
- Mayer, E.A.; Savidge, T.; Shulman, R.J. Brain-gut microbiome interactions and functional bowel disorders. Gastroenterology 2014, 146, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Wei, Y.; Hashimoto, K. Brain–gut–microbiota axis in depression: A historical overview and future directions. Brain Res. Bull. 2022, 182, 44–56. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Nusslock, R.; Miller, G.E. Early-life adversity and physical and emotional health across the lifespan: A neuroimmune network hypothesis. Biol. Psychiatry 2016, 80, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Hantsoo, L.; Jašarević, E.; Criniti, S.; Mcgeehan, B.; Tanes, C.; Sammel, M.D.; Elovitz, M.; Compher, C.; Wu, G.; Epperson, C.N. Childhood adversity impact on gut microbiota and inflammatory response to stress during pregnancy. Brain Behav. Immun. 2019, 75, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Franco, M.R.; Vargas-Luna, M.; Tienda, P.; Delgadillo-Holtfort, I.; Balleza-Ordaz, M.; Flores-Hernandez, C. Effects of occupational stress on the gastrointestinal tract. World J. Gastrointest. Pathophysiol. 2013, 4, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Lavoie-Tremblay, M.; Sanzone, L.; Aubé, T.; Paquet, M. Sources of stress and coping strategies among undergraduate nursing students across all years. Can. J. Nurs. Res. 2021, 54, 261–271. [Google Scholar] [CrossRef]
- Jimenez, C.; Navia-Osorio, M.P.; Diaz, C.V. Stress and health in novice and experienced nursing students. J. Adv. Nurs. 2010, 66, 442–455. [Google Scholar] [CrossRef]
- Rella, S.; Winwood, P.C.; Lushingtoon, K. When does nursing burnout begin? An investigation of the fatigue experience of australian nursing students. J. Nurs. Manag. 2009, 17, 886–897. [Google Scholar] [CrossRef]
- McKee-Lopez, G.; Robbins, L.; Provencio-Vasquez, E.; Olvera, H. The relationship of childhood adversity on burnout and depression among BSN students. J. Prof. Nurs. 2019, 35, 112–119. [Google Scholar] [CrossRef]
- Limbana, T.; Khan, F.; Eskander, N. Gut microbiome and depression: How microbes affect the way we think. Curēus 2020, 12, e9966. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, G.; Siopi, E.; Guenin-Macé, L.; Pascal, M.; Laval, T.; Rifflet, A.; Boneca, I.G.; Demangel, C.; Colsch, B.; Pruvost, A.; et al. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun. 2020, 11, 6363. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.G.; Goldenthal, A.R.; Uhlemann, A.; Mann, J.J.; Miller, J.M.; Sublette, M.E. Systematic review of gut microbiota and major depression. Front. Psychiatry 2019, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Petrak, F.; Herpertz, S.; Hirsch, J.; Röhrig, B.; Donati-Hirsch, I.; Juckel, G.; Meier, J.J.; Gatermann, S. Gut microbiota differs in composition between adults with type 1 diabetes with or without depression and healthy control participants: A case-control study. BMC Microbiol. 2022, 22, 169. [Google Scholar] [CrossRef]
- Gibbons, C. Stress, coping and burn-out in nursing students. Int. J. Nurs. Stud. 2010, 47, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.; Kim, J. Factors affecting academic burnout of nursing students according to clinical practice experience. BMC Med. Educ. 2022, 22, 346. [Google Scholar] [CrossRef] [PubMed]
- Njim, T.; Mbanga, C.; Mouemba, D.; Makebe, H.; Toukam, L.; Kika, B.; Mulango, I. Determinants of depression among nursing students in cameroon: A cross-sectional analysis. BMC Nurs. 2020, 19, 26. [Google Scholar] [CrossRef] [PubMed]
- Olvera, H.A.; Provencio-Vasquez, E.; Slavich, G.M.; Laurent, J.G.C.; Browning, M.; McKee-Lopez, G.; Robbins, L.; Spengler, J.D. Stress and health in nursing students: The nurse engagement and wellness study. Nurs. Res. 2019, 68, 453–463. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Adverse Childhood Experiences (ACEs). 2021. Available online: https://www.cdc.gov/violenceprevention/aces/index.html (accessed on 1 September 2023).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2020. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 1 September 2023).
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D. Ggplot2: Create Elegant Data Visualizations Using the Grammar of Graphics. 2020. Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 1 September 2023).
- Deng, C.; Daley, T.; Smith, A. preseqR: Predicting Species Accumulation Curves. 2018. Available online: https://CRAN.R-project.org/package=preseqR (accessed on 1 September 2023).
- Wickham, H. Reshape2: Flexibly Reshape Data: A Reboot of the Reshape Package. 2020. Available online: https://cran.r-project.org/web/packages/reshape2/index.html (accessed on 1 September 2023).
