
Urban Green Spaces and Vector-Borne Disease Risk in Africa: The Case of an Unclean Forested Park in Libreville (Gabon, Central Africa)

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Larval Sampling
2.3. Adult Mosquito Collection
2.4. Data Analysis
3. Results
3.1. Typology and Positivity of Larval Habitats Inside and Outside the Forest
3.2. Mosquito Species Composition and Diversity
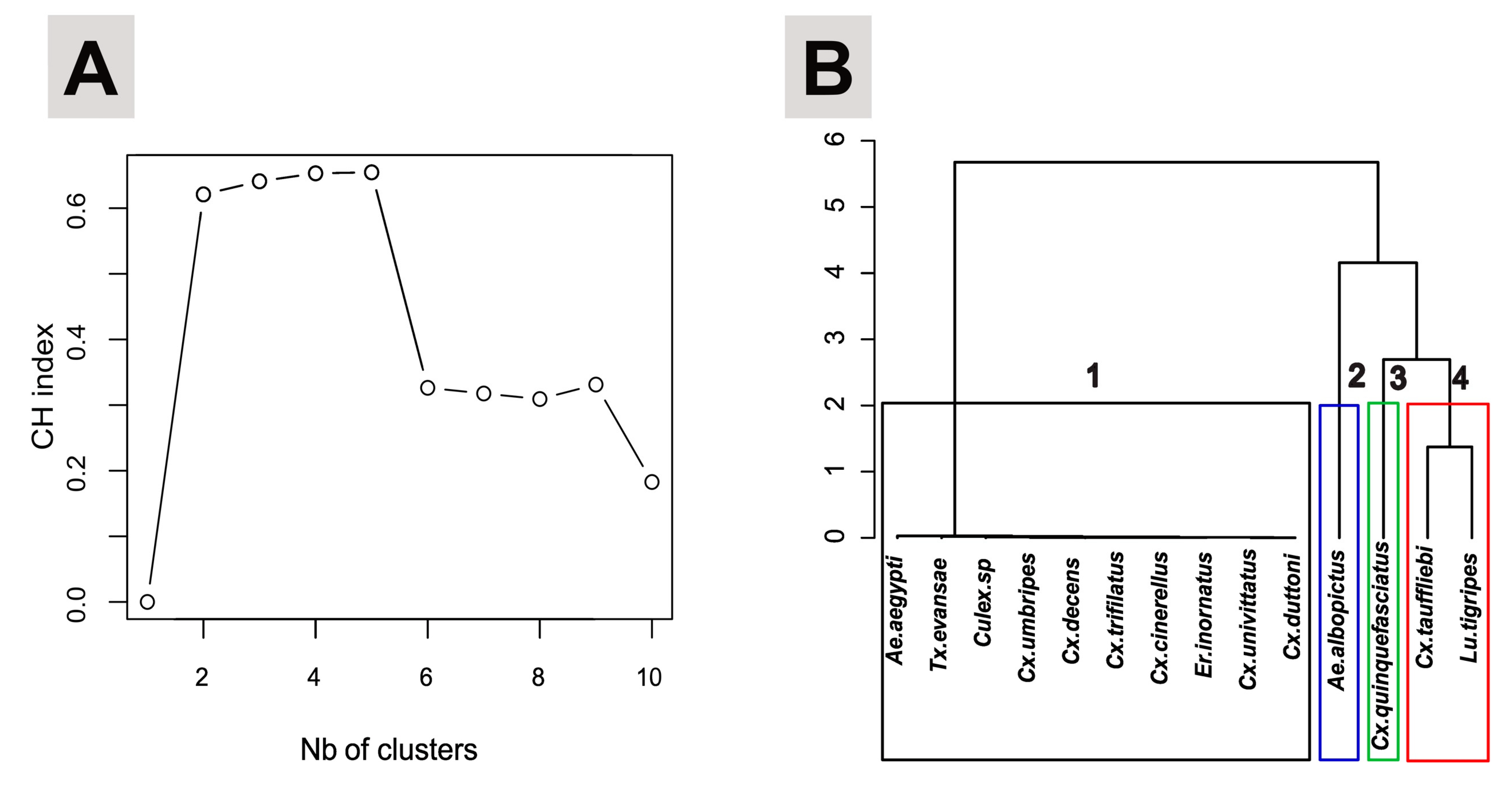
3.3. Larval Habitat Typology, Similarity and Species Clustering
3.4. Biting Patterns of Mosquito Species
4. Discussion
4.1. Mosquito Communities in the Urban Forested Area of Sibang
4.2. The Mosquito Proliferation Drivers in the Urban Forested Area of Sibang
4.3. Mosquito Aggressiveness in the Urban Forested Area of Sibang
4.4. Mosquito Aggressiveness and Urban Greening
4.5. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brady, O.J.; Hay, S.I. The global expansion of dengue: How Aedes aegypti mosquitoes enabled the first pandemic arbovirus. Annu. Rev. Entomol. 2020, 65, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Hoerauf, A.; Pfarr, K.; Mand, S.; Debrah, A.; Specht, S. Filariasis in Africa—Treatment challenges and prospects. Clin. Microbiol. Infect. 2011, 17, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Rebollo, M.P.; Bockarie, M.J. Can lymphatic filariasis be eliminated by 2020? Trends Parasitol. 2017, 33, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, chikungunya, and other emerging vector-borne viral diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Malaria Report; WHO: Geneva, Switzerland, 2021; pp. 2–5. [Google Scholar]
- Eder, M.; Cortes, F.; Teixeira de Siqueira Filha, N.; Araújo de França, G.V.; Degroote, S.; Braga, C.; Ridde, V.; Turchi Martelli, C.M. Scoping review on vector-borne diseases in urban areas: Transmission dynamics, vectorial capacity and co-infection. Infect. Dis. Poverty 2018, 7, 90. [Google Scholar] [CrossRef] [PubMed]
- Koukouikila-Koussounda, F.; Ntoumi, F. Malaria epidemiological research in the Republic of Congo. Malar. J. 2016, 15, 598. [Google Scholar] [CrossRef]
- Mbohou, C.N.; Foko, L.P.K.; Nyabeyeu, H.N.; Tonga, C.; Nono, L.K.; Kangam, L.; Bunda, G.W.; Mbou, I.M.; Ngo Hondt, E.O.; Mbe, A.J.K. Malaria screening at the workplace in Cameroon. PLoS ONE 2019, 14, e0225219. [Google Scholar] [CrossRef]
- Messina, J.P.; Taylor, S.M.; Meshnick, S.R.; Linke, A.M.; Tshefu, A.K.; Atua, B.; Mwandagalirwa, K.; Emch, M. Population, behavioural and environmental drivers of malaria prevalence in the Democratic Republic of Congo. Malar. J. 2011, 10, 161. [Google Scholar] [CrossRef]
