
Climate Stressors and Physiological Dysregulations: Mechanistic Connections to Pathologies



Abstract
:1. Introduction
2. Climate Change Impact on the Exogenous and Endogenous Environment
2.1. Heat and OS
2.2. Commonality of OS with Pathologies and Altered Immunity
2.3. OS and Neurological Disorders
2.4. OS and Cardiovascular Disorders
2.5. Genetic Variations and Susceptibility
2.6. Folate and Methionine Cycles
2.7. Genetics, Diversity, and Disparities
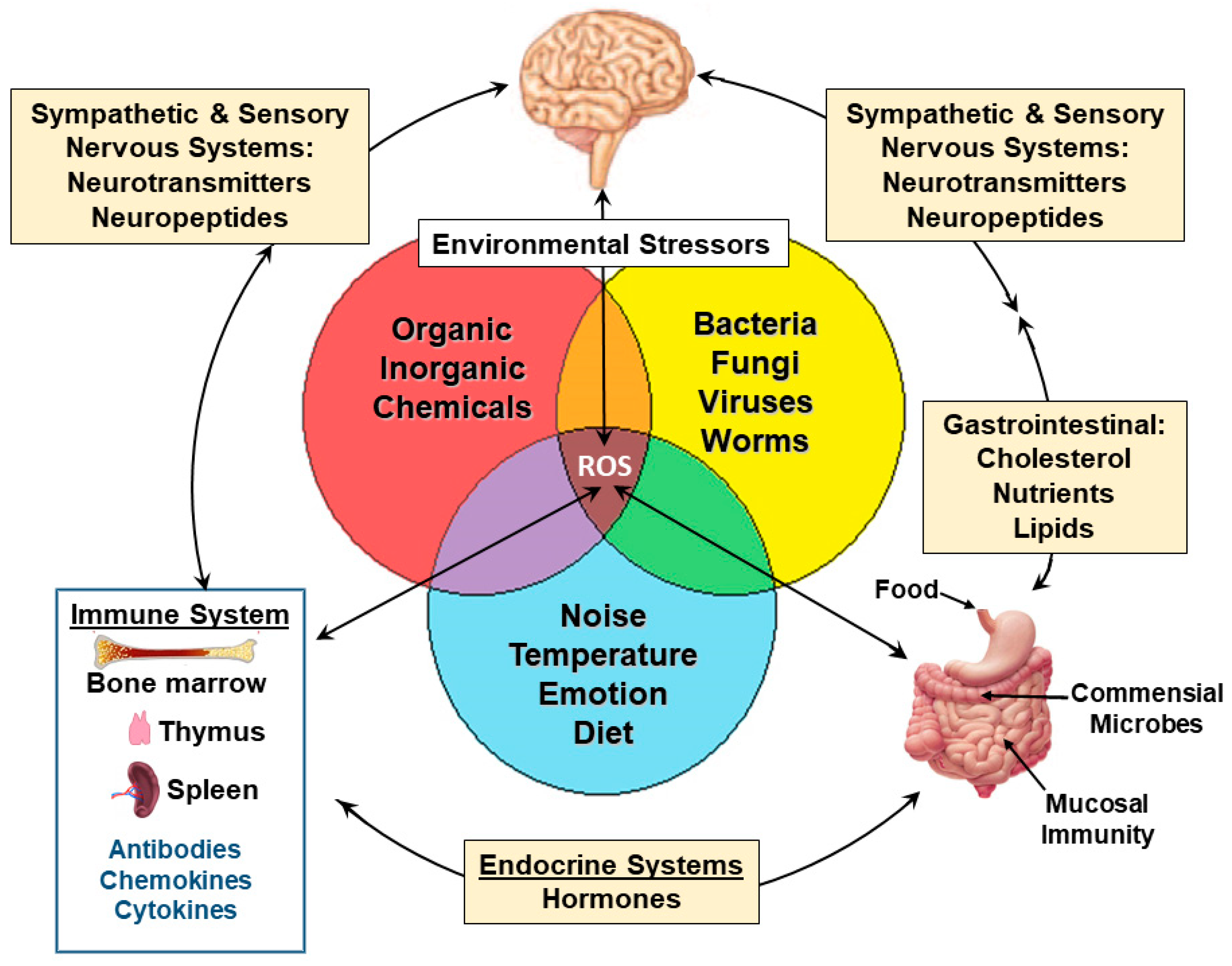
2.8. Mechanistic Connections of Genes and Proteins to Pathologies
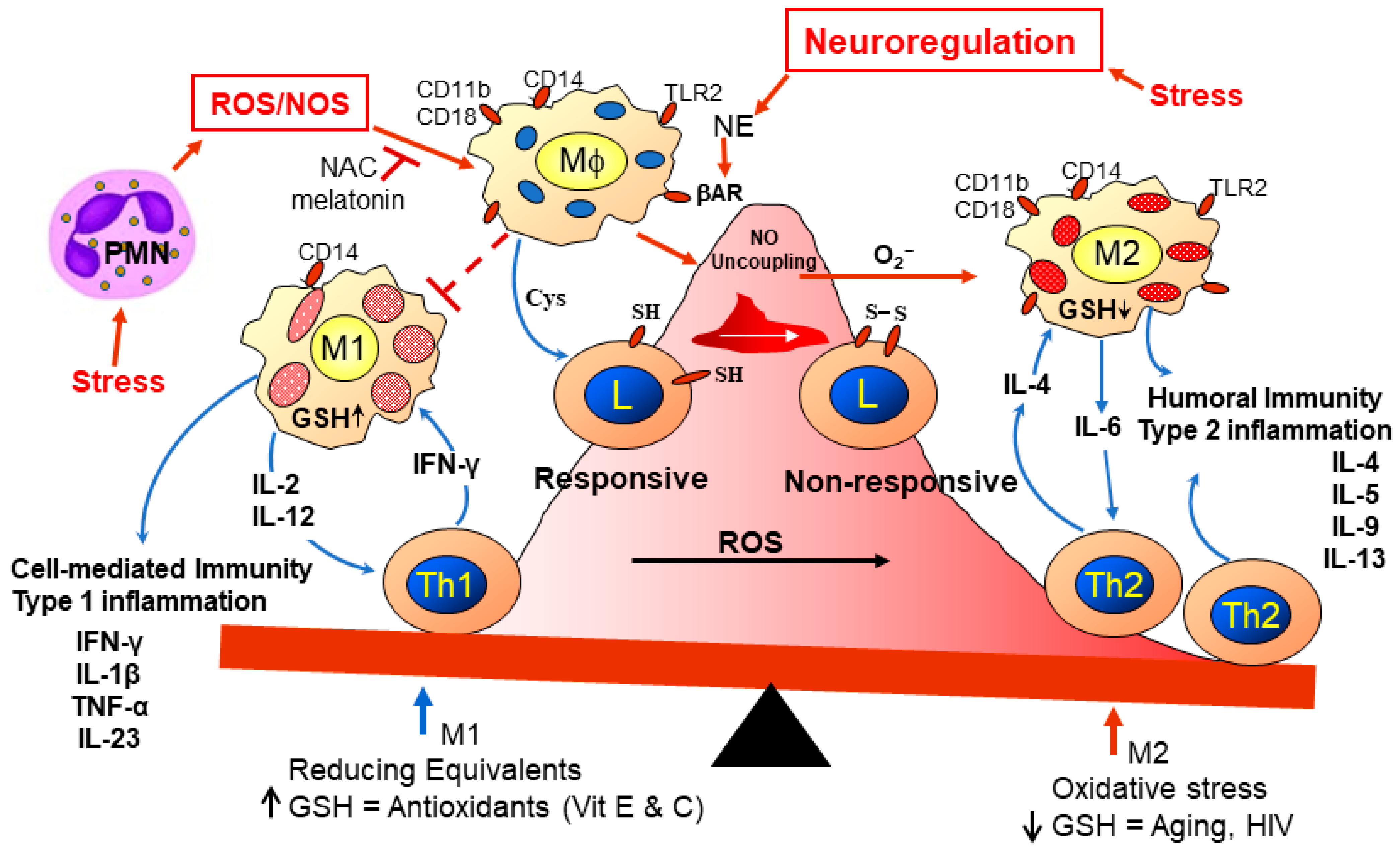
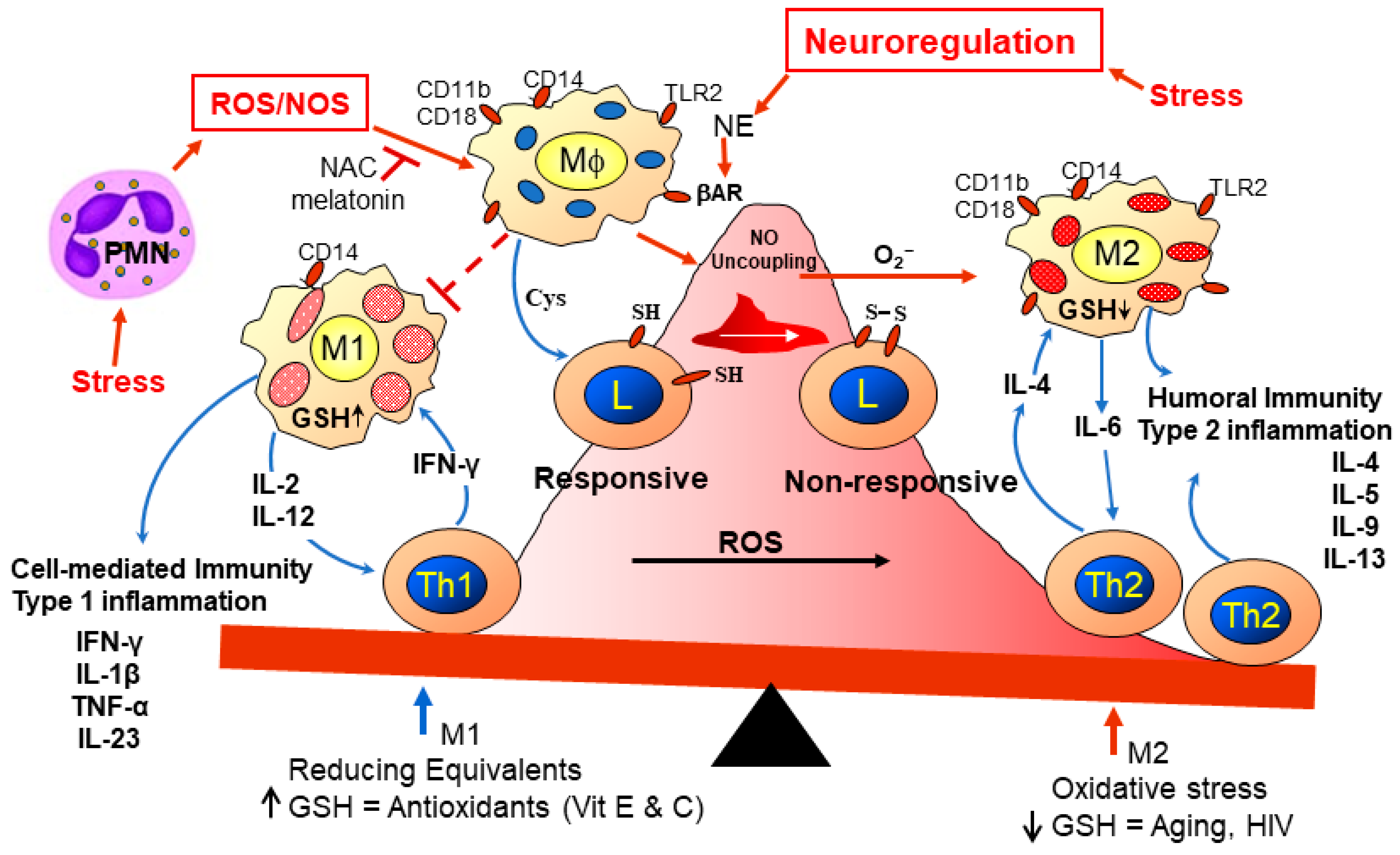
2.9. Metabolic Syndrome, Lipid Dysregulation, and Immune Activity

{kind=link}
{kind=link}
{kind=link}
| Factor | Connection with Metabolic Syndrome | Connection with Dyslipidemia | Connection with OS | Reference |
|---|---|---|---|---|
| Environmental stressors | Exacerbates Metabolic Syndrome by increasing stress hormones and inflammation. | May lead to dyslipidemia through dietary and lifestyle factors. | Can trigger oxidative stress through the release of free radicals in response to stress. | Masenga et al. [143] Fahed et al. [144] |
| Metabolic Syndrome | - | Dyslipidemia is a common component of Metabolic Syndrome. | Oxidative stress often accompanies Metabolic Syndrome due to chronic inflammation. | Huang [145] |
| Dyslipidemia | Dyslipidemia is a hallmark feature of Metabolic Syndrome. | - | Dyslipidemia can contribute to oxidative stress through the production of reactive oxygen species (ROS). | Tangvarasittichai [131] |
| Oxidative Stress | Oxidative stress is closely linked to Metabolic Syndrome, contributing to its progression. | Can be exacerbated by dyslipidemia due to increased ROS production. | - | Roberts and Sindhu [130] |
