
Anode Modification with Fe2O3 Affects the Anode Microbiome and Improves Energy Generation in Microbial Fuel Cells Powered by Wastewater




Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up
2.2. Electrochemical Analyses
2.3. Molecular Analyses
2.4. Statistical Analyses
3. Results and Discussion
3.1. Analysis of the Electrode Surface
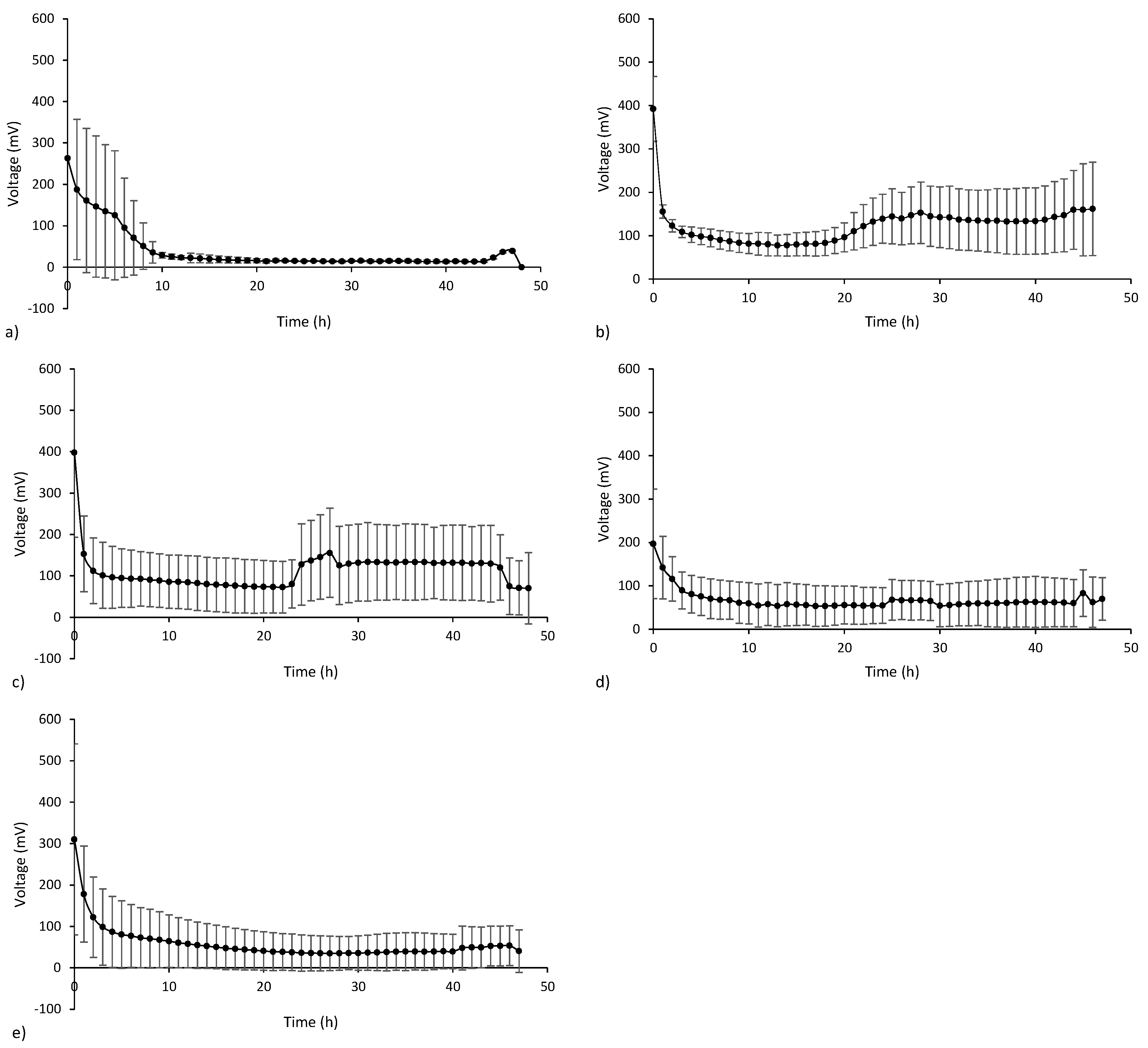
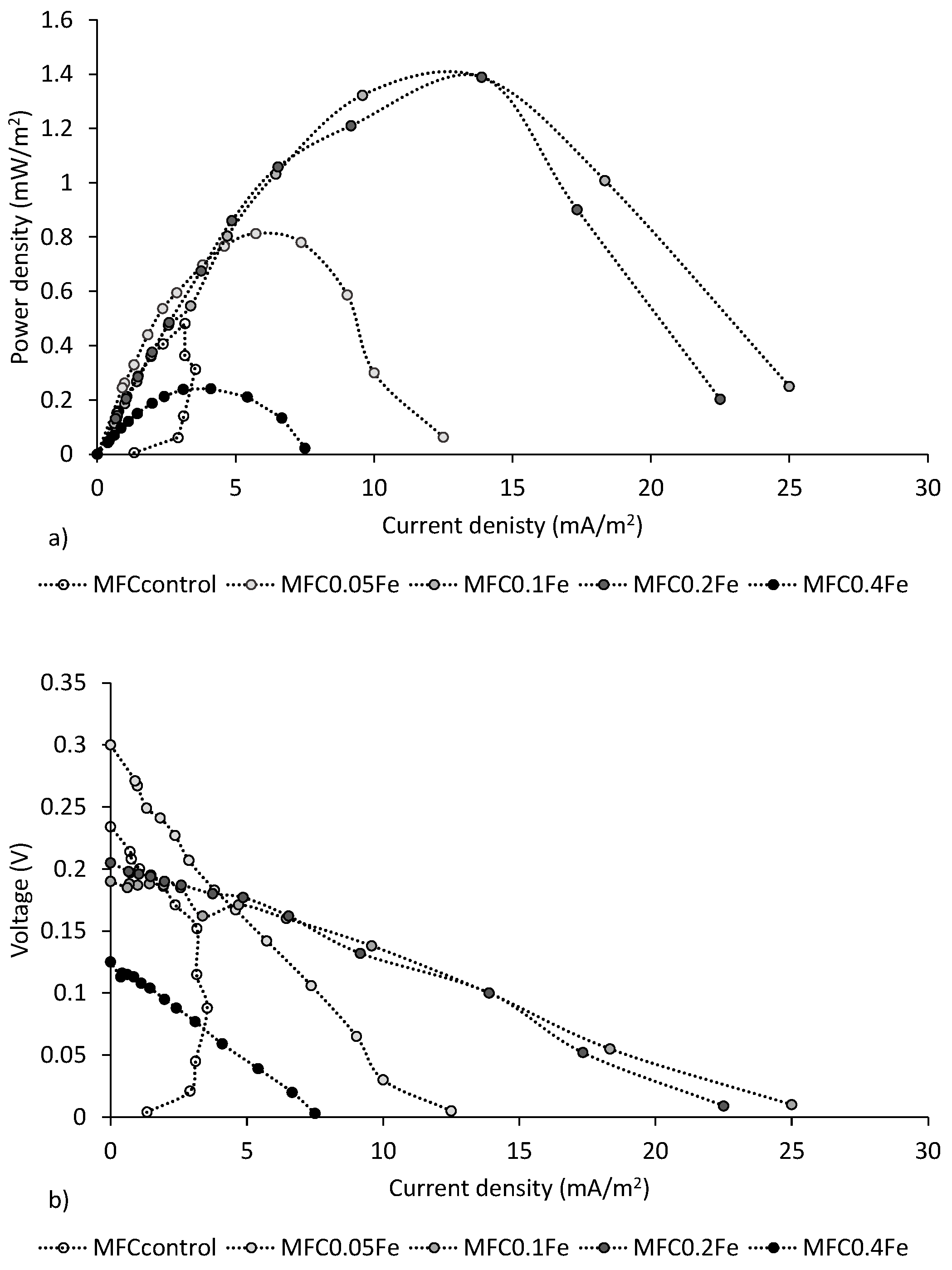
3.2. Electricity Generation
3.3. COD Removal
3.4. Chromatographic Analyses of VFAs
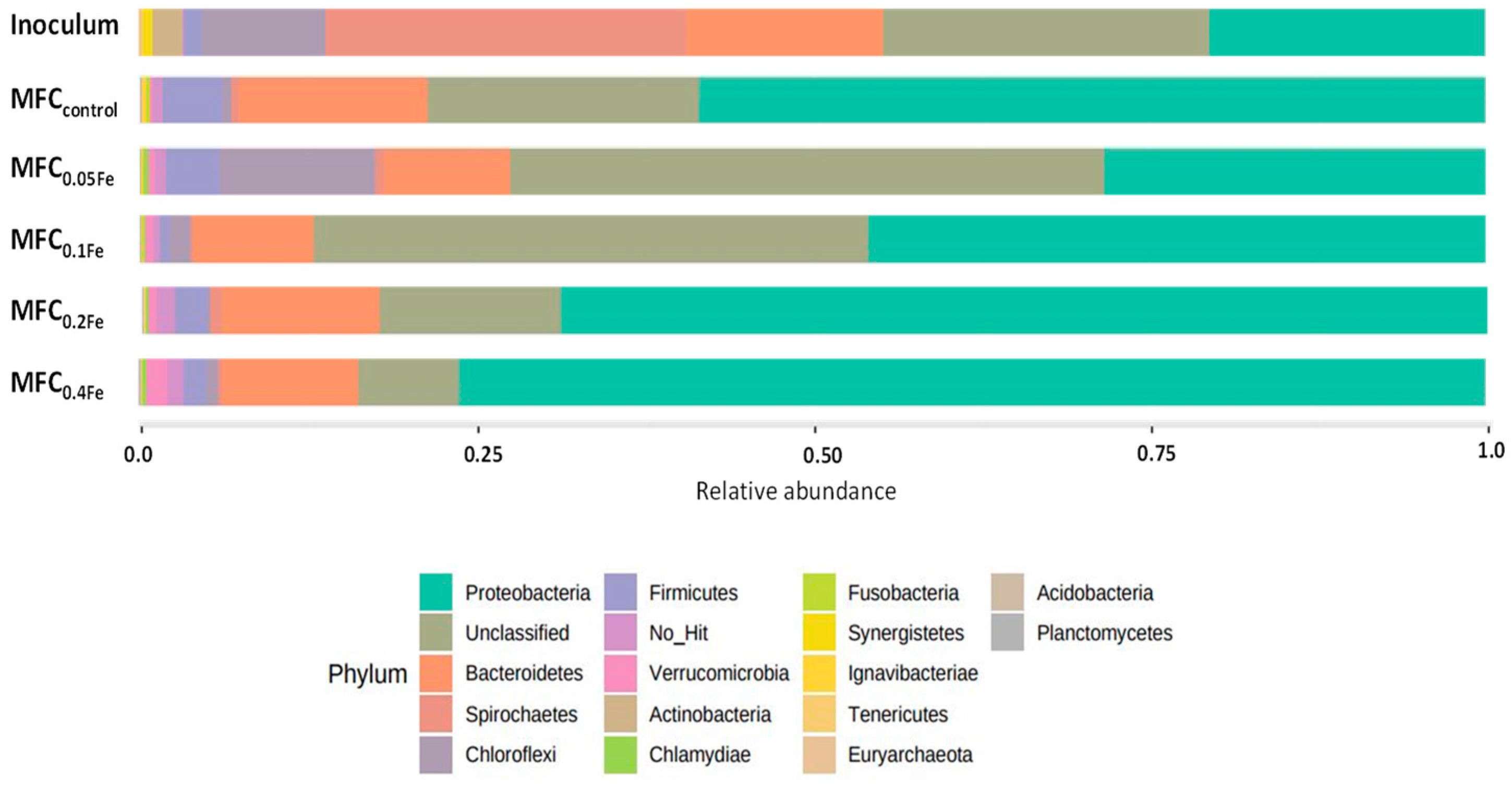
3.5. Microbial Structure of Inoculum and Anode Biofilm in the MFCs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jadhav, D.A.; Mungray, A.K.; Arkatkar, A.; Kumar, S.S. Recent advancement in scaling-up applications of microbial fuel cells: From reality to practicability. Sustain. Energy Technol. Assess. 2021, 45, 101226. [Google Scholar] [CrossRef]
- Mohamed, A.; Zmuda, H.M.; Ha, P.T.; Coats, E.R.; Beyenal, H. Large-scale switchable potentiostatically controlled/microbial fuel cell bioelectrochemical wastewater treatment system. Bioelectrochemistry 2021, 138, 107724. [Google Scholar] [CrossRef] [PubMed]
- Nosek, D.; Jachimowicz, P.; Cydzik-Kwiatkowska, A. Anode modification as an alternative approach to improve electricity generation in microbial fuel cells. Energies 2020, 13, 6596. [Google Scholar] [CrossRef]
- Pandit, S.; Khilari, S.; Roy, S.; Pradhan, D.; Das, D. Improvement of power generation using Shewanella putrefaciens mediated bioanode in a single chambered microbial fuel cell: Effect of different anodic operating conditions. Bioresour. Technol. 2014, 166, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Harshiny, M.; Samsudeen, N.; Kameswara, R.J.; Matheswaran, M. Biosynthesized FeO nanoparticles coated carbon anode for improving the performance of microbial fuel cell. Int. J. Hydrogen Energy 2017, 42, 26488–26495. [Google Scholar] [CrossRef]
- Mohamed, H.O.; Obaid, M.; Poo, K.M.; Abdelkareem, M.A.; Talas, S.A.; Fadali, O.A.; Kim, H.Y.; Chae, K.J. Fe/Fe2O3 nanoparticles as anode catalyst for exclusive power generation and degradation of organic compounds using microbial fuel cell. Chem. Eng. J. 2018, 349, 800–807. [Google Scholar] [CrossRef]
- Gao, X.; Qiu, S.; Lin, Z.; Xie, X.; Yin, W.; Lu, X. Carbon-based composites as anodes for microbial fuel cells: Recent advances and challenges. ChemPlusChem 2021, 86, 1322–1341. [Google Scholar] [CrossRef]
- Beliaev, A.S.; Saffarini, D.A.; McLaughlin, J.L.; Hunnicutt, D. MtrC, an outer membrane decahaem c cytochrome required for metal reduction in Shewanella putrefaciens MR-1. Mol. Microbiol. 2001, 39, 722–730. [Google Scholar] [CrossRef]
- Sharafat, I.; Ali, J.; Hussain, A.; Torres, C.I.; Ali, N. Trivalent iron shaped the microbial community structure to enhance the electrochemical performance of microbial fuel cells inoculated with soil and sediment. J. Environ. Chem. Eng. 2022, 10, 107790. [Google Scholar] [CrossRef]
- Yamashita, T.; Ishida, M.; Asakawa, S.; Kanamori, H.; Sasaki, H.; Ogino, A.; Katayose, Y.; Hatta, T.; Yokoyama, H. Enhanced electrical power generation using flame-oxidized stainless steel anode in microbial fuel cells and the anodic community structure. Biotechnol. Biofuels 2016, 9, 62. [Google Scholar] [CrossRef]
- Xu, X.; Zhao, Q.; Wu, M.; Ding, J.; Zhang, W. Biodegradation of organic matter and anodic microbial communities analysis in sediment microbial fuel cells with/without Fe (III) oxide addition. Bioresour. Technol. 2017, 225, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.H.; Chu, X.Z.; Huang, P.F.; Shan, K. Improved power performance of activated carbon anode by Fe2O3 addition in microbial fuel cells. Appl. Mech. Mater. 2015, 700, 170–174. [Google Scholar] [CrossRef]