- Olivier, B.M.B.; Eliezer, M.P.; Ngatimo, E.V.; Pierrette, B.K.E.; Tsalefac, M.; Chrysostome, G.J. Influence of climate variability on the dynamics of malaria transmission among children in Bangui, health challenges in central African Republic. Open J. Pediatr. 2022, 12, 461–475. [Google Scholar] [CrossRef]
- Tewara, M.A.; Mbah-Fongkimeh, P.N.; Dayimu, A.; Kang, F.; Xue, F. Small-area spatial statistical analysis of malaria clusters and hotspots in Cameroon; 2000–2015. BMC Infect. Dis. 2018, 18, 636. [Google Scholar] [CrossRef]
- Leroy, E.M.; Nkoghe, D.; Ollomo, B.; Nze-Nkogue, C.; Becquart, P.; Grard, G.; Pourrut, X.; Charrel, R.; Moureau, G.; Ndjoyi-Mbiguino, A. Concurrent chikungunya and dengue virus infections during simultaneous outbreaks, Gabon, 2007. Emerg. Infect. Dis. 2009, 15, 591. [Google Scholar] [CrossRef] [PubMed]
- Nana-Ndjangwo, S.M.; Djiappi-Tchamen, B.; Mony, R.; Demanou, M.; Keumezeu-Tsafack, J.; Bamou, R.; Awono-Ambene, P.; Bilong Bilong, C.F.; Antonio-Nkondjio, C. Assessment of Dengue and Chikungunya Infections among Febrile Patients Visiting Four Healthcare Centres in Yaoundé and Dizangué, Cameroon. Viruses 2022, 14, 2127. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, Y.; Abe, H.; Mbadinga, M.J.; Ondo, G.N.; Bikangui, R.; Agnandji, S.T.; Lell, B.; Yasuda, J. Re-emergence of dengue, chikungunya, and Zika viruses in 2021 after a 10-year gap in Gabon. IJID Reg. 2022, 5, 68–71. [Google Scholar] [CrossRef]
- Vairo, F.; Aimè Coussoud-Mavoungou, M.P.; Ntoumi, F.; Castilletti, C.; Kitembo, L.; Haider, N.; Carletti, F.; Colavita, F.; Gruber, C.E.; Iannetta, M. Chikungunya outbreak in the Republic of the Congo, 2019—Epidemiological, virological and entomological findings of a South-North Multidisciplinary Taskforce Investigation. Viruses 2020, 12, 1020. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Clark, G.G. Community involvement in the control of Aedes aegypti. Acta Trop. 1996, 61, 169–179. [Google Scholar] [CrossRef]
- Lindsay, S.W.; Wilson, A.; Golding, N.; Scott, T.W.; Takken, W. Improving the built environment in urban areas to control Aedes aegypti-borne diseases. Bull. World Health Organ. 2017, 95, 607. [Google Scholar] [CrossRef]
- Mathey, J.; Rößler, S.; Lehmann, I.; Bräuer, A. Urban green spaces: Potentials and constraints for urban adaptation to climate change. In Resilient Cities; Springer: Dordrecht, Germany, 2011; pp. 479–485. [Google Scholar]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- Bergero, P.E.; Ruggerio, C.A.; Lombardo Berchesi, R.J.; Schweigmann, N.J.; Solari, H.G. Dispersal of Aedes aegypti: Field study in temperate areas using a novel method. J. Vector Borne Dis. 2013, 50, 163–170. [Google Scholar]