| Pathology | Association with Metabolic Syndrome | Reference |
|---|---|---|
| Dyslipidemia | Dyslipidemia is a common component of metabolic syndrome, characterized by elevated triglycerides and low HDL cholesterol. | Pappan and Rehman [146] |
| Oxidative Stress | Metabolic syndrome is often accompanied by oxidative stress, marked by increased levels of reactive oxygen species (ROS). | Monserrat-Mesquida et al. [147] |
| Inflammation | Chronic inflammation is a hallmark of Metabolic Syndrome and contributes to its progression. | Pahwa et al. [148] |
| Insulin Resistance | Metabolic Syndrome is closely linked to insulin resistance, which leads to elevated blood sugar levels. | Fahed et al. [144] |
| Hypertension | Hypertension often co-occurs with metabolic syndrome and contributes to cardiovascular risks. | Stanciu et al. [149] |
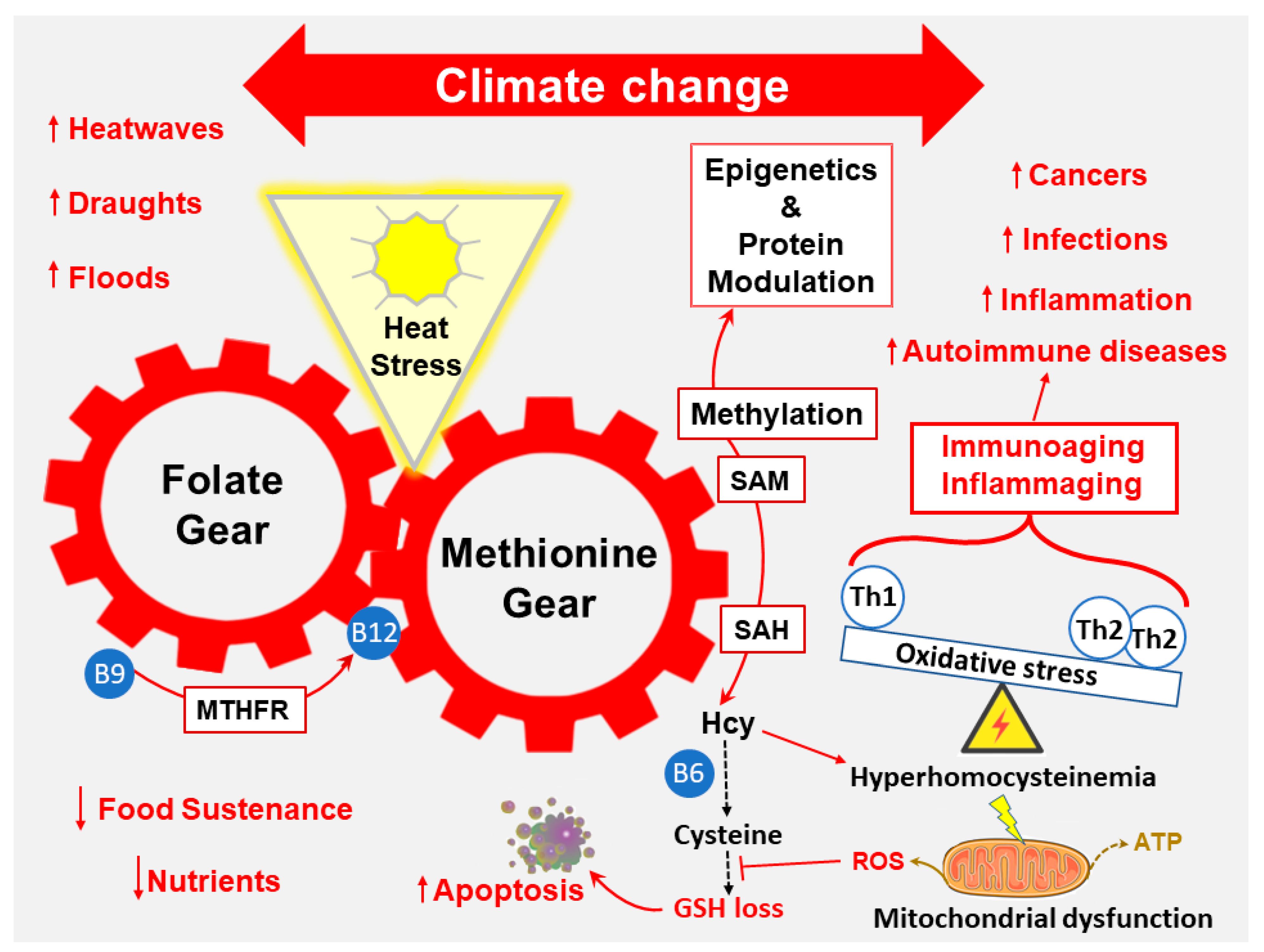
2.10. Environmental Stressors’ Influence on Metabolic Syndrome: A Complex Interplay
3. Conclusions
4. Future Directions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prüss-Ustün, A.; Wolf, J.; Corvalán, C.; Neville, T.; Bos, R.; Neira, M. Diseases due to unhealthy environments: An updated estimate of the global burden of disease attributable to environmental determinants of health. J. Public. Health 2017, 39, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Reuben, A.; Manczak, E.M.; Cabrera, L.Y.; Alegria, M.; Bucher, M.L.; Freeman, E.C.; Miller, G.W.; Solomon, G.M.; Perry, M.J. The Interplay of Environmental Exposures and Mental Health: Setting an Agenda. Environ. Health Perspect. 2022, 130, 25001. [Google Scholar] [CrossRef] [PubMed]