- Cutler, D.R.; Edwards, T.C.J.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Cao, L.J.; Chua, K.S.; Chong, W.K.; Lee, H.P.; Gu, Q.M. A comparison of PCA, KPCA and ICA for dimensionality reduction in support vector machine. Neurocomputing 2003, 55, 321–336. [Google Scholar] [CrossRef]
- Dietterich, T.G. Approximate statistical tests for comparing supervised classification learning algorithms. Neural Comput. 1998, 10, 1895–1923. [Google Scholar] [CrossRef]
- Louppe, G.; Wehenkel, L.; Sutera, A.; Geurts, P. Understanding variable importances in forests of randomized trees. Neural Inf. Process. Syst. 2013, 2013, 431–439. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Batista, G.E.; Monard, M.C. A study of k-nearest neighbour as an imputation method. Soft Comput. Syst. Des. Manag. Appl. 2002, 48, 251–260. [Google Scholar]
- Iljazovic, A.; Roy, U.; Gálvez, E.J.C.; Lesker, T.R.; Zhao, B.; Gronow, A.; Amend, L.; Will, S.E.; Hofmann, J.D.; Pils, M.C.; et al. Perturbation of the gut microbiome by Prevotella spp. enhances host susceptibility to mucosal inflammation. Mucosal Immunol. 2021, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Kummen, M.; Mayerhofer, C.C.K.; Vestad, B.; Broch, K.; Awoyemi, A.; Storm-Larsen, C.; Ueland, T.; Yndestad, A.; Hov, J.R.; Trøseid, M. Gut microbiota signature in heart failure defined from profiling of 2 independent cohorts. J. Am. Coll. Cardiol. 2018, 71, 1184–1186. [Google Scholar] [CrossRef]
- Kim, M.; Kim, H. The roles of glutamine in the intestine and its implication in intestinal diseases. Int. J. Mol. Sci. 2017, 18, 1051. [Google Scholar] [CrossRef]
- Graham, C.; Mullen, A.; Whelan, K. Obesity and the gastrointestinal microbiota: A review of associations and mechanisms. Nutr. Rev. 2015, 73, 376–385. [Google Scholar] [CrossRef]
- Wu, K.; Sun, W.; Li, N.; Chen, Y.; Wei, Y.; Chen, D. Small intestinal bacterial overgrowth is associated with diarrhea-predominant irritable bowel syndrome by increasing mainly Prevotella abundance. Scand. J. Gastroenterol. 2019, 54, 1419–1425. [Google Scholar] [CrossRef]
- Mönnikes, H.; Schmidt, B.G.; Taché, Y. Psychological stress-induced accelerated colonic transit in rats involves hypothalamic corticotropin-releasing factor. Gastroenterology 1993, 104, 716. [Google Scholar] [CrossRef]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. The Gut–Brain axis and the microbiome: Mechanisms and clinical implications. Clin. Gastroenterol. Hepatol. 2019, 17, 322–332. [Google Scholar] [CrossRef]
- Toh, T.S.; Wei Jie Lee, J.; Toh, K.Y.; Ho, J.P.; Fung Yen Lim, J.; Tan, A.H.; Chong, C.W. IDDF2021-ABS-0165 Psychological well-being and sleep quality among healthy stool donors in Singapore: A cross-sectional study. Gut 2021, 70, A50–A51. [Google Scholar]
- Qu, Y.; Yang, C.; Ren, Q.; Ma, M.; Dong, C.; Hashimoto, K. Comparison of (R)-ketamine and lanicemine on depression-like phenotype and abnormal composition of gut microbiota in a social defeat stress model. Sci. Rep. 2017, 7, 15725. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Qu, Y.; Fujita, Y.; Ren, Q.; Ma, M.; Dong, C.; Hashimoto, K. Possible role of the gut microbiota-brain axis in the antidepressant effects of (R)-ketamine in a social defeat stress model. Transl. Psychiatry 2017, 7, 1294. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, K.J.; Moran-Santa Maria, M.M.; Twal, W.O.; Shaftman, S.; Desantis, S.M.; Mcrae-Clark, A.L.; Brady, K.T. Association of elevated cytokines with childhood adversity in a sample of healthy adults. J. Psychiatr. Res. 2013, 47, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Rea, K.; Dinan, T.G.; Cryan, J.F. The microbiome: A key regulator of stress and neuroinflammation. Neurobiol. Stress 2016, 4, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Moussaoui, N.; Larauche, M.; Biraud, M.; Molet, J.; Million, M.; Mayer, E.; Taché, Y. Limited nesting stress alters maternal behavior and in vivo intestinal permeability in male wistar pup rats. PLoS ONE 2016, 11, e0155037. [Google Scholar] [CrossRef]