- Sayed, E.T.; Sayed, E.T.; Alawadhi, H.; Elsaid, K.; Olabi, A.G.; Adel Almakrani, M.; Bin Tamim, S.T.; Alafranji, G.H.M.; Abdelkareem, M.A. A carbon-cloth anode electroplated with iron nanostructure for microbial fuel cell operated with real wastewater. Sustainability 2020, 12, 6538. [Google Scholar] [CrossRef]
- Sekar, A.D.; Sekar, A.D.; Jayabalan, T.; Muthukumar, H.; Chandrasekaran, N.I.; Mohamed, S.N.; Matheswaran, M. Enhancing power generation and treatment of dairy waste water in microbial fuel cell using Cu-doped iron oxide nanoparticles decorated anode. Energy 2019, 172, 173–180. [Google Scholar] [CrossRef]
- Wang, M.; Wang, M.; Wang, Z.; Hu, F.; Fan, L.; Zhang, X. Polyelectrolytes/α-Fe2O3 modification of carbon cloth anode for dealing with food wastewater in microbial fuel cell. Carbon Resour. Convers. 2020, 3, 76–81. [Google Scholar] [CrossRef]
- Ahmed, M.; Lin, O.; Saup, C.M.; Wilkins, M.J.; Lin, L.S. Effects of Fe/S ratio on the kinetics and microbial ecology of an Fe (III)-dosed anaerobic wastewater treatment system. J. Hazard. Mater. 2019, 369, 593–600. [Google Scholar] [CrossRef]
- Fu, L.; Wang, H.; Huang, Q.; Song, T.; Xie, J. Modification of carbon felt anode with graphene/Fe2O3 composite for enhancing the performance of microbial fuel cell. Bioprocess Biosyst. Eng. 2020, 43, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Qian, G.; Qian, G.; Ye, L.; Li, L.; Hu, X.; Jiang, B.; Zhao, X. Influence of electric field and iron on the denitrification process from nitrogen-rich wastewater in a periodic reversal bio-electrocoagulation system. Bioresour. Technol. 2018, 258, 177–186. [Google Scholar] [CrossRef]
- Kulandaivelu, J.; Kulandaivelu, J.; Gao, J.; Song, Y.; Shrestha, S.; Li, X.; Li, J.; Doederer, K.; Keller, J.; Yuan, Z.; et al. Removal of pharmaceuticals and illicit drugs from wastewater due to ferric dosing in sewers. Environ. Sci. Technol. 2019, 53, 6245–6254. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J. Organic matter mineralization with reduction of ferric iron in anaerobic sediments. Appl. Environ. Microbiol. 1986, 51, 683–689. [Google Scholar] [CrossRef]
- Zheng, X.; Hou, S.; Amanze, C.; Zheng, Z.; Weimin, Z. Enhancing microbial fuel cell performance using anode modified with Fe3O4 nanoparticles. Bioprocess Biosyst. Eng. 2022, 45, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, Q.; Liu, B.; Li, W.; Zhao, X.; Zuo, W.; Xing, D. Impact of ferrous iron on microbial community of the biofilm in microbial fuel cells. Front. Microbiol. 2017, 8, 920. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yang, Y.; Mei, X.; Liu, B.; Chen, C.; Xing, D. Response of the microbial community structure of biofilms to ferric iron in microbial fuel cells. Sci. Total Environ. 2018, 631, 695–701. [Google Scholar] [CrossRef]
- Luo, T.; Xu, Q.; Wei, W.; Sun, J.; Dai, X.; Ni, B.J. Performance and mechanism of Fe3O4 improving biotransformation of waste activated sludge into liquid high-value products. Environ. Sci. Technol. 2022, 56, 3658–3668. [Google Scholar] [CrossRef]
- Gnana Kumar, G.; Sarathi, V.S.; Nahm, K.S. Recent advances and challenges in the anode architecture and their modifications for the applications of microbial fuel cells. Biosens. Bioelectron. 2013, 43, 461–475. [Google Scholar] [CrossRef]
- Yaqoob, A.A.; Ibrahim, M.N.M.; Umar, K. Biomass-derived composite anode electrode: Synthesis, characterizations, and application in microbial fuel cells (MFCs). J. Environ. Chem. Eng. 2021, 9, 106111. [Google Scholar] [CrossRef]
- Gnana Kumar, G.; Kirubaharan, C.J.; Yoo, D.J.; Kim, A.R. Graphene/poly (3, 4-ethylenedioxythiophene)/Fe3O4 nanocomposite–An efficient oxygen reduction catalyst for the continuous electricity production from wastewater treatment microbial fuel cells. Int. J. Hydrogen Energy 2016, 41, 13208–13219. [Google Scholar] [CrossRef]