- Cianconi, P.; Betrò, S.; Janiri, L. The Impact of Climate Change on Mental Health: A Systematic Descriptive Review. Front. Psychiatry 2020, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Felix, E.; Rubens, S.; Hambrick, E. The Relationship Between Physical and Mental Health Outcomes in Children Exposed to Disasters. Curr. Psychiatry Rep. 2020, 22, 33. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.; Colley, K.; Currie, M.; Eastwood, A.; Li, K.H.; Avery, L.M.; Beevers, L.C.; Braithwaite, I.; Dallimer, M.; Davies, Z.G.; et al. The Contribution of Environmental Science to Mental Health Research: A Scoping Review. Int. J. Environ. Res. Public Health 2023, 20, 5278. [Google Scholar] [CrossRef] [PubMed]
- Weilnhammer, V.; Schmid, J.; Mittermeier, I.; Schreiber, F.; Jiang, L.; Pastuhovic, V.; Herr, C.; Heinze, S. Extreme weather events in europe and their health consequences—A systematic review. Int. J. Hyg. Environ. Health. 2021, 233, 113688. [Google Scholar] [CrossRef] [PubMed]
- Caminade, C.; McIntyre, K.M.; Jones, A.E. Impact of recent and future climate change on vector-borne diseases. Ann. N. Y. Acad. Sci. 2019, 1436, 157–173. [Google Scholar] [CrossRef]
- Semenza, J.C.; Rocklöv, J.; Ebi, K.L. Climate Change and Cascading Risks from Infectious Disease. Infect Dis Ther. 2022, 11, 1371–1390. [Google Scholar] [CrossRef]
- Anderson, H.; Brown, C.; Cameron, L.L.; Christenson, M.; Conlon, K.C.; Dorevitch, S.; Dumas, J.; Eidson, M.; Ferguson, A.; Grossman, E.; et al. BRACE Midwest and Southeast Community of Practice. Climate and Health Intervention Assessment: Evidence on Public Health Interventions to Prevent the Negative Health Effects of Climate Change. Climate and Health Technical Report Series. Climate and Health Program, CDC. 2017. Available online: https://www.cdc.gov/climateandhealth/docs/ClimateAndHealthInterventionAssessment_508.pdf (accessed on 28 August 2023).
