
Exposure of the Gestating Mother to Sympathetic Stress Modifies the Cardiovascular Function of the Progeny in Male Rats

 ,
,
Abstract
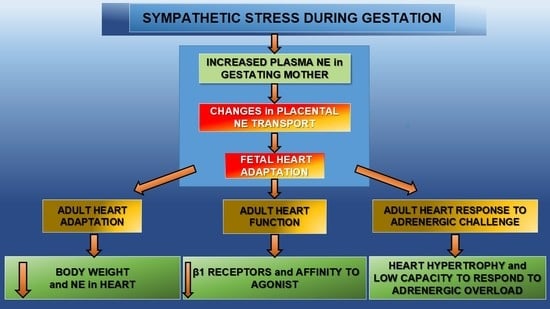
1. Introduction
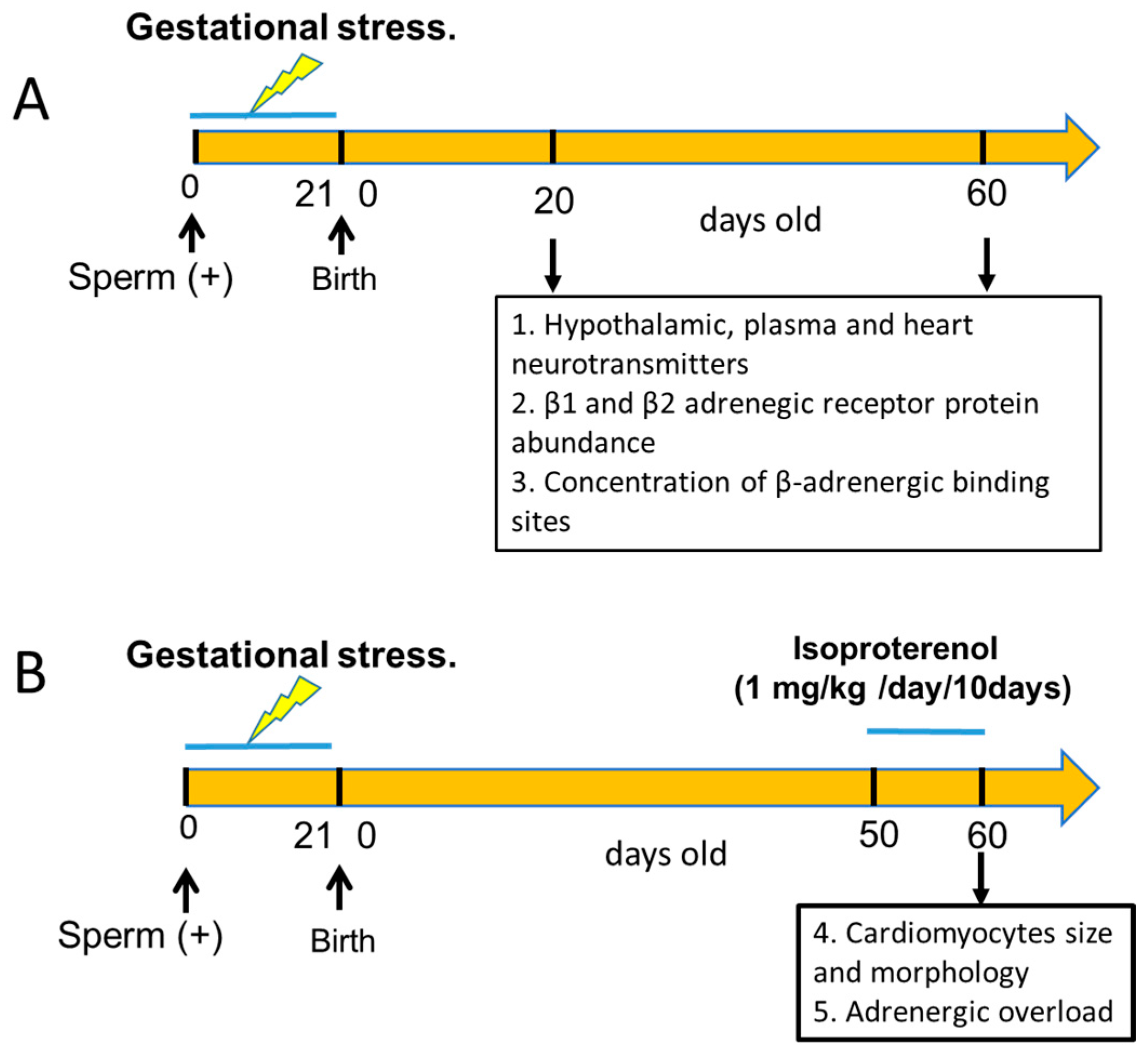
2. Animals and Experimental Design
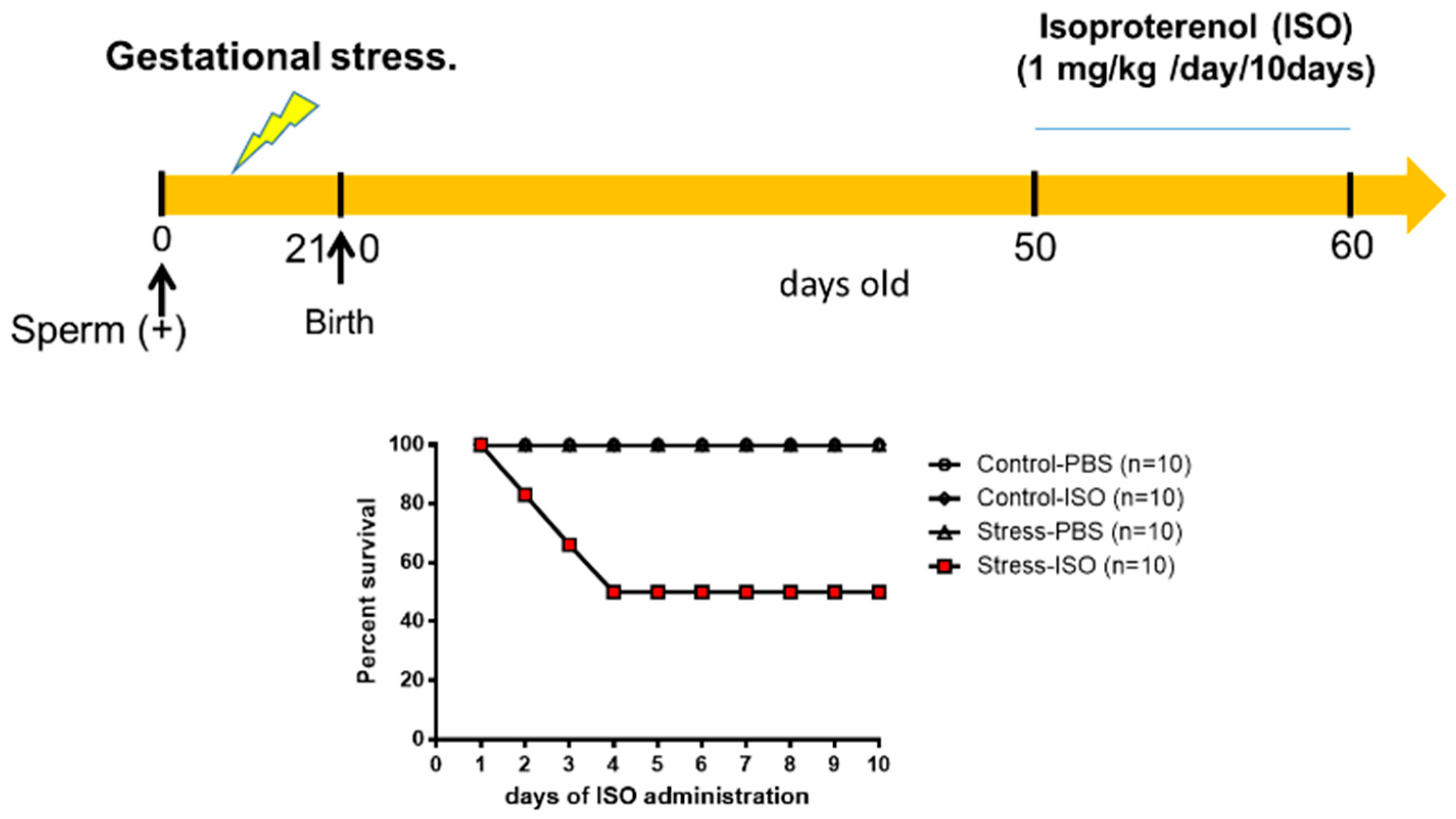
2.1. Gestational Stress Induction
2.2. Adrenergic Overload Protocol
2.3. Quantification of NE Levels
2.4. Quantification of Serotonin in the Medial Basal Hypothalamus (MBH) by ELISA
2.5. Quantification of Plasma Levels of Corticosterone by ELISA
2.6. Binding of [3H]dihydroalprenolol to Cardiac Membranes
2.6.1. Preparation of the Crude Membrane Fraction
2.6.2. Assay for β-Adrenergic Receptors
2.7. Determination of Cardiac β1- and β2-Adrenergic Receptors by Western Blot
2.8. Morphometric Analysis
2.9. Statistical Analysis
3. Results
3.1. Animal and Heart Relative Weight
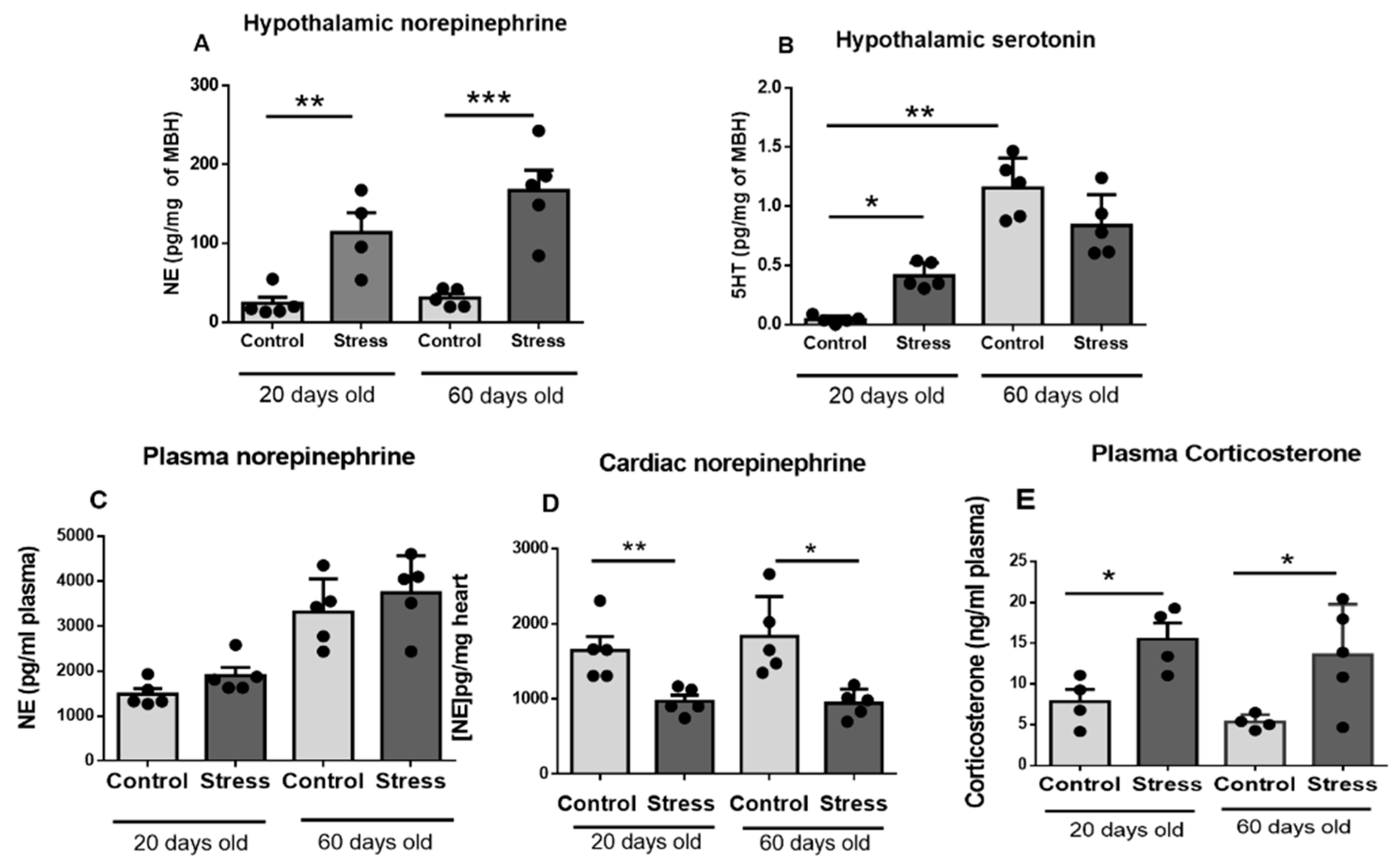
3.2. Hypothalamic, Plasma, and Cardiac Neurotransmitters and Plasma Corticosterone
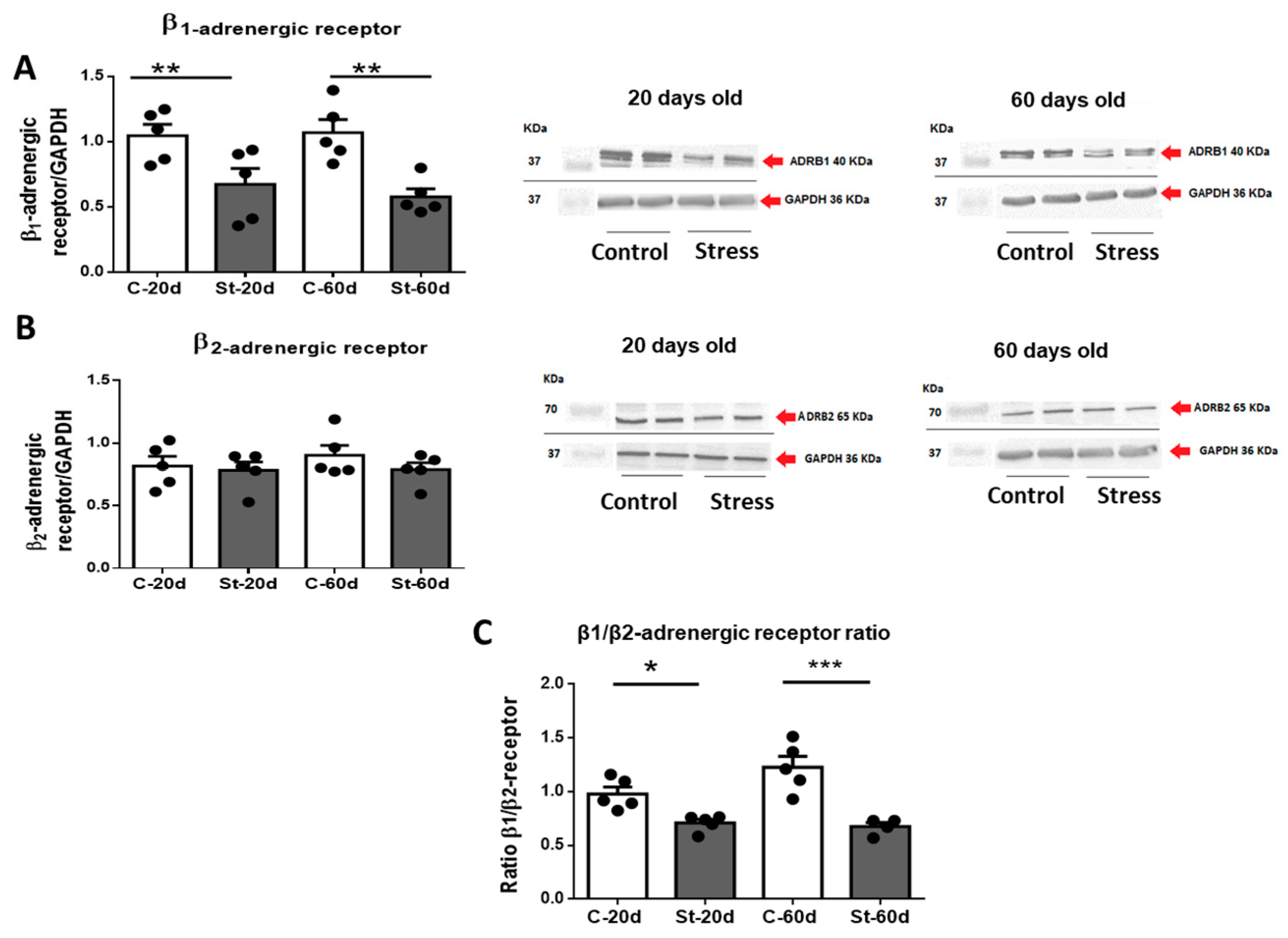
3.3. Changes in β1 and β2 Adrenergic Receptor Protein Abundance
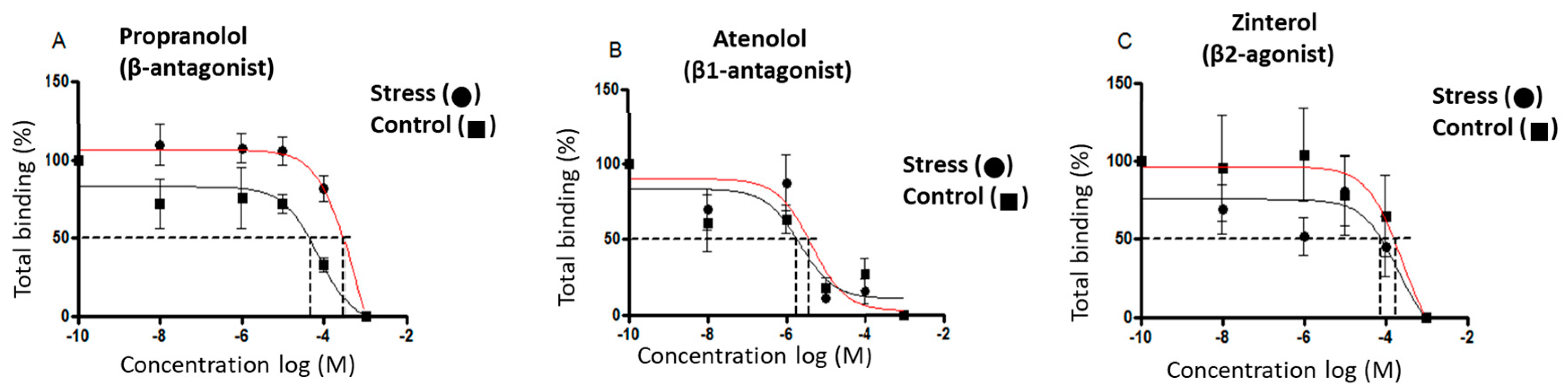
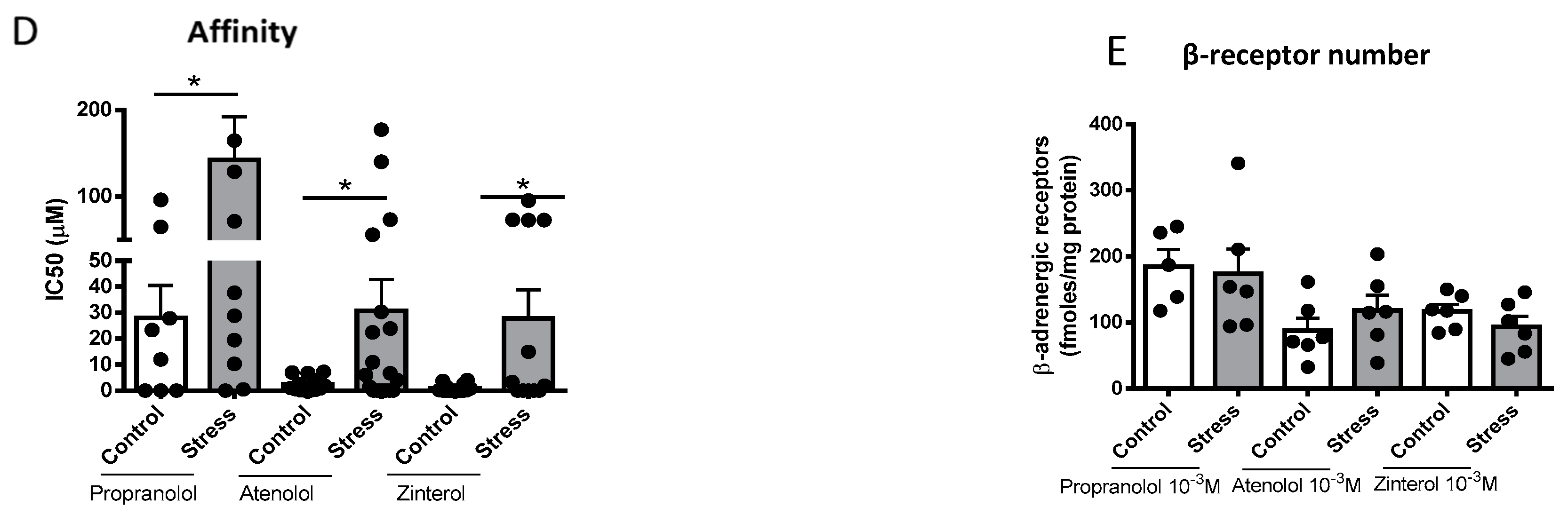
3.4. Concentration of β-Adrenergic Binding Sites
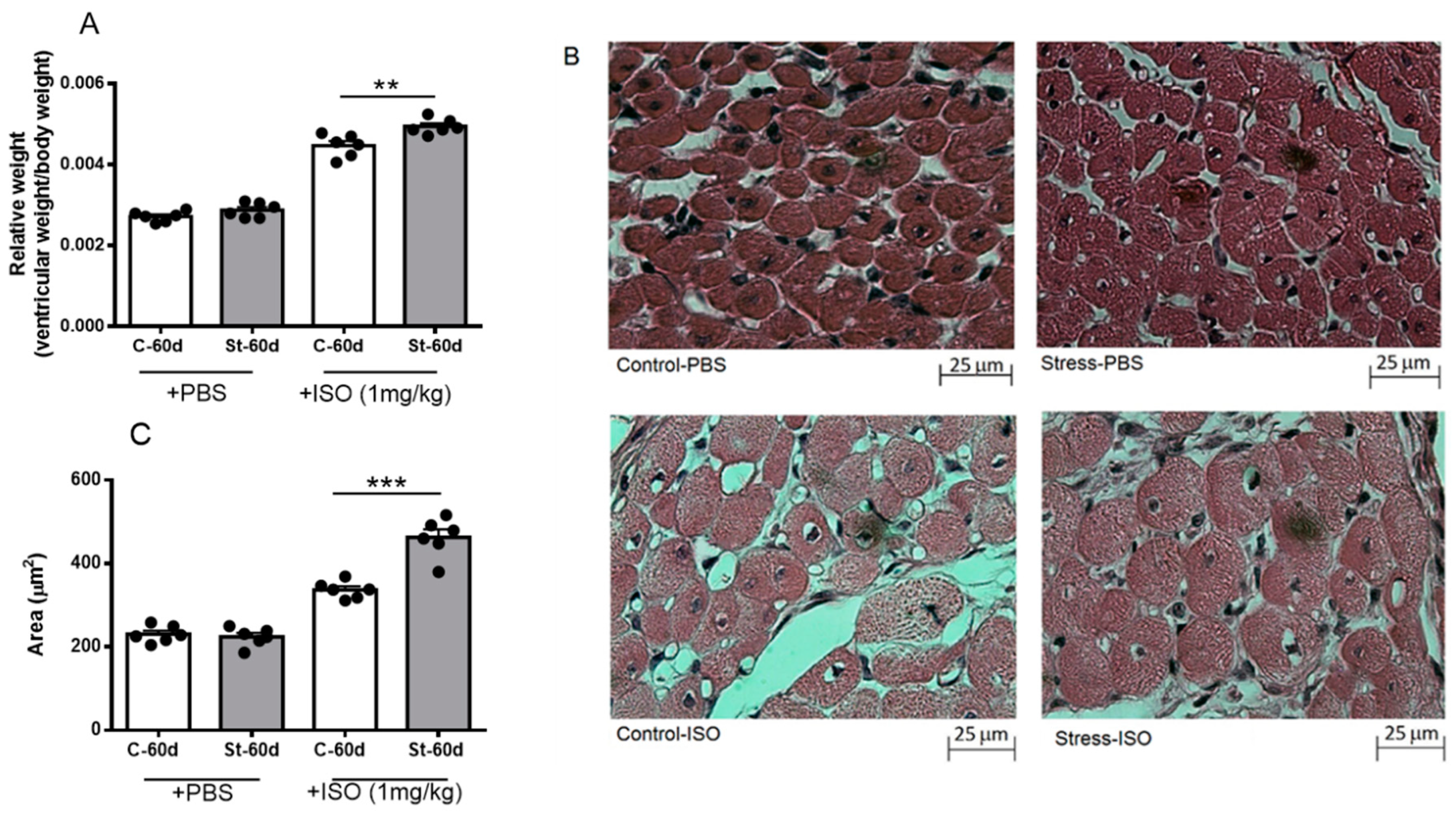
3.5. Effects of Isoproterenol on Ventricle Size and Cardiomyocyte Area in Gestationally Stressed Rats
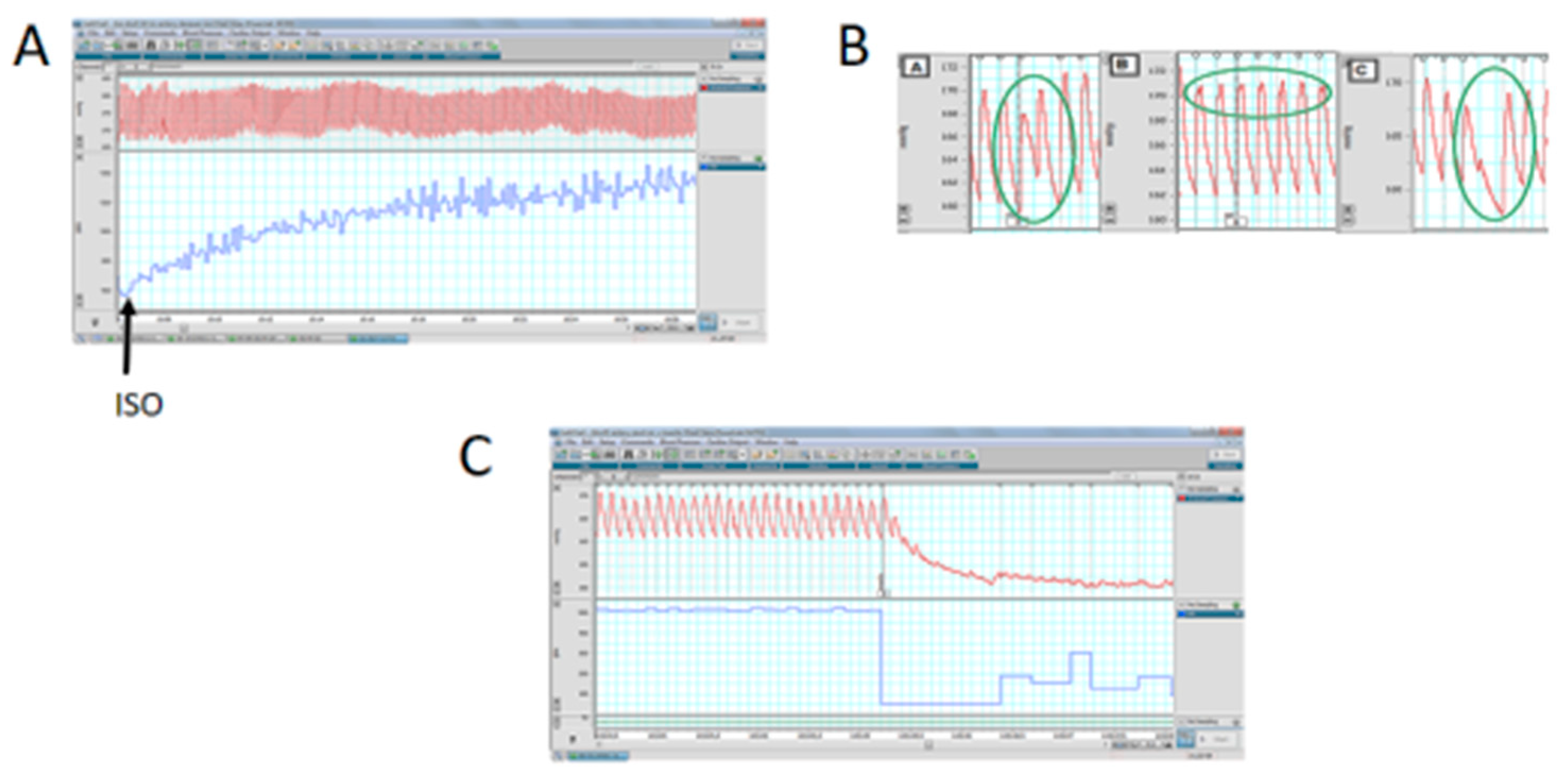
3.6. Cardiac Functional Parameters in Stressed Offspring Challenged with Isoproterenol Overload
4. Discussion
Effects of Gestational Stress in the Heart