- Hayden, M.H.; Uejio, C.K.; Walker, K.; Ramberg, F.; Moreno, R.; Rosales, C.; Gameros, M.; Mearns, L.O.; Zielinski-Gutierrez, E.; Janes, C.R. Microclimate and human factors in the divergent ecology of Aedes aegypti along the Arizona, US/Sonora, MX border. EcoHealth 2010, 7, 64–77. [Google Scholar] [CrossRef]
- Hendy, A.; Hernandez-Acosta, E.; Chaves, B.A.; Fé, N.F.; Valério, D.; Mendonça, C.; de Lacerda, M.V.G.; Buenemann, M.; Vasilakis, N.; Hanley, K.A. Into the woods: Changes in mosquito community composition and presence of key vectors at increasing distances from the urban edge in urban forest parks in Manaus, Brazil. Acta Trop. 2020, 206, 105441. [Google Scholar] [CrossRef]
- Chevalier, J.F.; Nguema Maganga, V.; Assoumou, S. Les Forêt du Gabon en 2008 [Internet]. MEFEPPN; 2008; pp. 61–73; Report No.: 3. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwjphuTlv7j8AhVEgP0HHdyKDFwQFnoECAkQAw&url=https%3A%2F%2Fwww.observatoire-comifac.net%2Ffile%2FeyJtb2RlbCI6IkFwcFxcTW9kZWxzXFxDYXRhbG9ndWVcXE1vZHVsZXNcXEZpbGUiLCJmaWVsZCI6ImRvY3VtZW50X2ZpbGUiLCJpZCI6NjU4fQ&usg=AOvVaw1fNKVWC7RI-UT7ZhkRMauD (accessed on 8 January 2023).
- Grard, G.; Caron, M.; Mombo, I.M.; Nkoghe, D.; Ondo, S.M.; Jiolle, D.; Fontenille, D.; Paupy, C.; Leroy, E.M. Zika virus in Gabon (Central Africa)—2007: A new threat from Aedes albopictus? PLoS Negl. Trop. Dis. 2014, 8, e2681. [Google Scholar] [CrossRef] [PubMed]
- M’bondoukwé, N.P.; Kendjo, E.; Mawili-Mboumba, D.P.; Koumba Lengongo, J.V.; Offouga Mbouoronde, C.; Nkoghe, D.; Touré, F.; Bouyou-Akotet, M.K. Prevalence of and risk factors for malaria, filariasis, and intestinal parasites as single infections or co-infections in different settlements of Gabon, Central Africa. Infect. Dis. Poverty 2018, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, Y.; Abe, H.; Nguema Ondo, G.; Bikangui, R.; Massinga Loembé, M.; Zadeh, V.R.; Essimengane, J.G.; Mbouna, A.V.; Bache, E.B.; Agnandji, S.T. Surveillance of the major pathogenic arboviruses of public health concern in Gabon, Central Africa: Increased risk of West Nile virus and dengue virus infections. BMC Infect. Dis. 2021, 21, 265. [Google Scholar] [CrossRef]
- Mayaux, P.; Bartholomé, E.; Fritz, S.; Belward, A. A new land-cover map of Africa for the year 2000. J. Biogeogr. 2004, 31, 861–877. [Google Scholar] [CrossRef]
- Cordier, S. Sibang, l’histoire d’un arboretum en Afrique. In Nîmes Lett. L’OCIM; 2000; pp. 19–27. Available online: https://doc.ocim.fr/LO/LO072/LO.72(3)-pp.19-27.pdf (accessed on 1 September 2020).