- Nosek, D.; Cydzik-Kwiatkowska, A. Microbial structure and energy generation in microbial fuel cells powered with waste anaerobic digestate. Energies 2020, 13, 4712. [Google Scholar] [CrossRef]
- Coelho, M.A.Z.; Russo, C.; Araujo, O.Q.F. Optimization of a sequencing batch reactor for biological nitrogen removal. Water Res. 2000, 34, 2809–2817. [Google Scholar] [CrossRef]
- Lobato, J.; Cañizares, P.; Fernández, F.J.; Rodrigo, M.A. An evaluation of aerobic and anaerobic sludges as start-up material for microbial fuel cell systems. New Biotechnol. 2012, 29, 415–420. [Google Scholar] [CrossRef]
- Nosek, D.; Cydzik-Kwiatkowska, A. Anode modification with reduced graphene oxide–iron oxide improves electricity generation in microbial fuel cell. J. Ecol. Eng. 2022, 23, 147–153. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Standard Methods for the Examination of Water and Wastewater, 18th ed.; American Public Health Association (APHA), American Water Works Association (AWWA), Water Pollution Control Federation (WPCF): Washington, DC, USA, 1992. [Google Scholar]
- Bułkowska, K.; Mikucka, W.; Pokój, T. Enhancement of biogas production from cattle manure using glycerine phase as a co-substrate in anaerobic digestion. Fuel 2022, 317, 123456. [Google Scholar] [CrossRef]
- Watson, V.J.; Logan, B.E. Analysis of polarization methods for elimination of power overshoot in microbial fuel cells. Electrochem. Commun. 2011, 13, 54–56. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Nat. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef]
- Świątczak, P.; Cydzik-Kwiatkowska, A. Performance and microbial characteristics of biomass in a full-scale aerobic granular sludge wastewater treatment plant. Environ. Sci. Pollut. Res. 2018, 25, 1655–1669. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst—A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using Microbiome Analyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Waste not, want not: Why rarefying microbiome data is inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef]
- Fan, L.; Xi, Y. Effect of polypyrrole-Fe3O4 composite modified anode and its electrodeposition time on the performance of microbial fuel cells. Energies 2021, 14, 2461. [Google Scholar] [CrossRef]
- O’Brien, F.J.; Harley, B.A.; Yannas, I.V.; Gibson, L.J. The effect of pore size on cell adhesion in collagen-GAG scaffolds. Biomaterials 2005, 26, 433–441. [Google Scholar] [CrossRef]
- Wei, L.; Han, H.; Shen, J. Effects of temperature and ferrous sulfate concentrationson the performance of microbial fuel cell. Int. J. Hydrogen Energy 2013, 38, 11110–11116. [Google Scholar] [CrossRef]
- Kuczyński, M.; Łuba, M.; Mikołajczyk, T.; Pierożyński, B.; Jasiecka-Mikołajczyk, A.; Smoczyński, L.; Sołowiej, P.; Wojtacha, P. Electrodegradation of acid mixture dye through the employment of Cu/Fe macro-corrosion galvanic cell in Na2SO4 synthetic wastewater. Molecules 2021, 26, 4580. [Google Scholar] [CrossRef] [PubMed]
- Pareek, A.; Shanthi Sravan, J.; Venkata Mohan, S. Graphene modified electrodes for bioelectricity generation in mediator-less microbial fuel cell. J. Mater. Sci. 2019, 54, 11604–11617. [Google Scholar] [CrossRef]
- Yang, Q.; Yang, S.; Liu, G.; Zhou, B.; Yu, X.; Yin, Y.; Yang, J.; Zhao, H. Boosting the anode performance of microbial fuel cells with a bacteria-derived biological iron oxide/carbon nanocomposite catalyst. Chemosphere 2021, 268, 128800. [Google Scholar] [CrossRef]
- Ren, Z.; Steinberg, L.M.; Regan, J.M. Electricity production and microbial biofilm characterization in cellulose-fed microbial fuel cells. Water Sci. Technol. 2008, 58, 617–622. [Google Scholar] [CrossRef]
