
Lead Responses and Tolerance Mechanisms of Koelreuteria paniculata: A Newly Potential Plant for Sustainable Phytoremediation of Pb-Contaminated Soil

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Design
2.2. Experimental Method
2.2.1. Plant Height, Biomass, and Root Length
2.2.2. Pb Changes in Subcellular Fractions in Each Tissue of the Plant
2.2.3. Determination of Chlorophyll and Antioxidant Enzyme in Plant Leaves
2.2.4. Microstructure and Functional Groups Analysis of the Plants
2.3. Statistics Analysis
3. Results
3.1. Effects of Lead Stress on Plant Height and Biomass
3.2. Effects of Lead Stress on Root Morphology
3.3. Enrichment Capacity of K. paniculata for Pb
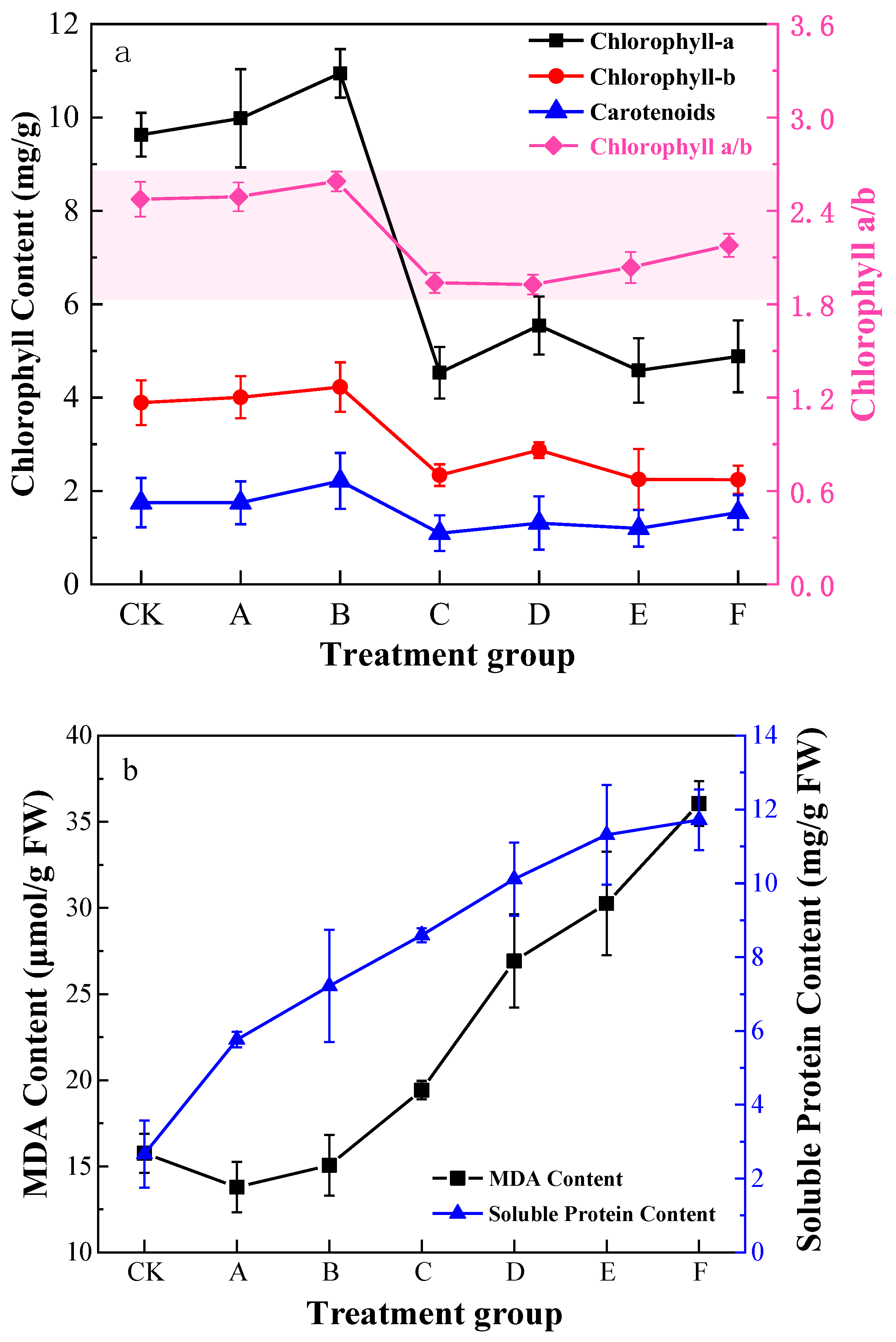
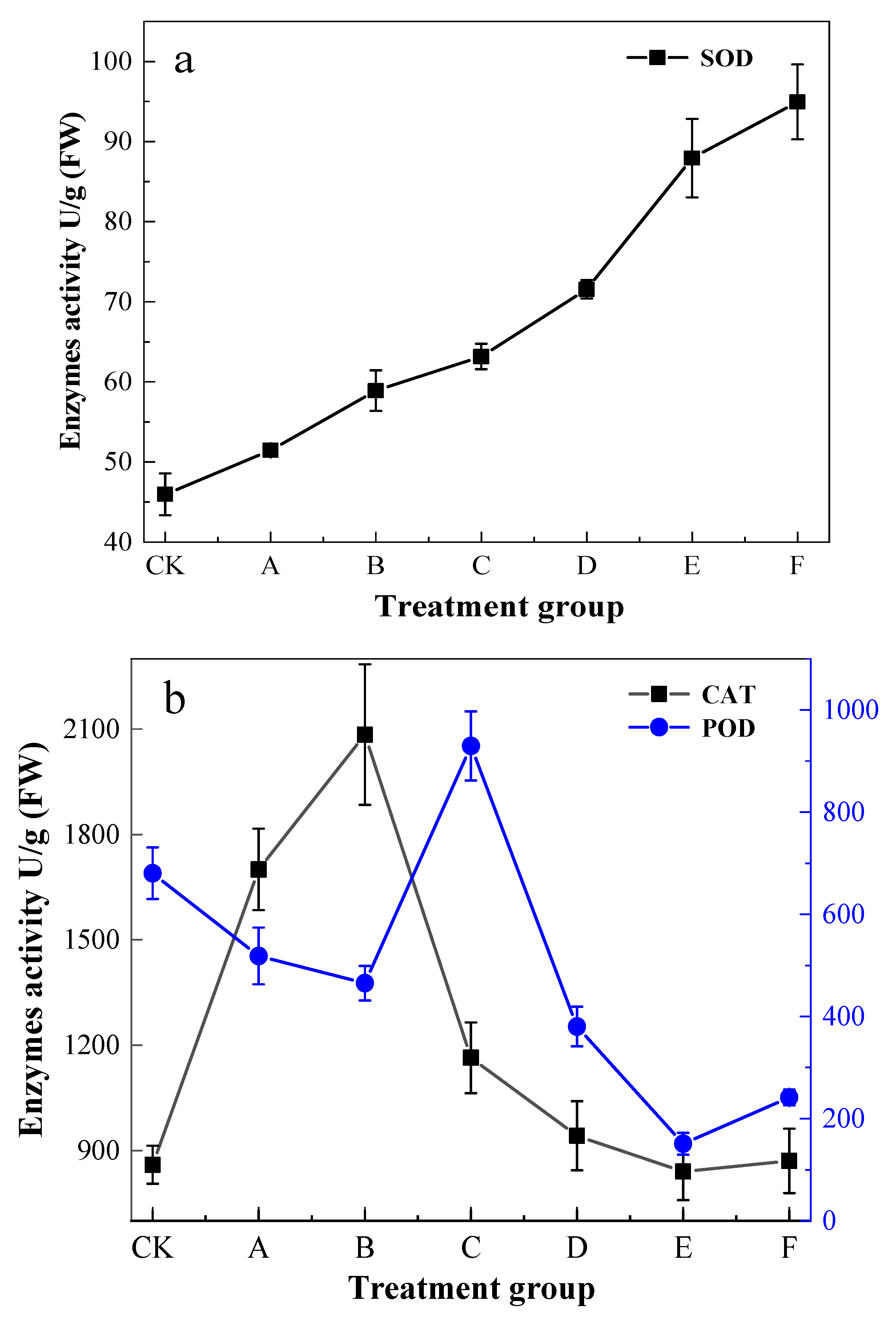
3.4. Physiological Indexes of Pb Stress in K. paniculata Leaves