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cottrell, E.C.; Seckl, J.R. Prenatal stress, glucocorticoids and the programming of adult disease. Front. Behav. Neurosci. 2009, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Kajantie, E. Fetal origins of stress-related adult disease. Ann. N. Y. Acad. Sci. 2006, 1083, 11–27. [Google Scholar] [CrossRef]
- Hales, C.N.; Barker, D.J. The thrifty phenotype hypothesis. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef]
- Godfrey, K.M.; Barker, D.J. Fetal programming and adult health. Public. Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef]
- Phillips, D.I.; Jones, A.; Goulden, P.A. Birth weight, stress, and the metabolic syndrome in adult life. Ann. N. Y. Acad. Sci. 2006, 1083, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.; Zimanyi, M.A.; Black, M.J. Effect of maternal protein restriction in rats on cardiac fibrosis and capillarization in adulthood. Pediatr. Res. 2006, 60, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Twinn, D.S.; Ekizoglou, S.; Wayman, A.; Petry, C.J.; Ozanne, S.E. Maternal low-protein diet programs cardiac beta-adrenergic response and signaling in 3-mo-old male offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R429–R436. [Google Scholar] [CrossRef] [PubMed]
- Igosheva, N.; Klimova, O.; Anishchenko, T.; Glover, V. Prenatal stress alters cardiovascular responses in adult rats. J. Physiol. 2004, 557 Pt 1, 273–285. [Google Scholar] [CrossRef]
- Piquer, B.; Fonseca, J.L.; Lara, H.E. Gestational stress, placental norepinephrine transporter and offspring fertility. Reproduction 2017, 153, 147–155. [Google Scholar] [CrossRef]