- De Palma, G.; Blennerhassett, P.; Lu, J.; Deng, Y.; Park, A.J.; Green, W.; Denou, E.; Silva, M.A.; Santacruz, A.; Sanz, Y.; et al. Microbiota and host determinants of behavioural phenotype in maternally separated mice. Nat. Commun. 2015, 6, 7735. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; Marchesi, J.R.; Scully, P.; Codling, C.; Ceolho, A.-M.; Quigley, E.M.M.; Cryan, J.F.; Dinan, T.G. Early life stress alters behavior, immunity, and microbiota in rats: Implications for irritable bowel syndrome and psychiatric illnesses. Biol. Psychiatry 2009, 65, 263–267. [Google Scholar] [CrossRef]
- Agargun, M.Y.; Kara, H.; Solmaz, M. Sleep disturbances and suicidal behavior in patients with major depression. J. Clin. Psychiatry 1997, 58, 249–251. [Google Scholar]
- Paterson, L.M.; Nutt, D.J.; Wilson, S.J. NAPSAQ-1: National patient sleep assessment questionnaire in depression. Int. J. Psychiatry Clin. Pract. 2009, 13, 48–58. [Google Scholar] [CrossRef]
- Khanijow, V.; Prakash, P.; Emsellem, H.A.; Borum, M.L.; Doman, D.B. Sleep dysfunction and gastrointestinal diseases. Gastroenterol. Hepatol. 2015, 11, 817–825. [Google Scholar]
- Graham, C.D.; Kyle, S.D. A preliminary investigation of sleep quality in functional neurological disorders: Poor sleep appears common, and is associated with functional impairment. J. Neurol. Sci. 2017, 378, 63–166. [Google Scholar] [CrossRef] [PubMed]
- Hyun, M.K.; Baek, Y.; Lee, S. Association between digestive symptoms and sleep disturbance: A cross-sectional community-based study. BMC Gastroenterol. 2019, 19, 34. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota diurnal rhythmicity programs host transcriptome oscillations. Cell 2016, 167, 1495–1510.e12. [Google Scholar] [CrossRef]
- Davies, S.; Ang, J.; Revell, V.; Holmes, B.; Mann, A.; Robertson, F.; Cui, N.; Middleton, B.; Ackermann, K.; Kayser, M.; et al. Effect of sleep deprivation on the human metabolome. Proc. Natl. Acad. Sci. USA 2014, 111, 10761–10766. [Google Scholar] [CrossRef]
- Baldwin, J.R.; Danese, A. Pathways from childhood maltreatment to cardiometabolic disease: A research review. Adopt. Foster. 2019, 43, 329–339. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dimension | n | Mean (%) | SD | |
|---|---|---|---|---|
| Age | - | 150 | 26 | 6.5 |
| BMI | - | 150 | 26.3 | 5.6 |
| Gender | Male | 26 | (17) | - |
| Female | 124 | (83) | - | |
| Ethnicity | Hispanic | 130 | (87) | - |
| Non-Hispanic | 20 | (13) | - | |
| Race | American Indian | 5 | (3) | - |
| Asian | 6 | (4) | - | |
| African American | 7 | (5) | - | |
| Native Hawaiian | 0 | (0) | - | |
| White | 132 | (88) | - | |
| ACE | Low (<3) | 80 | (53) | - |
| High (≥3) | 70 | (47) | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazemian, N.; Zhou, T.; Chalasani, N.; Narayan, A.; Cedeño Laurent, J.G.; Olvera Alvarez, H.A.; Pakpour, S. Long-Term Impact of Childhood Adversity on the Gut Microbiome of Nursing Students. Int. J. Environ. Res. Public Health 2024, 21, 68. https://doi.org/10.3390/ijerph21010068
Kazemian N, Zhou T, Chalasani N, Narayan A, Cedeño Laurent JG, Olvera Alvarez HA, Pakpour S. Long-Term Impact of Childhood Adversity on the Gut Microbiome of Nursing Students. International Journal of Environmental Research and Public Health. 2024; 21(1):68. https://doi.org/10.3390/ijerph21010068
Chicago/Turabian StyleKazemian, Negin, Tony Zhou, Naveen Chalasani, Apurva Narayan, Jose Guillermo Cedeño Laurent, Hector A. Olvera Alvarez, and Sepideh Pakpour. 2024. "Long-Term Impact of Childhood Adversity on the Gut Microbiome of Nursing Students" International Journal of Environmental Research and Public Health 21, no. 1: 68. https://doi.org/10.3390/ijerph21010068
APA StyleKazemian, N., Zhou, T., Chalasani, N., Narayan, A., Cedeño Laurent, J. G., Olvera Alvarez, H. A., & Pakpour, S. (2024). Long-Term Impact of Childhood Adversity on the Gut Microbiome of Nursing Students. International Journal of Environmental Research and Public Health, 21(1), 68. https://doi.org/10.3390/ijerph21010068