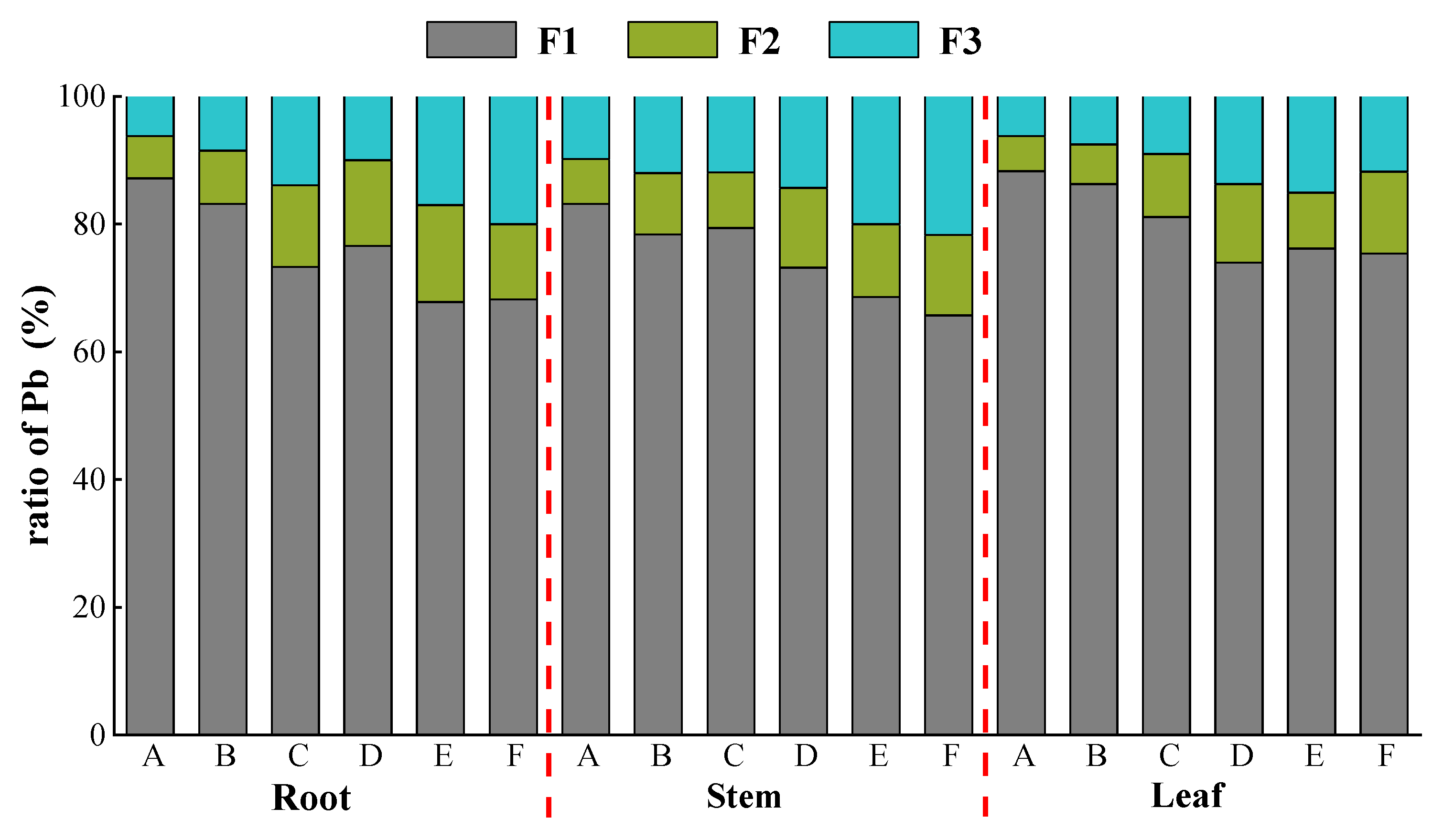
3.5. Effects of Lead Stress on Subcellular Distribution
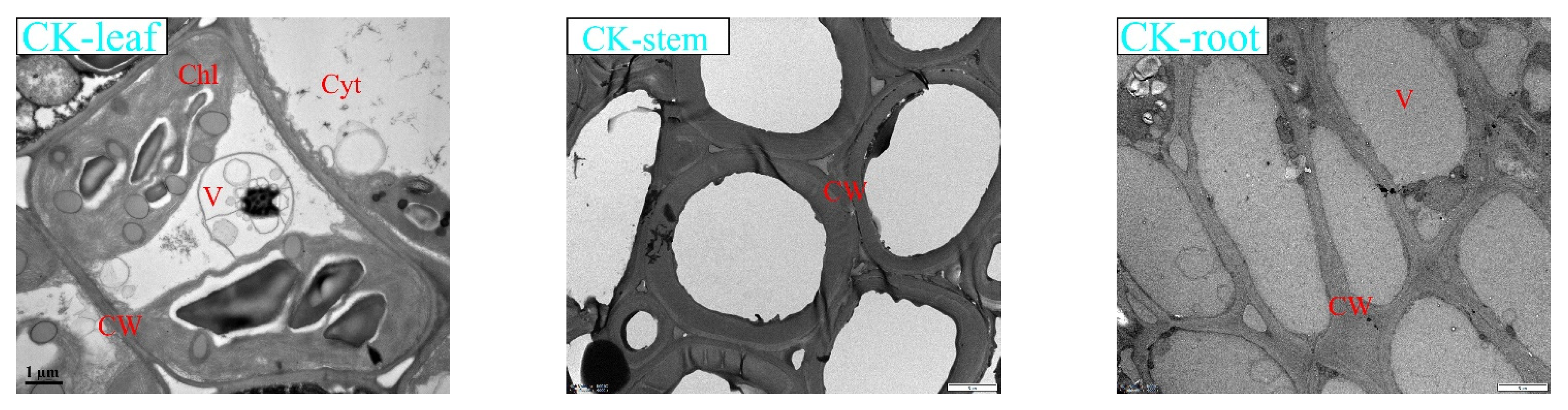
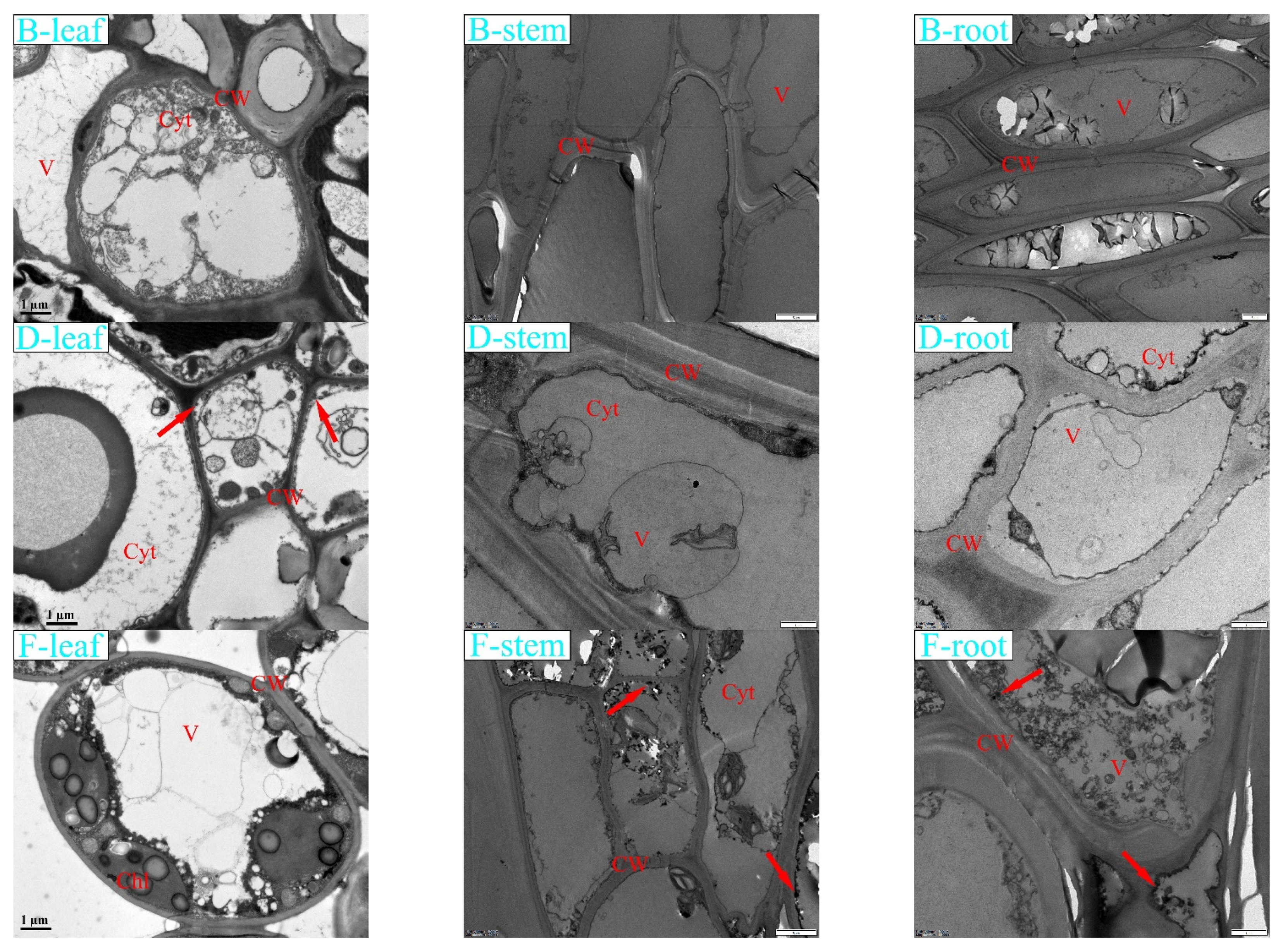
3.6. Effects of Lead Stress on K. paniculata Microstructure
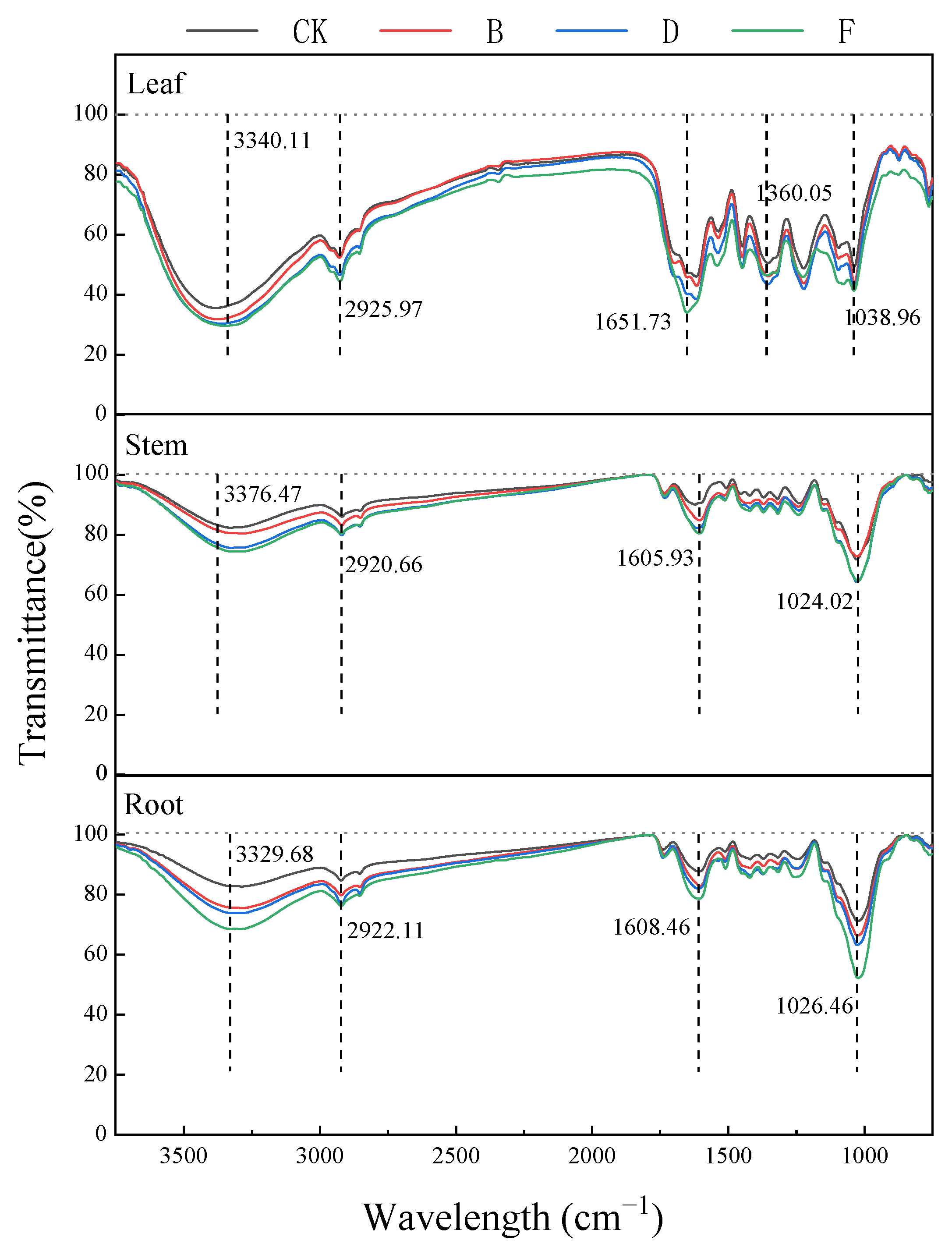
3.7. FTIR Analysis of Functional Group Composition of K. paniculata Tissue
4. Discussion
4.1. Effects on the Growth of K. paniculata under Pb Stress
4.2. Defense Mechanisms of K. paniculata Leaves under Pb Stress
4.3. Microcosmic Structure Response of K. paniculata under Pb Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, X.; An, J.; Yin, Y.; Gao, C.; Wang, B.; Wei, S. Heavy metals uptake and translocation of typical wetland plants and their ecological effects on the coastal soil of a contaminated bay in Northeast China. Sci. Total Environ. 