- Goldstein, D.S.; Kopin, I.J. Adrenomedullary, adrenocortical, and sympathoneural responses to stressors: A meta-analysis. Endocr Regul. 2008, 42, 111–119. [Google Scholar]
- Pacak, K.; Palkovits, M.; Yadid, G.; Kvetnansky, R.; Kopin, I.J.; Goldstein, D.S. Heterogeneous neurochemical responses to different stressors: A test of Selye’s doctrine of nonspecificity. Am. J. Physiol. 1998, 275 Pt 2, R1247–R1255. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; De Ferranti, S.; Després, J.P.; Fullerton, H.J.; et al. Heart Disease and Stroke Statistics-2016 Update: A Report From the American Heart Association. Circulation 2016, 133, e38–e360. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.R.; Fillenz, M.; Stanford, C. Noradrenaline release in rats during prolonged cold-stress and repeated swim-stress. Br. J. Pharmacol. 1979, 66, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Dorfman, M.; Arancibia, S.; Fiedler, J.L.; Lara, H.E. Chronic intermittent cold stress activates ovarian sympathetic nerves and modifies ovarian follicular development in the rat. Biol. Reprod. 2003, 68, 2038–2043. [Google Scholar] [CrossRef]
- Dorfman, M.; Ramirez, V.D.; Stener-Victorin, E.; Lara, H.E. Chronic-intermittent cold stress in rats induces selective ovarian insulin resistance. Biol. Reprod. 2009, 80, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Barra, R.; Cruz, G.; Mayerhofer, A.; Paredes, A.; Lara, H.E. Maternal sympathetic stress impairs follicular development and puberty of the offspring. Reproduction 2014, 148, 137–145. [Google Scholar] [CrossRef]