- Paucar, N.E.; Sato, C. Microbial fuel cell for energy production, nutrient removal and recovery from wastewater: A review. Processes 2021, 9, 1318. [Google Scholar] [CrossRef]
- Baby, M.G.; Ahammed, M.M. Nutrient removal and recovery from wastewater by microbial fuel cell-based systems–A review. Water Sci. Technol. 2022, 86, 29–55. [Google Scholar] [CrossRef]
- Huang, S.; Jaffé, P. Characterization of incubation experiments and development of an enrichment culture capable of ammonium oxidation under iron-reducing conditions. Biogeosciences 2015, 12, 769–779. [Google Scholar] [CrossRef]
- Bose, S.; Hochella, M.F., Jr.; Gorby, Y.A.; Kennedy, D.W.; McCready, D.E.; Madden, A.S.; Lower, B.H. Bioreduction of hematite nanoparticles by the dissimilatory iron reducing bacterium Shewanella oneidensis MR-1. Geochim. Cosmochim. Acta 2009, 73, 962–976. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, M.; You, S.; Ma, D.; Zhao, J.; Chen, Z. Effect of Fe3+ on the sludge properties and microbial community structure in a lab-scale A2O process. Sci. Total Environ. 2021, 780, 146505. [Google Scholar] [CrossRef]
- Cao, J.; Xie, Y.; Fang, F.; Hu, H. Influences of Fe3+ on phosphorus removal and microbial products in denitrifying phosphorus removal system. Res. Environ. Sci. 2015, 28, 125–133. [Google Scholar] [CrossRef]
- Mena, N.P.; Bulteau, A.L.; Salazar, J.; Hirsch, E.C.; Núñez, M.T. Effect of mitochondrial complex I inhibition on Fe–S cluster protein activity. Biochem. Biophys. Res. Commun. 2011, 409, 241–246. [Google Scholar] [CrossRef]
- Jadhav, G.S.; Ghangrekar, M.M. Performance of microbial fuel cell subjected to variation in pH, temperature, external load and substrate concentration. Bioresour. Technol. 2009, 100, 717–723. [Google Scholar] [CrossRef]
- Franks, A.E.; Nevin, K.P.; Jia, H.; Izallalen, M.; Woodard, T.L.; Lovley, D.R. Novel strategy for three-dimensional real-time imaging of microbial fuel cell communities: Monitoring the inhibitory effects of proton accumulation within the anode biofilm. Energy Environ. Sci. 2009, 2, 113–119. [Google Scholar] [CrossRef]
- Wu, S.; Zou, S.; Yang, Y.; Qian, G.; He, Z. Enhancing the performance of an osmotic microbial fuel cell through self-buffering with reverse-fluxed sodium bicarbonate. Chem. Eng. J. 2018, 349, 241–248. [Google Scholar] [CrossRef]
- Yu, J.; Park, Y.; Kim, B.; Lee, T. Power densities and microbial communities of brewery wastewater-fed microbial fuel cells according to the initial substrates. Bioprocess Biosyst. Eng. 2015, 38, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Arun, V.; Mino, T.; Matsuo, T. Biological mechanism of acetate uptake mediated by carbohydrate consumption in excess phosphorus removal systems. Water Res. 1988, 22, 565–570. [Google Scholar] [CrossRef]
- Wang, R.; Li, Y.; Chen, W.; Zou, J.; Chen, Y. Phosphate release involving PAOs activity during anaerobic fermentation of EBPR sludge and the extension of ADM1. J. Chem. Eng. 2016, 287, 436–447. [Google Scholar] [CrossRef]
- Barcenilla, A.; Barcenilla, A.; Pryde, S.E.; Martin, J.C.; Duncan, S.H.; Stewart, C.S.; Henderson, C.; Flint, H.J. Phylogenetic relationships of butyrate-producing bacteria from the human gut. Appl. Environ. Microbiol. 2000, 66, 1654–1661. [Google Scholar] [CrossRef]
- Ye, W.; Lu, J.; Ye, J.; Zhou, Y. The effects and mechanisms of zero-valent iron on anaerobic digestion of solid waste: A mini-review. J. Clean. Prod. 2021, 278, 123567. [Google Scholar] [CrossRef]