2022, 803, 149871. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Ren, Q.; Zheng, K.; Qin, Z.; Wang, Y.; Wang, Y. Spatial distribution and risk assessment of metal(loid)s in marine sediments in the Arctic Ocean and Bering Sea. Mar. Pollut. Bull. 2022, 179, 113729. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Xie, C.; Alhassan, S.I.; Huang, S.; Chen, R.; Xiang, S.; Wang, Z.; Huang, L. Oxygen reduction reaction in the field of water environment for application of nanomaterials. Nanomaterials 2020, 10, 1719. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, S.; Muhammad, I.; Muhammad, R.S.; Wajid, I.; Muhammad, A.K.; Amar, M.; Abdur, R.; Saddam, H.E. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar]
- Gu, J.; Zhang, X.; Han, J.; Zhao, Z. Global lead resources situation and the development of lead resources in China. China Min. Mag. 2017, 26, 16–20+44. [Google Scholar]
- Wang, F.; Dong, W.; Zhao, Z.; Wang, H.; Li, W.; Chen, G.; Wang, F.; Zhao, Y.; Huang, J.; Zhou, T. Heavy metal pollution in urban river sediment of different urban functional areas and its influence on microbial community structure. Sci. Total Environ. 2021, 778, 146383. [Google Scholar] [CrossRef]
- Mudd, G.M.; Jowitt, S.M.; Werner, T.T. The world’s lead-zinc mineral resources: Scarcity, data, issues and opportunities. Ore Geol. Rev. 2017, 80, 1160–1190. [Google Scholar] [CrossRef]
- Gyamfi, E.; Appiah-Adjei, E.K.; Adjei, K.A. Potential heavy metal pollution of soil and water resources from artisanal mining in Kokoteasua, Ghana. Groundw. Sustain. Dev. 2019, 8, 450–456. [Google Scholar] [CrossRef]