- Piquer, B.; Ruz, F.; Barra, R.; Lara, H.E. Gestational Sympathetic Stress Programs the Fertility of Offspring: A Rat Multi-Generation Study. Int. J. Environ. Res. Public Health 2022, 19, 3044. [Google Scholar] [CrossRef]
- Lian, S.; Wang, D.; Xu, B.; Guo, W.; Wang, L.; Li, W.; Ji, H.; Wang, J.; Kong, F.; Zhen, L.; et al. Prenatal cold stress: Effect on maternal hippocampus and offspring behavior in rats. Behav Brain Res. 2018, 346, 1–10. [Google Scholar] [CrossRef]
- Sun, D.; Chen, K.; Wang, J.; Zhou, L.; Zeng, C. In-utero cold stress causes elevation of blood pressure via impaired vascular dopamine D1 receptor in offspring. Clin. Exp. Hypertens. 2020, 42, 99–104. [Google Scholar] [CrossRef]
- AVMA. American Veterinary Medical Association Guidelines for the Euthanasia of Animals, 2020th ed.; American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Ayala, P.; Montenegro, J.; Vivar, R.; Letelier, A.; Urroz, P.A.; Copaja, M.; Pivet, D.; Humeres, C.; Troncoso, R.; Vicencio, J.M.; et al. Attenuation of endoplasmic reticulum stress using the chemical chaperone 4-phenylbutyric acid prevents cardiac fibrosis induced by isoproterenol. Exp. Mol. Pathol. 2012, 92, 97–104. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford method for protein quantitation. Methods Mol. Biol. 1994, 32, 9–15. [Google Scholar] [PubMed]
- Barria, A.; Leyton, V.; Ojeda, S.R.; Lara, H.E. Ovarian steroidal response to gonadotropins and beta-adrenergic stimulation is enhanced in polycystic ovary syndrome: Role of sympathetic innervation. Endocrinology 1993, 133, 2696–2703. [Google Scholar] [CrossRef] [PubMed]