- Edwards, F.W. Mosquitoes of the Ethiopian Region. III.-Culicine Adults and Pupae; Adlard and Sons Limited: London/Dorking, UK, 1941; p. 499. Available online: https://mosquito-taxonomic-inventory.myspecies.info/sites/mosquito-taxonomic-inventory.info/files/Edwards%201941.pdf (accessed on 1 September 2020).
- Huang, Y.-M. The subgenus Stegomyia of Aedes in the Afrotropical Region with keys to the species (Diptera: Culicidae). Zootaxa 2004, 700, 1–120. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The mathematical theory of information. Urbana Univ. Ill. Press 1949, 97, 128–164. [Google Scholar]
- Magurran, A.E. Measuring biological diversity. Curr. Biol. 2021, 31, R1174–R1177. [Google Scholar] [CrossRef]
- Hennig, C. fpc: Flexible Procedures for Clustering (Version 2.2-9). 2020. Available online: https://cran.r-project.org/web/packages/fpc/fpc.pdf (accessed on 1 September 2020).
- Caliński, T.; Harabasz, J. A dendrite method for cluster analysis. Commun. Stat.-Theory Methods 1974, 3, 1–27. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Koumba, A.A.; Koumba, C.R.Z.; Nguema, R.M.; Djogbenou, L.S.; Ondo, P.O.; Ketoh, G.K.; Comlan, P.; M’Batchi, B.; Mavoungou, J.F. Distribution spatiale et saisonnière des gîtes larvaires des moustiques dans les espaces agricoles de la zone de Mouila, Gabon. Int. J. Biol. Chem. Sci. 2018, 12, 1754–1769. [Google Scholar] [CrossRef]
- Paupy, C.; Ollomo, B.; Kamgang, B.; Moutailler, S.; Rousset, D.; Demanou, M.; Hervé, J.-P.; Leroy, E.; Simard, F. Comparative role of Aedes albopictus and Aedes aegypti in the emergence of Dengue and Chikungunya in central Africa. Vector-Borne Zoonotic Dis. 2010, 10, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Sevidzem, S.L.; Pamba, R.; Koumba, A.A.; Zinga-Koumba, C.R.; Mbouloungou, A.; Yacka, L.L.; Djogbenou, L.S.; Mavoungou, J.F.; M’Batchi, B. Typology of breeding sites and species diversity of culicids (Diptera: Culicidae) in Akanda and its environs (North West, Gabon). Eur. J. Biol. Biotechnol. 2020, 1. [Google Scholar] [CrossRef]
- Djoufounna, J.; Mayi, M.P.A.; Bamou, R.; Ningahi, L.G.; Magatsing, F.O.; Djiappi-Tchamen, B.; Djamouko-Djonkam, L.; Antonio-Nkondjio, C.; Tchuinkam, T. Larval habitats characterization and population dynamics of Culex mosquitoes in two localities of the Menoua Division, Dschang and Santchou, West Cameroon. J. Basic Appl. Zool. 2022, 83, 30. [Google Scholar] [CrossRef]
- Kamgang, B.; Happi, J.Y.; Boisier, P.; Njiokou, F.; Hervé, J.; Simard, F.; Paupy, C. Geographic and ecological distribution of the dengue and chikungunya virus vectors Aedes aegypti and Aedes albopictus in three major Cameroonian towns. Med. Vet. Entomol. 2010, 24, 132–141. [Google Scholar] [CrossRef]
- Ngoagouni, C.; Kamgang, B.; Nakouné, E.; Paupy, C.; Kazanji, M. Invasion of Aedes albopictus (Diptera: Culicidae) into central Africa: What consequences for emerging diseases? Parasit. Vectors 2015, 8, 191. [Google Scholar] [CrossRef]
- Chen, C.; Nazni, W.; Lee, H.; Seleena, B.; Mohd Masri, S.; Chiang, Y.; Sofian-Azirun, M. Mixed breeding of Aedes aegypti (L.) and Aedes albopictus Skuse in four dengue endemic areas in Kuala Lumpur and Selangor, Malaysia. Trop. Biomed. 2006, 23, 224–227. [Google Scholar]