- Barredo, M.S.; Evison, L. Effect of propionate toxicity on methanogen-enriched sludge, Methanobrevibacter smithii, and Methanospirillum hungatii at different pH values. Appl. Environ. Microbiol. 1991, 57, 1764–1769. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Li, H.; Peng, L.; Qin, Y.; Lin, X.; Zheng, L.; Li, C. Porous α-Fe2O3 nanofiber combined with carbon nanotube as anode to enhance the bioelectricity generation for microbial fuel cell. Electrochim. Acta 2021, 391, 138984. [Google Scholar] [CrossRef]
- Schilirò, T.; Tommasi, T.; Armato, C.; Hidalgo, D.; Traversi, D.; Bocchini, S.; Cilli, C.F.; Pirri, C.F. The study of electrochemically active planktonic microbes in microbial fuel cells in relation to different carbon-based anode materials. Energy 2016, 106, 277–284. [Google Scholar] [CrossRef]
- Luo, J.; Feng, L.; Chen, Y.; Sun, H.; Shen, Q.; Li, X.; Chen, H. Alkyl polyglucose enhancing propionic acid enriched short-chain fatty acids production during anaerobic treatment of waste activated sludge and mechanisms. Water Res. 2015, 73, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Chen, G.Q.; Wu, Y.H.; Chen, Z.; Luo, L.W.; Wang, Y.H.; Tong, X.; Bai, Y.; Wang, H.B.; Xu, Y.Q.; et al. Enhanced extracellular polymeric substances production and aggravated membrane fouling potential caused by different disinfection treatment. J. Membr. Sci. 2022, 642, 120007. [Google Scholar] [CrossRef]
- Lovley, D.R.; Ueki, T.; Zhang, T.; Malvankar, N.; Shrestha, P.M.; Flanagan, K.A.; Aklujkar, M.; Butler, J.E.; Giloteaux, L.; Rotaru, A.E.; et al. Geobacter: The microbe electric’s physiology, ecology, and practical applications. Adv. Microb. Physiol. 2011, 59, 1–100. [Google Scholar] [CrossRef]
- Kiely, P.D.; Call, D.F.; Yates, M.D.; Regan, J.R.; Logan, B.E. Anodic biofilms in microbial fuel cells harbor low numbers of higher- power producing bacteria than abundant genera. Appl. Microbiol. Biotechnol. 2010, 88, 371–380. [Google Scholar] [CrossRef]
- Logan, B.E.; Rossi, R.; Ragab, A.; Saikaly, P.E. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 2019, 17, 307–319. [Google Scholar] [CrossRef]
- Phillips, E.J.; Lovley, D.R.; Roden, E.E. Composition of non-microbially reducible Fe (III) in aquatic sediments. Appl. Environ. Microbiol. 1993, 59, 2727–2729. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.H.R.; Hixson, K.K.; Aklujkar, M.A.; Lipton, M.S.; Smith, R.D.; Lovley, D.R.; Mester, T. Proteome of Geobacter sulfurreducens grown with Fe(III) oxide or Fe(III) citrate as the electron acceptor. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2008, 1784, 1935–1941. [Google Scholar] [CrossRef]
- DiLoreto, Z.A.; Bontognali, T.R.; Al Disi, Z.A.; Al-Kuwari, H.A.S.; Williford, K.H.; Strohmenger, C.J.; Sadooni, F.; Palermo, C.; Rivers, J.M.; McKenzie, J.A.; et al. Microbial community composition and dolomite formation in the hypersaline microbial mats of the Khor Al-Adaid sabkhas, Qatar. Extremophiles 2019, 23, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Hasan, K.; Reddy, K.V.R.; Eßmann, V.; Górecki, K.; Conghaile, P.Ó.; Schuhmann, W.; Leech, D.; Hägerhäll, C.; Gorton, L. Electrochemical communication between electrodes and Rhodobacter capsulatus grown in different metabolic modes. Electroanalysis 2015, 27, 118–127. [Google Scholar] [CrossRef]
- Wong, M.T.; Cheng, D.; Wang, R.; Hsing, I.M. Modifying the endogenous electron fluxes of Rhodobacter sphaeroides 2.4. 1 for improved electricity generation. Enzym. Microb. Technol. 2016, 86, 45–51. [Google Scholar] [CrossRef]