- Lara, H.E.; Porcile, A.; Espinoza, J.; Romero, C.; Luza, S.M.; Fuhrer, J.; Miranda, C.; Roblero, L. Release of norepinephrine from human ovary: Coupling to steroidogenic response. Endocrine 2001, 15, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Fushimi, K.; Kouchi, H.; Mihara, K.; Miyazaki, M.; Ohe, T.; Namba, M. Inhibitory effects of antioxidants on neonatal rat cardiac myocyte hypertrophy induced by tumor necrosis factor-alpha and angiotensin, II. Circulation 1998, 98, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Zar, J. Biostatistical Analysis, 2nd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1984; 5p. [Google Scholar]
- Sengupta, P. The Laboratory Rat: Relating Its Age With Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Ricu, M.; Paredes, A.; Greiner, M.; Ojeda, S.R.; Lara, H.E. Functional development of the ovarian noradrenergic innervation. Endocrinology 2008, 149, 50–56. [Google Scholar] [CrossRef][Green Version]
- Burnstock, G.; Verkhratsky, A. Vas deferens--a model used to establish sympathetic cotransmission. Trends Pharmacol. Sci. 2010, 31, 131–139. [Google Scholar] [CrossRef]
- Bernuci, M.P.; Szawka, R.E.; Helena, C.V.; Leite, C.M.; Lara, H.E.; Anselmo-Franci, J.A. Locus coeruleus mediates cold stress-induced polycystic ovary in rats. Endocrinology 2008, 149, 2907–2916. [Google Scholar] [CrossRef]
- Cayupe, B.; Morgan, C.; Puentes, G.; Valladares, L.; Burgos, H.; Castillo, A.; Hernández, A.; Constandil, L.; Ríos, M.; Sáez-Briones, P.; et al. Hypertension in Prenatally Undernourished Young-Adult Rats Is Maintained by Tonic Reciprocal Paraventricular-Coerulear Excitatory Interactions. Molecules 2021, 26, 3568. [Google Scholar] [CrossRef]