- Ranganayaki, S.; Jamshidi, N.; Aiyaz, M.; Rashmi, S.K.; Gayathri, N.; Harsha, P.K.; Padmanabhan, B.; Srinivas Bharath, M.M. Inhibition of mitochondrial complex II in neuronal cells triggers unique pathways culminating in autophagy with implications for neurodegeneration. Sci. Rep. 2021, 11, 1483. [Google Scholar] [CrossRef]
- Kenney, W.L.; Craighead, D.H.; Alexander, L.M. Heat waves, aging, and human cardiovascular health. Med. Sci. Sports Exerc. 2014, 46, 1891–1899. [Google Scholar] [CrossRef]
- Marchand, M.; Gin, K. The Cardiovascular System in Heat Stroke. CJC Open 2021, 4, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.; White, P.C.L.; Bell, A.; Coventry, P.A. Effect of Extreme Weather Events on Mental Health: A Narrative Synthesis and Meta-Analysis for the UK. Int. J. Environ. Res. Public. Health. 2020, 17, 8581. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, R.; Schymanski, E.L.; Barabási, A.L.; Miller, G.W. The exposome and health: Where chemistry meets biology. Science 2020, 367, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Halverson, S.; Ezinwa, N. Mosquito-Borne Diseases. Prim. Care 2018, 45, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Prüss-Ustün, A.; Wolf, J.; Corvalán, C.; Bos, R.; Neira, M. Preventing Disease through Healthy Environments: A Global Assessment of the Burden of Disease from Environmental Risks; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Bou Zerdan, M.; Moussa, S.; Atoui, A.; Assi, H.I. Mechanisms of Immunotoxicity: Stressors and Evaluators. Int. J. Mol. Sci. 2021, 22, 8242. [Google Scholar] [CrossRef] [PubMed]
- Marazziti, D.; Cianconi, P.; Mucci, F.; Foresi, L.; Chiarantini, I.; Della Vecchia, A. Climate change, environment pollution, COVID-19 pandemic and mental health. Sci. Total Environ. 2021, 773, 145182. [Google Scholar] [CrossRef] [PubMed]
- Emeny, R.T.; Carpenter, D.O.; Lawrence, D.A. Health disparities: Intracellular consequences of social determinants of health. Toxicol. Appl. Pharmacol. 2021, 416, 115444. [Google Scholar] [CrossRef] [PubMed]
- Downey, L.; Van Willigen, M. Environmental stressors: The mental health impacts of living near industrial activity. J. Health Soc. Behav. 2005, 46, 289–305. [Google Scholar] [CrossRef]
- Karimi, P.; Kamali, E.; Mousavi, S.M.; Karahmadi, M. Environmental factors influencing the risk of autism. J. Res. Med. Sci. 2017, 22, 27. [Google Scholar] [CrossRef]
- Deng, S.Z.; Jalaludin, B.B.; Antó, J.M.; Hess, J.J.; Huang, C.R. Climate change, air pollution, and allergic respiratory diseases: A call to action for health professionals. Chin. Med. J. 2020, 133, 1552–1560. [Google Scholar] [CrossRef]
- Choi, Y.; Kwon, H.K.; Park, S. Polygenic Variants Linked to Oxidative Stress and the Antioxidant System Are Associated with Type 2 Diabetes Risk and Interact with Lifestyle Factors. Antioxidants 2023, 12, 1280. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J.A.; Teichmann, S.A. Predicting pathogenic protein variants. Science 2023, 381, 1284–1285. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.A.; Khatami, M.; Baglole, C.J.; Sun, J.; Harris, S.A.; Moon, E.Y.; Al-Mulla, F.; Al-Temaimi, R.; Brown, D.G.; Colacci, A.; et al. Environmental immune disruptors, inflammation and cancer risk. Carcinogenesis 2015, 36 (Suppl. S1), S232–S253. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ren, Z.; Zhang, J.; Chuang, C.C.; Kandaswamy, E.; Zhou, T.; Zuo, L. Role of ROS and Nutritional Antioxidants in Human Diseases. Front. Physiol. 2018, 9, 477. [Google Scholar] [CrossRef]
- Kale, A.; Sharma, A.; Stolzing, A.; Desprez, P.Y.; Campisi, J. Role of immune cells in the removal of deleterious senescent cells. Immun. Ageing. 2020, 17, 16. [Google Scholar] [CrossRef]
- Tinelli, C.; Di Pino, A.; Ficulle, E.; Marcelli, S.; Feligioni, M. Hyperhomocysteinemia as a Risk Factor and Potential Nutraceutical Target for Certain Pathologies. Front. Nutr. 2019, 6, 49. [Google Scholar] [CrossRef]
- Raghubeer, S.; Matsha, T.E. Methylenetetrahydrofolate (MTHFR), the One-Carbon Cycle, and Cardiovascular Risks. Nutrients 2021, 13, 4562. [Google Scholar] [CrossRef]
- Peng, J.; Wu, Z. MTHFR act as a potential cancer biomarker in immune checkpoints blockades, heterogeneity, tumor microenvironment and immune infiltration. Discov. Oncol. 2023, 14, 112. [Google Scholar] [CrossRef]
- Zhou, H.; Ge, T.; Li, H.; Fang, T.; Li, H.; Shi, Y.; Zhang, R.; Dong, X. A Multi-Medium Analysis of Human Health Risk of Toxic Elements in Rice-Crayfish System: A Case Study from Middle Reach of Yangtze River, China. Food 2022, 11, 1160. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.L.; Zanobetti, A.; Dominici, F. Who is more affected by ozone pollution? A systematic review and meta-analysis. Am. J. Epidemiol. 2014, 180, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Frumkin, H.; Holloway, T.; Vimont, D.J.; Haines, A. Climate change: Challenges and opportunities for global health. JAMA 2014, 312, 1565–1580. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Landrigan, P.J. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Haines, A.; McMichael, A.J.; Smith, K.R.; Roberts, I.; Woodcock, J.; Markandya, A.; Armstrong, B.G.; Campbell-Lendrum, D.; Dangour, A.D.; Davies, M.; et al. 2009. Public health benefits of strategies to reduce greenhouse-gas emissions: Overview and implications for policy makers. Lancet 2009, 374, 2104–2114. [Google Scholar] [CrossRef]
- Watts, N.; Adger, W.N.; Ayeb-Karlsson, S.; Bai, Y.; Byass, P.; Campbell-Lendrum, D.; Colbourn, T.; Cox, P.; Davies, M.; Depledge, M.; et al. The Lancet Countdown: Tracking progress on health and climate change. Lancet 2017, 389, 1151–1164. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Campbell-Lendrum, D.; Holloway, T.; Foley, J.A. Impact of regional climate change on human health. Nature 2005, 438, 310–317. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, G.; Pawankar, R.; Vitale, C.; Lanza, M.; Molino, A.; Stanziola, A.; Sanduzzi, A.; Vatrella, A.; D’Amato, M. Climate Change and Air Pollution: Effects on Respiratory Allergy. Allergy Asthma Immunol. Res. 2016, 8, 391–395. [Google Scholar] [CrossRef]
- Hansen, A.; Bi, P.; Nitschke, M.; Ryan, P.; Pisaniello, D.; Tucker, G. The effect of heat waves on mental health in a temperate Australian city. Environ. Health Perspect. 2008, 116, 1369–1375. [Google Scholar] [CrossRef]