- Zheng, W.; Cai, T.; Huang, M.; Chen, D. Comparison of electrochemical performances and microbial community structures of two photosynthetic microbial fuel cells. J. Biosci. Bioeng. 2017, 124, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Price-Whelan, A.; Dietrich, L.E.; Newman, D.K. Rethinking ‘secondary’ metabolism: Physiological roles for phenazine antibiotics. Nat. Chem. Biol. 2006, 2, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Žalnėravičius, R.; Paškevičius, A.; Samukaitė-Bubnienė, U.; Ramanavičius, S.; Vilkienė, M.; Mockevičienė, I.; Ramanavičius, A. Microbial fuel cell based on nitrogen-fixing Rhizobium anhuiense bacteria. Biosensors 2022, 12, 113. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fu, B.; Xi, J.; Hu, H.Y.; Liang, P.; Huang, X.; Zhang, X. Remediation of simulated malodorous surface water by columnar air-cathode microbial fuel cells. Sci. Total Environ. 2019, 687, 287–296. [Google Scholar] [CrossRef]
- Whaley-Martin, K.; Jessen, G.L.; Nelson, T.C.; Mori, J.F.; Apte, S.; Jarolimek, C.; Warren, L.A. The potential role of Halothiobacillus spp. in sulfur oxidation and acid generation in circum-neutral mine tailings reservoirs. Front. Microbiol. 2019, 10, 297. [Google Scholar] [CrossRef]
- Miettinen, H.; Bomberg, M.; Le, T.M.K.; Kinnunen, P. Identification and metabolism of naturally prevailing microorganisms in zinc and copper mineral processing. Minerals 2021, 11, 156. [Google Scholar] [CrossRef]
- Yamada, T.; Sekiguchi, Y.; Hanada, S.; Imachi, H.; Ohashi, A.; Harada, H.; Kamagata, Y. Anaerolinea thermolimosa sp. nov., Levilinea saccharolytica gen. nov., sp. nov. and Leptolinea tardivitalis gen. nov., sp. nov., novel filamentous anaerobes, and description of the new classes Anaerolineae classis nov. and Caldilineae classis nov. in the bacterial phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 2006, 56, 1331–1340. [Google Scholar] [CrossRef]
- Yang, C.; Xiao, N.; Chang, Z.A.; Huang, J.J.; Zeng, W. Biodegradation of TOC by nano-Fe2O3 modified SMFC and its potential environmental effects. ChemistrySelect 2021, 6, 5597–5602. [Google Scholar] [CrossRef]
- Zhang, D.; Shen, J.; Shi, H.; Su, G.; Jiang, X.; Li, J.; Liu, J.; Mu, Y.; Wang, L. Substantially enhanced anaerobic reduction of nitrobenzene by biochar stabilized sulfide-modified nanoscale zero-valent iron: Process and mechanisms. Environ. Int. 2019, 131, 105020. [Google Scholar] [CrossRef]
- Zhang, K.; Wu, X.; Luo, H.; Li, X.; Chen, W.; Chen, J.; Mo, Y.; Wang, W. CH4 control and associated microbial process from constructed wetland (CW) by microbial fuel cells (MFC). J. Environ. Manag. 2020, 260, 110071. [Google Scholar] [CrossRef] [PubMed]
- Iigatani, R.; Ito, T.; Watanabe, F.; Nagamine, M.; Suzuki, Y.; Inoue, K. Electricity generation from sweet potato-shochu waste using microbial fuel cells. J. Biosci. Bioeng. 2019, 128, 56–63. [Google Scholar] [CrossRef]
- Wang, A.; Liu, L.; Sun, D.; Ren, N.; Lee, D.J. Isolation of Fe(III)-reducing fermentative bacterium Bacteroides sp. W7 in the anode suspension of a microbial electrolysis cell (MEC). Int. J. Hydrogen Energy 2010, 35, 3178–3182. [Google Scholar] [CrossRef]
- Wang, J.; Song, X.; Wang, Y.; Abayneh, B.; Li, Y.; Yan, D.; Bai, J. Nitrate removal and bioenergy production in constructed wetland coupled with microbial fuel cell: Establishment of electrochemically active bacteria community on anode. Bioresour. Technol. 2016, 221, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Kisková, J.; Perháčová, Z.; Vlčko, L.; Sedláková, J.; Kvasnová, S.; Pristaš, P. The bacterial population of neutral mine drainage water of Elizabeth’s shaft (Slovinky, Slovakia). Curr. Microbiol. 2018, 75, 988–996. [Google Scholar] [CrossRef]
- Li, X.; Zhang, W.; Liu, T.; Chen, L.; Chen, P.; Li, F. Changes in the composition and diversity of microbial communities during anaerobic nitrate reduction and Fe(II) oxidation at circumneutral pH in paddy soil. Soil Biol. Biochem. 2016, 94, 70–79. [Google Scholar] [CrossRef]