- SNR, S.; Norma-Rashid, Y.; Sofian-Azirun, M. Mosquitoes larval breeding habitat in urban and suburban areas, Peninsular Malaysia. Int. J. Bioeng. Life Sci. 2011, 5, 599–603. [Google Scholar]
- Ferreira-de-Lima, V.H.; Câmara, D.C.P.; Honorio, N.A.; Lima-Camara, T.N. The Asian tiger mosquito in Brazil: Observations on biology and ecological interactions since its first detection in 1986. Acta Trop. 2020, 205, 105386. [Google Scholar] [CrossRef]
- Pereira, E.d.S.; Ferreira, R.L.; Hamada, N.; Lichtwardt, R.W. Trichomycete fungi (Zygomycota) associated with mosquito larvae (Diptera: Culicidae) in natural and artificial habitats in Manaus, AM Brazil. Neotrop. Entomol. 2005, 34, 325–329. [Google Scholar] [CrossRef]
- Wermelinger, E.D.; Carvalho, R.W. de Methods and procedures used in Aedes aegypti control in the successful campaign for yellow fever prophylaxis in Rio de Janeiro, Brazil, in 1928 and 1929. Epidemiol. Serviços Saúde 2016, 25, 837–844. [Google Scholar] [CrossRef]
- Appawu, M.; Quartey, S. Effect of temperature on the development and predatory behaviour of Culex (Lutzia) tigripes (Grandpre and Charmoy). Int. J. Trop. Insect Sci. 2000, 20, 129–134. [Google Scholar] [CrossRef]
- Appawu, M.A.; Dadzie, S.K.; Quartey, S.Q. Studies on the feeding behaviour of larvae of the predaceous mosquito Culex (Lutzia) tigripes Grandpre and Chamoy (Diptera: Culicidae). Int. J. Trop. Insect Sci. 2000, 20, 245–250. [Google Scholar] [CrossRef]
- Jackson, N. Observations on the Feeding Habits of a Predaoeous Mosquito Larva, Culex (Lutzia) tigripes Grandpré and Charmoy (Diptera). Physiol. Entomol. 1953, 28, 153–159. [Google Scholar]
- Moirangthem, B.D.; Singh, S.N.; Singh, D.C. Lutzia tigripes (Diptera: Culicidae, Metalutzia) for the mosquito larval control: A new prospect of mosquito control. Intl. J. Mosq. Res. 2018, 5, 1–4. [Google Scholar]
- Longo-Pendy, N.M.; Tene-Fossog, B.; Tawedi, R.E.; Akone-Ella, O.; Toty, C.; Rahola, N.; Braun, J.-J.; Berthet, N.; Kengne, P.; Costantini, C. Ecological plasticity to ions concentration determines genetic response and dominance of Anopheles coluzzii larvae in urban coastal habitats of Central Africa. Sci. Rep. 2021, 11, 15781. [Google Scholar] [CrossRef] [PubMed]
- Mutuku, F.M.; Alaii, J.A.; Bayoh, M.N.; Gimnig, J.E.; Vulule, J.M.; Walker, E.D.; Kabiru, E.; Hawley, W.A. Distribution, Description, and Local Knowledge of Larval Habitats of Anopheles Gambiae sl in a Village in Western Kenya. Am. J. Trop. Med. Hyg. 2006, 74, 44–53. [Google Scholar] [CrossRef]
- Simard, F.; Nchoutpouen, E.; Toto, J.C.; Fontenille, D. Geographic distribution and breeding site preference of Aedes albopictus and Aedes aegypti (Diptera: Culicidae) in Cameroon, Central Africa. J. Med. Entomol. 2005, 42, 726–731. [Google Scholar] [CrossRef]
- Tene Fossog, B.; Ayala, D.; Acevedo, P.; Kengne, P.; Ngomo Abeso Mebuy, I.; Makanga, B.; Magnus, J.; Awono-Ambene, P.; Njiokou, F.; Pombi, M. Habitat segregation and ecological character displacement in cryptic African malaria mosquitoes. Evol. Appl. 2015, 8, 326–345. [Google Scholar] [CrossRef]