- Pacheco, S.E. Catastrophic effects of climate change on children’s health start before birth. J. Clin. Invest. 2020, 130, 562–564. [Google Scholar] [CrossRef]
- Singh, A.B.; Kumar, P. Climate change and allergic diseases: An overview. Front. Allergy 2022, 3, 964987. [Google Scholar] [CrossRef] [PubMed]
- Ghaisas, S.; Maher, J.; Kanthasamy, A. Gut microbiome in health and disease: Linking the microbiome-gut-brain axis and environmental factors in the pathogenesis of systemic and neurodegenerative diseases. Pharmacol. Ther. 2016, 158, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sharma, P.; Pal, N.; Kumawat, M.; Shubham, S.; Sarma, D.K.; Tiwari, R.R.; Kumar, M.; Nagpal, R. Impact of Environmental Pollutants on Gut Microbiome and Mental Health via the Gut-Brain Axis. Microorganisms 2022, 10, 1457. [Google Scholar] [CrossRef] [PubMed]
- Gronlund, C.J. Racial and socioeconomic disparities in heat-related health effects and their mechanisms: A review. Curr. Epidemiol. Rep. 2014, 1, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Ahima, R.S. Global warming threatens human thermoregulation and survival. J. Clin. Invest. 2020, 130, 559–561. [Google Scholar] [CrossRef] [PubMed]
- Crandall, C.G.; Wilson, T.E. Human cardiovascular responses to passive heat stress. Compr. Physiol. 2015, 5, 17–43. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.B. Physiology of sweat gland function: The roles of sweating and sweat composition in human health. Temperature 2019, 6, 211–259. [Google Scholar] [CrossRef]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell. 2012, 48, 158–167. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Paray, B.A.; Albeshr, M.F.; Jan, A.T.; Rather, I.A. Leaky Gut and Autoimmunity: An Intricate Balance in Individuals Health and the Diseased State. Int. J. Mol. Sci. 2020, 21, 9770. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Lawrence, D.A.; McCabe, M.J., Jr. Immunomodulation by metals. Int. Immunopharmacol. 2002, 2, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Mishra, K.P. Lead exposure and its impact on immune system: A review. Toxicol. In Vitro 2009, 23, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Filipov, N.M.; Lawrence, D.A. Sympathetic nervous system plays a major role in acute cold/restraint stress inhibition of host resistance to Listeria monocytogenes. J. Neuroimmunol. 2002, 125, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Childs, C.E.; Calder, P.C.; Miles, E.A. Diet and Immune Function. Nutrients 2019, 11, 1933. [Google Scholar] [CrossRef] [PubMed]
- Glencross, D.A.; Ho, T.R.; Camiña, N.; Hawrylowicz, C.M.; Pfeffer, P.E. Air pollution and its effects on the immune system. Free Radic. Biol. Med. 2020, 151, 56–68. [Google Scholar] [CrossRef]
- Pan, S.; Wang, J.; Wu, A.; Guo, Z.; Wang, Z.; Zheng, L.; Dai, Y.; Zhu, L.; Nie, J.; Hei, T.K.; et al. Radiation Exposure-Induced Changes in the Immune Cells and Immune Factors of Mice with or without Primary Lung Tumor. Dose Response 2020, 18, 1559325820926744. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Rashamol, V.P.; Bagath, M.; Sejian, V.; Dunshea, F.R. Impacts of heat stress on immune responses and oxidative stress in farm animals and nutritional strategies for amelioration. Int. J. Biometeorol. 2021, 65, 1231–1244. [Google Scholar] [CrossRef]
- Schraml, E.; Quan, P.; Stelzer, I.; Fuchs, R.; Skalicky, M.; Viidik, A.; Schauenstein, K. Norepinephrine treatment and aging lead to systemic and intracellular oxidative stress in rats. Exp. Gerontol. 2007, 42, 1072–1078. [Google Scholar] [CrossRef]
- Fields, C.T.; Chassaing, B.; de Vries, G.J. Gut Barrier Dysfunction and Type 2 Immunity: Implications for Compulsive Behavior. Med. Hypotheses 2022, 161, 110799. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, C.M.; Snelgrove, R.J. Type 2 immunity: Expanding our view. Sci. Immunol. 2018, 3, eaat1604. [Google Scholar] [CrossRef] [PubMed]
- Clerici, M.; Shearer, G.M. A TH1-->TH2 switch is a critical step in the etiology of HIV infection. Immunol. Today. 1993, 14, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.D.; Herzenberg, L.A.; Vasquez, K.; Waltenbaugh, C. Glutathione levels in antigen-presenting cells modulate Th1 versus Th2 response patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 3071–3076. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Jia, J.; Yan, S.; Shen, H.; Zhu, P.; Yu, J. Paeoniflorin ameliorates ulcerative colitis by modulating the dendritic cell-mediated TH17/Treg balance. Inflammopharmacology 2020, 28, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Rana, M.N.; Lu, J.; Xue, E.; Ruan, J.; Liu, Y.; Zhang, L.; Dhar, R.; Li, Y.; Hu, Z.; Zhou, J.; et al. PDE9 Inhibitor PF-04447943 Attenuates DSS-Induced Colitis by Suppressing Oxidative Stress, Inflammation, and Regulating T-Cell Polarization. Front. Pharmacol. 2021, 12, 643215. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Cannavò, L.; Manti, S.; Rullo, I.; Buonocore, G.; Esposito, S.M.R.; Gitto, E. Pediatric Multisystem Syndrome Associated with SARS-CoV-2 (MIS-C): The Interplay of Oxidative Stress and Inflammation. Int. J. Mol. Sci. 2022, 23, 12836. [Google Scholar] [CrossRef] [PubMed]
- Kotani, K. Neutrophil/lymphocyte ratio and the oxidative stress burden. Can. J. Cardiol. 2015, 31, 365.e9. [Google Scholar] [CrossRef]
- Kulaksizoglu, B.; Kulaksizoglu, S. Relationship between neutrophil/lymphocyte ratio with oxidative stress and psychopathology in patients with schizophrenia. Neuropsychiatr. Dis. Treat. 2016, 12, 1999–2005. [Google Scholar] [CrossRef]
- Chua, R.X.Y.; Tay, M.J.Y.; Ooi, D.S.Q.; Siah, K.T.H.; Tham, E.H.; Shek, L.P.; Meaney, M.J.; Broekman, B.F.P.; Loo, E.X.L. Understanding the Link Between Allergy and Neurodevelopmental Disorders: A Current Review of Factors and Mechanisms. Front. Neurol. 2021, 11, 603571. [Google Scholar] [CrossRef]
- San Segundo-Acosta, P.; Montero-Calle, A.; Jernbom-Falk, A.; Alonso-Navarro, M.; Pin, E.; Andersson, E.; Hellström, C.; Sánchez-Martínez, M.; Rábano, A.; Solís-Fernández, G.; et al. Multiomics Profiling of Alzheimer’s Disease Serum for the Identification of Autoantibody Biomarkers. J. Proteome Res. 2021, 20, 5115–5130. [Google Scholar] [CrossRef] [PubMed]