- Drewniak, L.; Krawczyk, P.S.; Mielnicki, S.; Adamska, D.; Sobczak, A.; Lipinski, L.; Burec-Drewniak, W.; Sklodowska, A. Physiological and metagenomic analyses of microbial mats involved in self-purification of mine waters contaminated with heavy metals. Front. Microbiol. 2016, 7, 1252. [Google Scholar] [CrossRef]
- Li, Z.L.; Zhu, Z.L.; Lin, X.Q.; Chen, F.; Li, X.; Liang, B.; Huang, C.; Zhamg, Y.M.; Sun, K.; Zhou, A.A.; et al. Microbial fuel cell-upflow biofilter coupling system for deep denitrification and power recovery: Efficiencies, bacterial succession and interactions. Environ. Res. 2021, 196, 110331. [Google Scholar] [CrossRef]
- Varanasi, J.L.; Sinha, P.; Das, D. Maximizing power generation from dark fermentation effluents in microbial fuel cell by selective enrichment of exoelectrogens and optimization of anodic operational parameters. Biotechnol. Lett. 2017, 39, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Kuever, J.; Rainey, F.A.; Widdel, F. Desulfobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley Online Library: Hoboken, NJ, USA, 2015; pp. 1–7. [Google Scholar] [CrossRef]
- Nicolova, M.; Groudev, S.; Spasova, I.; Groudeva, V.; Georgiev, P. Electricity generation by means of microorganisms from different physiological groups. Industry 4.0 2018, 3, 80–81. [Google Scholar]
- Tang, R.; Wu, D.; Chen, W.; Feng, C.; Wei, C. Biocathode denitrification of coke wastewater effluent from an industrial aeration tank: Effect of long-term adaptation. Biochem. Eng. J. 2017, 125, 151–160. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, K.; Ouyang, H.; Li, M.K.; Luo, Z.; Li, Y.; Chen, C.; Yang, X.; Shao, Z.; Yan, D.Y. Simultaneous PAHs degradation, odour mitigation and energy harvesting by sediment microbial fuel cell coupled with nitrate-induced biostimulation. J. Environ. Manag. 2021, 284, 112045. [Google Scholar] [CrossRef] [PubMed]
- Sangcharoen, A.; Niyom, W.; Suwannasilp, B.B. A microbial fuel cell treating organic wastewater containing high sulfate under continuous operation: Performance and microbial community. Process Biochem. 2015, 50, 1648–1655. [Google Scholar] [CrossRef]
- Mohite, B.V.; Koli, S.H.; Patil, S.V. Heavy metal stress and its consequences on exopolysaccharide (EPS)-producing Pantoea agglomerans. Appl. Biochem. Biotechnol. 2018, 186, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, I.M.; Konnova, S.A.; Sigida, E.N.; Lyubun, E.V.; Muratova, A.Y.; Fedonenko, Y.P.; Elbanna, K. Bioremediation potential of a halophilic Halobacillus sp. strain, EG1HP4QL: Exopolysaccharide production, crude oil degradation, and heavy metal tolerance. Extremophiles 2020, 24, 157–166. [Google Scholar] [CrossRef]
- Giese, E.C. Evidences of EPS-iron (III) ions interactions on bioleaching process mini-review: The key to improve performance. Orbital-Electron. J. Chem. 2019, 11, 200–204. [Google Scholar] [CrossRef]
- Sag, Y.; Kutsal, T. Determination of the biosorption heats of heavy metal ions on Zoogloea ramigera and Rhizopus arrhizus. Biochem. Eng. J. 2000, 6, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Perpetuo, E.A.; Souza, C.B.; Nascimento, C.A.O. Engineering Bacteria for Bioremediation. Prog. Mol. Biol. Transl. Sci. 2011, 28, 605–632. [Google Scholar] [CrossRef]
- Mei, X.; Guo, C.; Liu, B.; Tang, Y.; Xing, D. Shaping of bacterial community structure in microbial fuel cells by different inocula. RSC Adv. 2015, 5, 78136–78141. [Google Scholar] [CrossRef]
- Lee, J.; Phung, N.T.; Chang, I.S.; Kim, B.H.; Sung, H.C. Use of acetate for enrichment of electrochemically active microorganisms and their 16S rDNA analyses. FEMS Microbiol. Lett. 2003, 223, 185–191. [Google Scholar] [CrossRef]
- Alatraktchi, F.A.A.; Zhang, Y.; Angelidaki, I. Nanomodification of the electrodes in microbial fuel cell: Impact of nanoparticle density on electricity production and microbial community. Appl. Energy 2014, 116, 216–222. [Google Scholar] [CrossRef]