- Mnzava, A.; Monroe, A.C.; Okumu, F. Anopheles stephensi in Africa requires a more integrated response. Malar. J. 2022, 21, 156. [Google Scholar] [CrossRef]
- Diallo, D.; Diagne, C.T.; Buenemann, M.; Ba, Y.; Dia, I.; Faye, O.; Sall, A.A.; Faye, O.; Watts, D.M.; Weaver, S.C. Biodiversity pattern of mosquitoes in southeastern Senegal, epidemiological implication in arbovirus and malaria transmission. J. Med. Entomol. 2019, 56, 453–463. [Google Scholar] [CrossRef]
- Pereira dos Santos, T.; Roiz, D.; Santos de Abreu, F.V.; Luz, S.L.B.; Santalucia, M.; Jiolle, D.; Santos Neves, M.S.A.; Simard, F.; Lourenço-de-Oliveira, R.; Paupy, C. Potential of Aedes albopictus as a bridge vector for enzootic pathogens at the urban-forest interface in Brazil. Emerg. Microbes Infect. 2018, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kamgang, B.; Nchoutpouen, E.; Simard, F.; Paupy, C. Notes on the blood-feeding behavior of Aedes albopictus (Diptera: Culicidae) in Cameroon. Parasit. Vectors 2012, 5, 57. [Google Scholar] [CrossRef]
- Delatte, H.; Desvars, A.; Bouétard, A.; Bord, S.; Gimonneau, G.; Vourc’h, G.; Fontenille, D. Blood-feeding behavior of Aedes albopictus, a vector of Chikungunya on La Réunion. Vector-Borne Zoonotic Dis. 2010, 10, 249–258. [Google Scholar] [CrossRef]
- Fontenille, D.; Powell, J.R. From anonymous to public enemy: How does a mosquito become a feared arbovirus vector? Pathogens 2020, 9, 265. [Google Scholar] [CrossRef]
- Pagès, F.; Peyrefitte, C.N.; Mve, M.T.; Jarjaval, F.; Brisse, S.; Iteman, I.; Gravier, P.; Nkoghe, D.; Grandadam, M. Aedes albopictus mosquito: The main vector of the 2007 Chikungunya outbreak in Gabon. PLoS ONE 2009, 4, e4691. [Google Scholar] [CrossRef]
- Paupy, C.; Kassa Kassa, F.; Caron, M.; Nkoghé, D.; Leroy, E.M. A chikungunya outbreak associated with the vector Aedes albopictus in remote villages of Gabon. Vector-Borne Zoonotic Dis. 2012, 12, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Laganier, R.; Nangouma, A. First record of Ae. albopictus (Skuse 1894), in Central African Republic. Trop. Med. Int. Health 2010, 15, 1185–1189. [Google Scholar] [CrossRef]
- Kamgang, B.; Ngoagouni, C.; Manirakiza, A.; Nakouné, E.; Paupy, C.; Kazanji, M. Temporal patterns of abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) and mitochondrial DNA analysis of Ae. albopictus in the Central African Republic. PLoS Negl. Trop. Dis. 2013, 7, e2590. [Google Scholar] [CrossRef] [PubMed]
- Bagny Beilhe, L.; Arnoux, S.; Delatte, H.; Lajoie, G.; Fontenille, D. Spread of invasive Aedes albopictus and decline of resident Aedes aegypti in urban areas of Mayotte 2007–2010. Biol. Invasions 2012, 14, 1623–1633. [Google Scholar] [CrossRef]
- O’meara, G.F.; Evans Jr, L.F.; Gettman, A.D.; Cuda, J.P. Spread of Aedes albopictus and decline of Ae. aegypti (Diptera: Culicidae) in Florida. J. Med. Entomol. 1995, 32, 554–562. [Google Scholar] [CrossRef]
- McLean, R.G.; Ubico, S.R. Arboviruses in birds. Infect. Dis. Wild Birds 2007, 17, 62. [Google Scholar]