- Huber, V.C.; Mondal, T.; Factor, S.A.; Seegal, R.F.; Lawrence, D.A. Serum antibodies from Parkinson’s disease patients react with neuronal membrane proteins from a mouse dopaminergic cell line and affect its dopamine expression. J. Neuroinflamm. 2006, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.S.Y.; Lee, B.; Lim, J.; Ma, D.R.; Goh, J.X.; Goh, S.Y.; Gulam, M.Y.; Koh, S.M.; Lee, W.W.; Feng, L.; et al. Parkinson’s Disease-Specific Autoantibodies against the Neuroprotective Co-Chaperone STIP1. Cells 2022, 11, 1649. [Google Scholar] [CrossRef] [PubMed]
- Mangalhara, K.C.; Varanasi, S.K.; Johnson, M.A.; Burns, M.J.; Rojas, G.R.; Esparza Moltó, P.B.; Sainz, A.G.; Tadepalle, N.; Abbott, K.L.; Mendiratta, G.; et al. Manipulating mitochondrial electron flow enhances tumor immunogenicity. Science 2023, 381, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Pangrazzi, L.; Balasco, L.; Bozzi, Y. Oxidative Stress and Immune System Dysfunction in Autism Spectrum Disorders. Int. J. Mol. Sci. 2020, 21, 3293. [Google Scholar] [CrossRef] [PubMed]
- Salinas, R.D.; Connolly, D.R.; Song, H. Invited Review: Epigenetics in neurodevelopment. Neuropathol. Appl. Neurobiol. 2020, 46, 6–27. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, D.A.; Genuis, S.J.; Frye, R.E.; Larson, G. Environmental toxicants and autism spectrum disorders: A systematic review. Transl. Psychiatry 2014, 4, e360. [Google Scholar] [CrossRef]
- Gu, F.; Chauhan, V.; Chauhan, A.; Gąssowska, M. Alterations in mitochondrial DNA copy number and the activities of electron transport chain complexes and pyruvate dehydrogenase in the frontal cortex from subjects with autism. Transl. Psychiatry 2013, 3, e2992013. [Google Scholar] [CrossRef]
- Siddiqui, M.F.; Elwell, C.; Johnson, M.H. Mitochondrial Dysfunction in Autism Spectrum Disorders. Autism Open Access. 2016, 6, 1000190. [Google Scholar] [CrossRef]
- Wang, P.Y.; Ma, J.; Kim, Y.C.; Son, A.Y.; Syed, A.M.; Liu, C.; Mori, M.P.; Huffstutler, R.D.; Stolinski, J.L.; Talagala, S.L.; et al. WASF3 disrupts mitochondrial respiration and may mediate exercise intolerance in myalgic encephalomyelitis/chronic fatigue syndrome. Proc. Natl. Acad. Sci. USA 2023, 120, e23027381202023. [Google Scholar] [CrossRef]
- Liu, X.; Hussain, R.; Mehmood, K.; Tang, Z.; Zhang, H.; Li, Y. Mitochondrial-Endoplasmic Reticulum Communication-Mediated Oxidative Stress and Autophagy. Biomed. Res. Int. 2022, 2022, 6459585. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, A.; Mendez, R.; Zhang, K. Cross talk between ER stress, oxidative stress, and inflammation in health and disease. Methods Mol. Biol. 2015, 1292, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Luca, M.; Di Mauro, M.; Di Mauro, M.; Luca, A. Gut Microbiota in Alzheimer’s Disease, Depression, and Type 2 Diabetes Mellitus: The Role of Oxidative Stress. Oxid. Med. Cell Longev. 2019, 2019, 4730539. [Google Scholar] [CrossRef]
- Liu, S.; Gao, J.; Zhu, M.; Liu, K.; Zhang, H.L. Gut Microbiota and Dysbiosis in Alzheimer’s Disease: Implications for Pathogenesis and Treatment. Mol. Neurobiol. 2020, 57, 5026–5043. [Google Scholar] [CrossRef] [PubMed]
- Ortega, A.D.S.V.; Szabó, C. Adverse Effects of Heat Stress on the Intestinal Integrity and Function of Pigs and the Mitigation Capacity of Dietary Antioxidants: A Review. Animals 2021, 11, 1135. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Cottrell, J.J.; Furness, J.B.; Rivera, L.R.; Kelly, F.W.; Wijesiriwardana, U.; Pustovit, R.V.; Fothergill, L.J.; Bravo, D.M.; Celi, P.; et al. Selenium and vitamin E together improve intestinal epithelial barrier function and alleviate oxidative stress in heat-stressed pigs. Exp. Physiol. 2016, 101, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Shaito, A.; Aramouni, K.; Assaf, R.; Parenti, A.; Orekhov, A.; Yazbi, A.E.; Pintus, G.; Eid, A.H. Oxidative Stress-Induced Endothelial Dysfunction in Cardiovascular Diseases. Front Biosci. 2022, 27, 105. [Google Scholar] [CrossRef]
- Förstermann, U.; Münzel, T. Endothelial Nitric Oxide Synthase in Vascular Disease: From Marvel to Menace. Circulation 2006, 113, 1708–1714. [Google Scholar] [CrossRef]
- Gimbrone, M.A., Jr.; García-Cardeña, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circulation Res. 2000, 118, 620–636. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef]
- Van Houten, B.; Santa-Gonzalez, G.A.; Camargo, M. DNA repair after oxidative stress: Current challenges. Curr. Opin. Toxicol. 2018, 7, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Alhmoud, J.F.; Woolley, J.F.; Al Moustafa, A.E.; Malki, M.I. DNA Damage/Repair Management in Cancers. Cancers 2020, 12, 1050. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Xu, Y.; Pang, D.; Zhao, Q.; Zhang, L.; Li, M.; Li, W.; Duan, G.; Zhu, C. Interrelation between homocysteine metabolism and the development of autism spectrum disorder in children. Front. Mol. Neurosci. 2022, 15, 947513. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qiu, S.; Shi, J.; Guo, Y.; Li, Z.; Cheng, Y.; Liu, Y. Association between MTHFR C677T/A1298C and susceptibility to autism spectrum disorders: A meta-analysis. BMC Pediatr. 2020, 20, 449. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Takai, D. The role of DNA methylation in mammalian epigenetics. Science 2019, 293, 1068–1070. [Google Scholar] [CrossRef] [PubMed]
- Duffney, L.J.; Valdez, P.; Tremblay, M.W.; Cao, X.; Montgomery, S.; McConkie-Rosell, A.; Jiang, Y.H. Epigenetics and autism spectrum disorder: A report of an autism case with mutation in H1 linker histone HIST1H1E and literature review. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2018, 177, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Wiśniowiecka-Kowalnik, B.; Nowakowska, B.A. Genetics and epigenetics of autism spectrum disorder-current evidence in the field. J. Appl. Genet. 2019, 60, 37–47. [Google Scholar] [CrossRef]
- Srikanth, K.; Kwon, A.; Lee, E.; Chung, H. Characterization of genes and pathways that respond to heat stress in Holstein calves through transcriptome analysis. Cell Stress Chaperones. 2017, 22, 29–42. [Google Scholar] [CrossRef]
- Bouchama, A.; Aziz, M.A.; Mahri, S.A.; Gabere, M.N.; Dlamy, M.A.; Mohammad, S.; Abbad, M.A.; Hussein, M. A Model of Exposure to Extreme Environmental Heat Uncovers the Human Transcriptome to Heat Stress. Sci. Rep. 2017, 7, 9429. [Google Scholar] [CrossRef]
- Heard, E.; Martienssen, R.A. Transgenerational epigenetic inheritance: Myths and mechanisms. Cell 2014, 157, 95–109. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Novati, G.; Pan, J.; Bycroft, C.; Žemgulytė, A.; Applebaum, T.; Pritzel, A.; Wong, L.H.; Zielinski, M.; Sargeant, T.; et al. Accurate proteome-wide missense variant effect prediction with AlphaMissense. Science 2023, 381, eadg7492. [Google Scholar] [CrossRef] [PubMed]