- Jara, P.; Rage, F.; Dorfman, M.; Grouselle, D.; Barra, R.; Arancibia, S.; Lara, H.E. Cold-induced glutamate release in vivo from the magnocellular region of the paraventricular nucleus is involved in ovarian sympathetic activation. J. Neuroendocrinol. 2010, 22, 979–986. [Google Scholar] [CrossRef]
- Spergel, D.J. Modulation of Gonadotropin-Releasing Hormone Neuron Activity and Secretion in Mice by Non-peptide Neurotransmitters, Gasotransmitters, and Gliotransmitters. Front. Endocrinol. 2019, 10, 329. [Google Scholar] [CrossRef]
- Muir, J.L.; Pfister, H.P. Psychological stress and oxytocin treatment during pregnancy affect central norepinephrine, dopamine and serotonin in lactating rats. Int. J. Neurosci. 1989, 48, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Oberlander, T.F. In-utero Selective Serotonin Reuptake Inhibitor Antidepressant Exposure: Fetal Programing and Developmental Interactions with Context. Clin. Pharmacol. Ther. 2018, 104, 616–618. [Google Scholar] [CrossRef]
- Wang, J.; Gareri, C.; Rockman, H.A. G-Protein-Coupled Receptors in Heart Disease. Circ. Res. 2018, 123, 716–735. [Google Scholar] [CrossRef] [PubMed]
- Bristow, M.R.; Altman, N.L. Heart Rate in Preserved Ejection Fraction Heart Failure. JACC Heart Fail. 2017, 5, 792–794. [Google Scholar] [CrossRef] [PubMed]
- Jevdjovic, T.; Dakic, T.; Kopanja, S.; Lakic, I.; Vujovic, P.; Jasnic, N.; Djordjevic, J. Sex-Related Effects of Prenatal Stress on Region-Specific Expression of Monoamine Oxidase A and beta Adrenergic Receptors in Rat Hearts. Arq. Bras. Cardiol. 2019, 112, 67–75. [Google Scholar]