- Araujo, R.V.; Albertini, M.R.; Costa-da-Silva, A.L.; Suesdek, L.; Franceschi, N.C.S.; Bastos, N.M.; Katz, G.; Cardoso, V.A.; Castro, B.C.; Capurro, M.L. São Paulo urban heat islands have a higher incidence of dengue than other urban areas. Braz. J. Infect. Dis. 2015, 19, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Balany, F.; Ng, A.W.; Muttil, N.; Muthukumaran, S.; Wong, M.S. Green infrastructure as an urban heat island mitigation strategy—A review. Water 2020, 12, 3577. [Google Scholar] [CrossRef]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Temperature-dependent effects on the replication and transmission of arthropod-borne viruses in their insect hosts. Curr. Opin. Insect Sci. 2016, 16, 108–113. [Google Scholar] [CrossRef]
- Chan, M.; Johansson, M.A. The incubation periods of dengue viruses. PLoS ONE 2012, 7, e50972. [Google Scholar] [CrossRef]
- Coffinet, T.; Mourou, J.; Pradines, B.; Toto, J.; Jarjaval, F.; Amalvict, R.; Kombila, M.; Carnevale, P.; Pages, F. First record of Aedes albopictus in Gabon. J. Am. Mosq. Control Assoc. 2007, 23, 471–472. [Google Scholar] [CrossRef]
- Longo-Pendy, N.M.; Boundenga, L.; Kutomy, P.O.O.; Mbou-Boutambe, C.; Makanga, B.; Moukodoum, N.; Obame-Nkoghe, J.; Makouloutou, P.N.; Mounioko, F.; Akone-Ella, R. Systematic Review on Diversity and Distribution of Anopheles Species in Gabon: A Fresh Look at the Potential Malaria Vectors and Perspectives. Pathogens 2022, 11, 668. [Google Scholar] [CrossRef]
- Mourou, J.-R.; Coffinet, T.; Jarjaval, F.; Cotteaux, C.; Pradines, E.; Godefroy, L.; Kombila, M. Malaria transmission in Libreville: Results of a one year survey. Malar. J. 2012, 11, 40. [Google Scholar] [CrossRef]
- Service, M. Contribution to the knowledge of the mosquitoes (Diptera, Culicidae) of Gabon. Cah. Orstom. Sér. Entomol. Méd. Parasitol 1976, 3, 259–263. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Nature | Inside the Forest | Outside the Forest | Overall |
|---|---|---|---|
| (n = 28) | (n = 76) | (n = 104) | |
| Artificial | |||
| Glass jars | 1 (5.0%) | 1 (1.4%) | 2 (2.1%) |
| Gutters | 1 (5.0%) | 16 (21.6%) | 17 (18.1%) |
| Metallic containers | 0 (0.0%) | 6 (8.1%) | 6 (6.4%) |
| Plastic containers | 15 (75.0%) | 17 (23.0%) | 32 (34.0%) |
| Puddles | 0 (0.0%) | 18 (24.3%) | 18 (19.1%) |
| Tires | 0 (0.0%) | 14 (18.9%) | 14 (14.9%) |
| Wash basins | 1 (5.0%) | 1 (1.4%) | 2 (2.1%) |
| Discarded freezer | 0 (0.0%) | 1 (1.4%) | 1 (1.1%) |
| Discarded toilet bowls | 2 (10.0%) | 0 (0.0%) | 2 (2.1%) |
| Subtotal | 20 (100%) | 74 (100%) | 94 (100%) |
| Natural | |||
| Puddles | 1 (12.5%) | 0 (0.0%) | 1 (10.0%) |
| Tree holes | 5 (62.5%) | 0 (0.0%) | 5 (50.0%) |
| Streams | 2 (25.0%) | 2 (100%) | 4 (40.0%) |
| Subtotal | 8 (100%) | 2 (100%) | 10 (100%) |
| Species | Inside Forest | Outside Forest | Overall | ||||
|---|---|---|---|---|---|---|---|
| Artificial | Natural | Sub-Total | Artificial | Natural | Sub-Total | ||
| Ae. aegypti | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 27 (4.3%) | 0 (0.0%) | 27 (4.1%) | 27 (3.5%) |
| Ae. albopictus | 67 (77.0%) | 3 (13.6%) | 70 (64.3%) | 188 (29.7%) | 0 (0.0%) | 188 (28.5%) | 258 (33.5%) |
| An. gambiae s. l. | 0 (0.0%) | 1 (4.6%) | 1 (0.9%) | 41 (6.5%) | 0 (0.0%) | 41 (6.2%) | 42 (5.4%) |
| Cx. cinerellus | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 1 (0.1%) | 0 (0.0%) | 1 (0.1%) | 1 (0.1%) |