- Roufael, M.; Bitar, T.; Sacre, Y.; Andres, C.; Hleihel, W. Folate-Methionine Cycle Disruptions in ASD Patients and Possible Interventions: A Systematic Review. Genes 2023, 14, 709. [Google Scholar] [CrossRef] [PubMed]
- Berbert, A. Further comment on articles pertaining to: “Homocysteine as a potential predictor of cardiovascular risk in patients with COVID-19”. Med. Hypotheses 2021, 155, 110676. [Google Scholar] [CrossRef] [PubMed]
- Škovierová, H.; Vidomanová, E.; Mahmood, S.; Sopková, J.; Drgová, A.; Červeňová, T.; Halašová, E.; Lehotský, J. The Molecular and Cellular Effect of Homocysteine Metabolism Imbalance on Human Health. Int. J. Mol. Sci. 2016, 17, 1733. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.R.; Tyagi, S.C. Homocysteine and reactive oxygen species in metabolic syndrome, type 2 diabetes mellitus, and atheroscleropathy: The pleiotropic effects of folate supplementation. Nutr. J. 2004, 3, 4. [Google Scholar] [CrossRef]
- Ganguly, P.; Alam, S.F. Role of homocysteine in the development of cardiovascular disease. Nutr. J. 2015, 14, 6. [Google Scholar] [CrossRef]
- Hayden, M.R.; Tyagi, S.C. Impaired Folate-Mediated One-Carbon Metabolism in Type 2 Diabetes, Late-Onset Alzheimer’s Disease and Long COVID. Medicina 2021, 58, 16. [Google Scholar] [CrossRef]
- Ali, A.; Waly, M.I.; Al-Farsi, Y.M.; Essa, M.M.; Al-Sharbati, M.M.; Deth, R.C. Hyperhomocysteinemia among Omani autistic children: A case-control study. Acta Biochim. Pol. 2011, 58, 547–551. [Google Scholar] [CrossRef]
- Hocking, A.M.; Buckner, J.H. Genetic basis of defects in immune tolerance underlying the development of autoimmunity. Front. Immunol. 2022, 13, 972121. [Google Scholar] [CrossRef]
- Rylaarsdam, L.; Guemez-Gamboa, A. Genetic Causes and Modifiers of Autism Spectrum Disorder. Front. Cell Neurosci. 2019, 13, 385. [Google Scholar] [CrossRef] [PubMed]
- Nagar, S.D.; Nápoles, A.M.; Jordan, I.K.; Mariño-Ramírez, L. Socioeconomic deprivation and genetic ancestry interact to modify type 2 diabetes ethnic disparities in the United Kingdom. EClinicalMedicine. 2021, 37, 100960. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.B.; Wei, W.Q.; Weeraratne, D.; Frisse, M.E.; Misulis, K.; Rhee, K.; Zhao, J.; Snowdon, J.L. Precision Medicine, AI, and the Future of Personalized Health Care. Clin. Transl. Sci. 2021, 14, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Buccellato, F.R.; D’Anca, M.; Fenoglio, C.; Scarpini, E.; Galimberti, D. Role of Oxidative Damage in Alzheimer’s Disease and Neurodegeneration: From Pathogenic Mechanisms to Biomarker Discovery. Antioxidants 2021, 10, 1353. [Google Scholar] [CrossRef] [PubMed]
- Luca, M.; Luca, A.; Calandra, C. The Role of Oxidative Damage in the Pathogenesis and Progression of Alzheimer’s Disease and Vascular Dementia. Oxidative Med. Cell. Longev. 2015, 504678. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.E.; Paek, S.H. Mitochondrial Dysfunction in Parkinson’s Disease. Exp. Neurobiol. 2015, 24, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Shehata, M.F. Role of the IRS-1 and/or -2 in the pathogenesis of insulin resistance in Dahl salt-sensitive (S) rats. Heart Int. 2009, 4, e62009. [Google Scholar] [CrossRef] [PubMed]
- Vaseghi, H.; Jadali, Z. Th1/Th2 cytokines in Type 1 diabetes: Relation to duration of disease and gender. Indian. J. Endocrinol. Metab. 2016, 20, 312–316. [Google Scholar] [CrossRef]
- Moon, J.; Kitty, I.; Renata, K.; Qin, S.; Zhao, F.; Kim, W. DNA Damage and Its Role in Cancer Therapeutics. Int. J. Mol. Sci. 2023, 24, 4741. [Google Scholar] [CrossRef]
- Sun, H.J.; Wu, Z.Y.; Nie, X.W.; Bian, J.S. Role of Endothelial Dysfunction in Cardiovascular Diseases: The Link Between Inflammation and Hydrogen Sulfide. Front. Pharmacol. 2020, 10, 1568. [Google Scholar] [CrossRef]
- Farrugia, M.; Baron, B. The role of TNF-α in rheumatoid arthritis: A focus on regulatory T cells. J. Clin. Translat Res. 2016, 2, 84–90. [Google Scholar] [CrossRef]
- Ameer, M.A.; Chaudhry, H.; Mushtaq, J.; Khan, O.S.; Babar, M.; Hashim, T.; Zeb, S.; Tariq, M.A.; Patlolla, S.R.; Ali, J.; et al. An Overview of Systemic Lupus Erythematosus (SLE) Pathogenesis, Classification, and Management. Cureus 2022, 14, e30330. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, A.M.; Jones, D.P.; Brown, L.A. Glutathione redox control of asthma: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2012, 17, 375–408. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.; Hahn, B.H. Atherosclerosis and systemic lupus erythematosus: Mechanistic basis of the association. Curr. Opin. Immunol. 2007, 19, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Mehrabadi, M.E.; Hemmati, R.; Tashakor, A.; Homaei, A.; Yousefzadeh, M.; Hemati, K.; Hosseinkhani, S. Induced dysregulation of ACE2 by SARS-CoV-2 plays a key role in COVID-19 severity. Biomed. Pharmacother. 2021, 137, 111363. [Google Scholar] [CrossRef] [PubMed]
- Vassiliou, A.G.; Kotanidou, A.; Dimopoulou, I.; Orfanos, S.E. Endothelial Damage in Acute Respiratory Distress Syndrome. Int. J. Mol. Sci. 2020, 21, 8793. [Google Scholar] [CrossRef]
- Wickenhagen, A.; Sugrue, E.; Lytras, S.; Kuchi, S.; Noerenberg, M.; Turnbull, M.L.; Loney, C.; Herder, V.; Allan, J.; Jarmson, I.; et al. A prenylated dsRNA sensor protects against severe COVID-19. Science 2021, 374, eabj3624. [Google Scholar] [CrossRef]
- Roberts, C.K.; Sindhu, K.K. Oxidative stress and metabolic syndrome. Life Sci. 2009, 84, 705–712. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef]
- Perticone, F.; Ceravolo, R.; Candigliota, M.; Ventura, G.; Iacopino, S.; Sinopoli, F.; Mattioli, P.L. Obesity and body fat distribution induce endothelial dysfunction by oxidative stress: Protective effect of vitamin C. Diabetes 2001, 50, 159–165. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Michailidou, Z.; Gomez-Salazar, M.; Alexaki, V.I. Innate Immune Cells in the Adipose Tissue in Health and Metabolic Disease. J. Innate Immun. 2022, 14, 4–30. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Zmora, N.; Adolph, T.E.; Elinav, E. The intestinal microbiota fuelling metabolic inflammation. Nat. Rev. Immunol. 2020, 20, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Gurung, M.; Bruce, A.M.; Brown, K.; Bobe, G.; Sharpton, T.J.; Shulzhenko, N.; Maier, C.S.; Stevens, J.F.; Gombart, A.F.; Morgun, A. Reducing gut microbiome-driven adipose tissue inflammation alleviates metabolic syndrome. Microbiome 2023, 11, 208. [Google Scholar] [CrossRef]