- Kalyuzhnaya, M.G.; De Marco, P.; Bowerman, S.; Pacheco, C.C.; Lara, J.C.; Lidstrom, M.E.; Chistoserdova, L. Methyloversatilis universalis gen. nov., sp. nov., a novel taxon within the Betaproteobacteria represented by three methylotrophic isolates. Int. J. Syst. Evol. 2006, 56, 2517–2522. [Google Scholar] [CrossRef]
- Ramanaiah, S.V.; Cordas, C.M.; Matias, S.C.; Reddy, M.V.; Leitao, J.H.; Fonseca, L.P. Bioelectricity generation using long-term operated biocathode: RFLP based microbial diversity analysis. Biotechnol. Rep. 2021, 32, e00693. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Weitz, A.C.; Paris, J.; Tang, Y.; Zhang, J.; Song, H.; Naowarojna, N.; Li, K.; Qiao, L.; Lopez, J.; et al. OvoAMtht from Methyloversatilis thermotolerans ovothiol biosynthesis is a bifunction enzyme: Cysteine dioxygenase and sulfoxide synthase activities. Chem. Sci. 2022, 13, 3589–3598. [Google Scholar] [CrossRef] [PubMed]
- Issayeva, A.U.; Pankiewicz, R.; Otarbekova, A.A. Bioleaching of metals from wastes of phosphoric fertilizers production. Pol. J. Environ. Stud. 2020, 29, 4101–4108. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Wang, Y.L.; Li, W.H.; Bao, L.N.; Wang, L.H.; Huang, X.H.; Huang, B. Biogas emission from an anaerobic reactor. Aerosol Air. Qual. Res. 2018, 18, 1493–1502. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reactor—Cycle Number | OTU | Chao1 | Shannon | ACE | Total Read Counts |
|---|---|---|---|---|---|
| Inoculum | 91 | 91.5 | 2.51 | 92.7 | 26,877 |
| MFCcontrol—3 | 103 | 103.6 | 3.14 | 103.6 | 25,053 |
| MFCcontrol—10 | 108 | 108.5 | 2.97 | 108.2 | 29,183 |
| MFCcontrol—18 | 112 | 114.3 | 2.85 | 114.8 | 28,548 |
| MFC0.05Fe—3 | 108 | 108.5 | 2.85 | 108.3 | 35,811 |
| MFC0.05Fe—10 | 105 | 108.0 | 1.51 | 108.1 | 38,831 |
| MFC0.05Fe—18 | 111 | 111.0 | 2.77 | 111.3 | 24,160 |
| MFC0.1Fe—3 | 109 | 110.6 | 1.47 | 111.1 | 43,390 |
| MFC0.1Fe—10 | 101 | 101.6 | 1.23 | 102.8 | 49,705 |
| MFC0.1Fe—18 | 117 | 117.9 | 2.69 | 118.5 | 33,249 |
| MFC0.2Fe—3 | 66 | 69.0 | 1.72 | 68.5 | 24,281 |
| MFC0.2Fe—10 | 101 | 101.5 | 3.14 | 103.5 | 28,223 |
| MFC0.2Fe—18 | 118 | 122.7 | 2.57 | 122.5 | 30,545 |
| MFC0.4Fe—3 | 64 | 65.3 | 1.45 | 66.8 | 28,970 |
| MFC0.4Fe—10 | 103 | 104.1 | 3.21 | 105.3 | 17,967 |
| MFC0.4Fe—18 | 111 | 111.8 | 2.56 | 113.0 | 34,113 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nosek, D.; Mikołajczyk, T.; Cydzik-Kwiatkowska, A. Anode Modification with Fe2O3 Affects the Anode Microbiome and Improves Energy Generation in Microbial Fuel Cells Powered by Wastewater. Int. J. Environ. Res. Public Health 2023, 20, 2580. https://doi.org/10.3390/ijerph20032580
Nosek D, Mikołajczyk T, Cydzik-Kwiatkowska A. Anode Modification with Fe2O3 Affects the Anode Microbiome and Improves Energy Generation in Microbial Fuel Cells Powered by Wastewater. International Journal of Environmental Research and Public Health. 2023; 20(3):2580. https://doi.org/10.3390/ijerph20032580
Chicago/Turabian StyleNosek, Dawid, Tomasz Mikołajczyk, and Agnieszka Cydzik-Kwiatkowska. 2023. "Anode Modification with Fe2O3 Affects the Anode Microbiome and Improves Energy Generation in Microbial Fuel Cells Powered by Wastewater" International Journal of Environmental Research and Public Health 20, no. 3: 2580. https://doi.org/10.3390/ijerph20032580
APA StyleNosek, D., Mikołajczyk, T., & Cydzik-Kwiatkowska, A. (2023). Anode Modification with Fe2O3 Affects the Anode Microbiome and Improves Energy Generation in Microbial Fuel Cells Powered by Wastewater. International Journal of Environmental Research and Public Health, 20(3), 2580. https://doi.org/10.3390/ijerph20032580