- Galvez, A.; Paredes, A.; Fiedler, J.L.; Venegas, M.; Lara, H.E. Effects of adrenalectomy on the stress-induced changes in ovarian sympathetic tone in the rat. Endocrine 1999, 10, 131–135. [Google Scholar] [CrossRef]
- Elmes, M.J.; Haase, A.; Gardner, D.S.; Langley-Evans, S.C. Sex differences in sensitivity to beta-adrenergic agonist isoproterenol in the isolated adult rat heart following prenatal protein restriction. Br. J. Nutr. 2009, 101, 725–734. [Google Scholar] [CrossRef]
- Luna, S.L.; Neuman, S.; Aguilera, J.; Brown, D.I.; Lara, H.E. In vivo beta-adrenergic blockade by propranolol prevents isoproterenol-induced polycystic ovary in adult rats. Horm. Metab. Res. 2012, 44, 676–681. [Google Scholar]
- Bristow, M.R.; Ginsburg, R.; Umans, V.; Fowler, M.; Minobe, W.; Rasmussen, R.; Zera, P.; Menlove, R.; Shah, P.; Jamieson, S. Beta 1- and beta 2-adrenergic-receptor subpopulations in nonfailing and failing human ventricular myocardium: Coupling of both receptor subtypes to muscle contraction and selective beta 1-receptor down-regulation in heart failure. Circ. Res. 1986, 59, 297–309. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Gestational Stress | |
|---|---|---|
| Days of gestation | 21.8 ± 0.2 (n = 8) | 21.9 ± 0.1 (n = 12) |
| Pups per mother | 14.3 ± 0.3 (n = 8 mothers, 100 pups) | 12.1 ± 1.1 (n = 12 mothers, 133 pups) |
| Number of pups by sex | ||
| Females | 7.0 ± 0.7 (n = 49 pups) | 6.2 ± 0.7 (n = 68 pups) |
| Males | 7.2 ± 0.8 (n = 51 pups) | 5.9 ± 0.8 (n = 65 pups) |
| Males rats used in this study | ||
| Weight of males | ||
| 20 days old | 53.7 ± 3.0 (n = 9) | 54.8 ± 1.4 (n = 10) |