- Prasun, P. Mitochondrial dysfunction in metabolic syndrome. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165838. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; R PRM, P.; Brown, L.; Ayyappan, P.; Raghu, K.G. Endoplasmic reticulum stress: A master regulator of metabolic syndrome. Eur. J. Pharmacol. 2019, 860, 172553. [Google Scholar] [CrossRef] [PubMed]
- Condelli, V.; Crispo, F.; Pietrafesa, M.; Lettini, G.; Matassa, D.S.; Esposito, F.; Landriscina, M.; Maddalena, F. HSP90 Molecular Chaperones, Metabolic Rewiring, and Epigenetics: Impact on Tumor Progression and Perspective for Anticancer Therapy. Cells 2019, 8, 532. [Google Scholar] [CrossRef] [PubMed]
- Zuehlke, A.; Johnson, J.L. Hsp90 and co-chaperones twist the functions of diverse client proteins. Biopolymers 2010, 93, 211–217. [Google Scholar] [CrossRef]
- Bahr, T.; Katuri, J.; Liang, T.; Bai, Y. Mitochondrial chaperones in human health and disease. Free Radic. Biol. Med. 2022, 179, 363–374. [Google Scholar] [CrossRef]
- Castro, J.P.; Wardelmann, K.; Grune, T.; Kleinridders, A. Mitochondrial Chaperones in the Brain: Safeguarding Brain Health and Metabolism? Front Endocrinol. 2018, 9, 196. [Google Scholar] [CrossRef]
- Masenga, S.K.; Kabwe, L.S.; Chakulya, M.; Kirabo, A. Mechanisms of Oxidative Stress in Metabolic Syndrome. Int. J. Mol. Sci. 2023, 24, 7898. [Google Scholar] [CrossRef] [PubMed]
- Fahed, G.; Aoun, L.; Bou Zerdan, M.; Allam, S.; Bou Zerdan, M.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Pappan, N.; Rehman, A. Dyslipidemia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Monserrat-Mesquida, M.; Quetglas-Llabrés, M.; Capó, X.; Bouzas, C.; Mateos, D.; Pons, A.; Tur, J.A.; Sureda, A. Metabolic Syndrome is Associated with Oxidative Stress and Proinflammatory State. Antioxidants 2020, 9, 236. [Google Scholar] [CrossRef]
- Pahwa, R.; Goyal, A.; Jialal, I. Chronic Inflammation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Stanciu, S.; Rusu, E.; Miricescu, D.; Radu, A.C.; Axinia, B.; Vrabie, A.M.; Ionescu, R.; Jinga, M.; Sirbu, C.A. Links between Metabolic Syndrome and Hypertension: The Relationship with the Current Antidiabetic Drugs. Metabolites 2023, 13, 87. [Google Scholar] [CrossRef]
- Natour, S.; Damri, O.; Agam, G. The Effect of Global Warming on Complex Disorders (Mental Disorders, Primary Hypertension, and Type 2 Diabetes). Int. J. Environ. Res. Public Health 2022, 19, 9398. [Google Scholar] [CrossRef]
- Ryan, K.K. Stress and Metabolic Disease. In Committee on Population; Division of Behavioral and Social Sciences and Education, National Research Council, Sociality, Hierarchy, Health: Comparative Biodemography: A Collection of Papers; Weinstein, M., Lane, M.A., Eds.; National Academies Press: Washington, DC, USA, 2014. [Google Scholar]
- Castro-Barquero, S.; Ruiz-León, A.M.; Sierra-Pérez, M.; Estruch, R.; Casas, R. Dietary Strategies for Metabolic Syndrome: A Comprehensive Review. Nutrients 2020, 12, 2983. [Google Scholar] [CrossRef]
| Pathology | Mechanistic Connection | Gene/Protein | Reference |
|---|---|---|---|
| Alzheimer’s Disease | Oxidative damage | APOE, APP, PSEN1, PSEN2 | Buccellato et al. [116] Luca et al. [117] |
| Parkinson’s Disease | Mitochondrial dysfunction | DJ-1, PINK1, PARKIN | Moon and Paek [118] |
| Type 2 Diabetes | Insulin resistance, Mitochondrial Dysfunction | IRS1, Nrf2 | Shehata [119] |
| Type 1 Diabetes | Dysregulation of Th1/Th2 balance and immune cell activation; autoimmunity | Th1 and Th2 cells Tumor Necrosis Factor-alpha (TNF-α) | Vaseghi and Jadali [120] |
| Cancer | DNA damage, Angiogenesis Antioxidant Defense | TP53, BRCA1, VEGF, NRF2 | Moon et al. [121] |
| Cardiovascular Disease | Oxidative modification of lipids and endothelial dysfunction | NOS3, SOD, HO-1 | Sun et al. [122] |
| Rheumatoid Arthritis | Inflammation | TNF-alpha, NF-kB, etc. | Farrugia and Baron [123] |
| Autism Spectrum Disorder (ASD) | Neuroinflammation | NLGN3, NLGN4, SHANK3, etc. | Pangrazzi et al. [77] |
| Systemic Lupus Erythematosus (SLE) | Excessive ROS production | IFN-α, IL-1, IL-6, and TNF-α | Ameer et al. [124] |
| Asthma | Regulation of oxidative stress and inflammation | GPX1 (Glutathione Peroxidase 1), NFE2L2 (Nuclear Factor Erythroid 2 Like 2) | Fitzpatrick et al. [125] |
| Atherosclerosis | Oxidative Modification of Lipids | Low-density Lipoproteins (LDL) Scavenger Receptors (e.g., SR-A, CD36) | McMahon and Hahn [126] |
| Severe Acute Respiratory Syndrome (SARS) | Endothelial Dysfunction Coagulation Imbalance ACE2 Receptor Modulation | Endothelin-1 (ET-1) Plasminogen Activator Inhibitor-1 (PAI-1) Angiotensin-Converting Enzyme 2 (ACE2) Angiotensin II (Ang II) | Mehrabadi et al. [127] Vassiliou et al. [128] |
| Environmental Stressor | Effect on Metabolic Syndrome | Reference |
|---|---|---|
| Climate Change and Temperature Rise | May exacerbate metabolic dysfunction through heat stress, disrupted sleep, and increased inflammation. | Natour et al. [150] |
| Psychological Stress | Can lead to hormonal changes and unhealthy coping behaviors, contributing to metabolic syndrome. | Ryan. [151] |
| Diet and Nutrition | Poor dietary choices in response to stressors can contribute to metabolic syndrome. | Castro-Barquero et al. [152] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heidari, H.; Lawrence, D.A. Climate Stressors and Physiological Dysregulations: Mechanistic Connections to Pathologies. Int. J. Environ. Res. Public Health 2024, 21, 28. https://doi.org/10.3390/ijerph21010028
Heidari H, Lawrence DA. Climate Stressors and Physiological Dysregulations: Mechanistic Connections to Pathologies. International Journal of Environmental Research and Public Health. 2024; 21(1):28. https://doi.org/10.3390/ijerph21010028
Chicago/Turabian StyleHeidari, Hajar, and David A. Lawrence. 2024. "Climate Stressors and Physiological Dysregulations: Mechanistic Connections to Pathologies" International Journal of Environmental Research and Public Health 21, no. 1: 28. https://doi.org/10.3390/ijerph21010028
APA StyleHeidari, H., & Lawrence, D. A. (2024). Climate Stressors and Physiological Dysregulations: Mechanistic Connections to Pathologies. International Journal of Environmental Research and Public Health, 21(1), 28. https://doi.org/10.3390/ijerph21010028