| Cx. decens | 2 (2.3%) | 0 (0.0%) | 2 (1.8%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 2 (0.3%) |
| Cx. duttoni | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 6 (0.9%) | 0 (0.0%) | 6 (0.9%) | 6 (0.8%) |
| Cx. quinquefasciatus | 12 (13.8%) | 0 (0.0%) | 12 (11.0%) | 222 (35.1%) | 0 (0.0%) | 222 (33.6%) | 234 (30.4%) |
| Cx. tauffliebi | 0 (0.0%) | 11 (50.0%) | 11 (10.1%) | 17 (2.7%) | 19 (67.8%) | 36 (5.4%) | 47 (6.1%) |
| Cx. trifilatus | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 3 (0.5%) | 0 (0.0%) | 3 (0.5%) | 3 (0.4%) |
| Cx. umbripes | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 1 (0.1%) | 0 (0.0%) | 1 (0.1%) | 1 (0.1%) |
| Cx. univittatus | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 4 (0.6%) | 0 (0.0%) | 4 (0.6%) | 4 (0.5%) |
| Culex sp. | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 12 (1.9%) | 1 (3.6%) | 13 (2.0%) | 13 (1.7%) |
| Er. inornatus | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 3 (0.5%) | 0 (0.0 %) | 3 (0.5%) | 3 (0.4%) |
| Lu. tigripes | 6 (6.9%) | 5 (22.7%) | 11 (10.1%) | 108 (17.1%) | 8 (28.6%) | 116 (17.5%) | 127 (16.5%) |
| Tx. evansae | 0 (0.0%) | 2 (9.1%) | 2 (1.8%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 2 (0.3%) |
| Total | 87 (100%) | 22 (100%) | 109 (100%) | 633 (100%) | 28 (100%) | 661 (100%) | 770 (100%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obame-Nkoghe, J.; Makanga, B.K.; Zongo, S.B.; Koumba, A.A.; Komba, P.; Longo-Pendy, N.-M.; Mounioko, F.; Akone-Ella, R.; Nkoghe-Nkoghe, L.C.; Ngangue-Salamba, M.-F.; et al. Urban Green Spaces and Vector-Borne Disease Risk in Africa: The Case of an Unclean Forested Park in Libreville (Gabon, Central Africa). Int. J. Environ. Res. Public Health 2023, 20, 5774. https://doi.org/10.3390/ijerph20105774
Obame-Nkoghe J, Makanga BK, Zongo SB, Koumba AA, Komba P, Longo-Pendy N-M, Mounioko F, Akone-Ella R, Nkoghe-Nkoghe LC, Ngangue-Salamba M-F, et al. Urban Green Spaces and Vector-Borne Disease Risk in Africa: The Case of an Unclean Forested Park in Libreville (Gabon, Central Africa). International Journal of Environmental Research and Public Health. 2023; 20(10):5774. https://doi.org/10.3390/ijerph20105774
Chicago/Turabian StyleObame-Nkoghe, Judicaël, Boris Kevin Makanga, Sylvie Brizard Zongo, Aubin Armel Koumba, Prune Komba, Neil-Michel Longo-Pendy, Franck Mounioko, Rodolphe Akone-Ella, Lynda Chancelya Nkoghe-Nkoghe, Marc-Flaubert Ngangue-Salamba, and et al. 2023. "Urban Green Spaces and Vector-Borne Disease Risk in Africa: The Case of an Unclean Forested Park in Libreville (Gabon, Central Africa)" International Journal of Environmental Research and Public Health 20, no. 10: 5774. https://doi.org/10.3390/ijerph20105774
APA StyleObame-Nkoghe, J., Makanga, B. K., Zongo, S. B., Koumba, A. A., Komba, P., Longo-Pendy, N.-M., Mounioko, F., Akone-Ella, R., Nkoghe-Nkoghe, L. C., Ngangue-Salamba, M.-F., Yangari, P., Aboughe-Angone, S., Fournet, F., Kengne, P., & Paupy, C. (2023). Urban Green Spaces and Vector-Borne Disease Risk in Africa: The Case of an Unclean Forested Park in Libreville (Gabon, Central Africa). International Journal of Environmental Research and Public Health, 20(10), 5774. https://doi.org/10.3390/ijerph20105774