| 60 days old | 361.3 ± 11.1 (n = 34) | 373.4 ± 12.4 (n = 36) |
| Heart weight/rat weight | ||
| 20 days old | 0.005 ± 0.0002 (n = 9) | 0.005 ± 0.0001 (n = 10) |
| 60 days old | 0.003 ± 0.0001 (n = 14) | 0.003 ± 0.0001 (n = 16) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piquer, B.; Olmos, D.; Flores, A.; Barra, R.; Bahamondes, G.; Diaz-Araya, G.; Lara, H.E. Exposure of the Gestating Mother to Sympathetic Stress Modifies the Cardiovascular Function of the Progeny in Male Rats. Int. J. Environ. Res. Public Health 2023, 20, 4285. https://doi.org/10.3390/ijerph20054285
Piquer B, Olmos D, Flores A, Barra R, Bahamondes G, Diaz-Araya G, Lara HE. Exposure of the Gestating Mother to Sympathetic Stress Modifies the Cardiovascular Function of the Progeny in Male Rats. International Journal of Environmental Research and Public Health. 2023; 20(5):4285. https://doi.org/10.3390/ijerph20054285
Chicago/Turabian StylePiquer, Beatriz, Diandra Olmos, Andrea Flores, Rafael Barra, Gabriela Bahamondes, Guillermo Diaz-Araya, and Hernan E. Lara. 2023. "Exposure of the Gestating Mother to Sympathetic Stress Modifies the Cardiovascular Function of the Progeny in Male Rats" International Journal of Environmental Research and Public Health 20, no. 5: 4285. https://doi.org/10.3390/ijerph20054285
APA StylePiquer, B., Olmos, D., Flores, A., Barra, R., Bahamondes, G., Diaz-Araya, G., & Lara, H. E. (2023). Exposure of the Gestating Mother to Sympathetic Stress Modifies the Cardiovascular Function of the Progeny in Male Rats. International Journal of Environmental Research and Public Health, 20(5), 4285. https://doi.org/10.3390/ijerph20054285