- Su, R.; Ou, Q.; Wang, H.; Luo, Y.; Dai, X.; Wang, Y.; Chen, Y.; Shi, L. Comparison of phytoremediation potential of Nerium indicum with inorganic modifier calcium carbonate and organic modifier mushroom residue to lead-zinc tailings. Int. J. Environ. Res. Public Health 2022, 19, 10353. [Google Scholar] [CrossRef]
- Liu, L.; Wei, L.; Song, W.; Guo, M. Remediation techniques for heavy metal-contaminated soils: Principles and applicability. Sci. Total Environ. 2018, 633, 206–219. [Google Scholar] [CrossRef]
- Collin, M.S.; Venkataraman, S.K.; Vijayakumar, N.; Kanimozhi, V.; Arbaaz, S.M.; Stacey, R.G.S.; Anusha, J.; Choudhary, R.; Lvov, V.; Tovar, G.I.; et al. Bioaccumulation of lead (Pb) and its effects on human: A review. JHM Adv. 2022, 7, 100094. [Google Scholar] [CrossRef]
- Yang, L.; Ren, Q.; Zheng, K.; Jiao, Z.; Ruan, X.; Wang, Y. Migration of heavy metals in the soil-grape system and potential health risk assessment. Sci. Total Environ. 2022, 806, 150646. [Google Scholar] [CrossRef]
- Lanphear, B.P.; Rauch, S.; Auinger, P.; Allen, R.W.; Hornung, R.W. Low-level lead exposure and mortality in US adults: A population-based cohort study. Lancet Public Health 2018, 3, 177–184. [Google Scholar] [CrossRef]
- Shen, Z.; Jin, F.; O’Connor, D.; Hou, D. Solidification/stabilization for soil remediation: An old technology with new vitality. Environ. Sci. Technol. 2019, 53, 11615–11617. [Google Scholar] [CrossRef]
- Li, Y.; Liao, X.; Li, W. Combined sieving and washing of multi-metal-contaminated soils using remediation equipment: A pilot-scale demonstration. J. Clean. Prod. 2019, 212, 81–89. [Google Scholar] [CrossRef]
- Lian, M.; Feng, Q.; Wang, L.; Niu, L.; Zhao, Z.; Li, X.; Zhang, Z. Highly effective immobilization of Pb and Cd in severely contaminated soils by environment-compatible, mercapto-functionalized reactive nanosilica. J. Clean. Prod. 2019, 235, 583–589. [Google Scholar] [CrossRef]
- Jiang, N.-J.; Liu, R.; Du, Y.-J.; Bi, Y.-Z. Microbial induced carbonate precipitation for immobilizing Pb contaminants: Toxic effects on bacterial activity and immobilization efficiency. Sci. Total Environ. 2019, 672, 722–731. [Google Scholar] [CrossRef]
- Zeng, G.; Wan, J.; Huang, D.; Hu, L.; Huang, C.; Cheng, M.; Xue, W.; Gong, X.; Wang, R.; Jiang, D. Precipitation, adsorption and rhizosphere effect: The mechanisms for phosphate-induced Pb immobilization in soils—A review. J. Hazard. Mater. 2017, 339, 354–367. [Google Scholar] [CrossRef]
- Li, L.; Han, L.; Liu, A.; Wang, F. Imperfect but hopeful: New advances in soil pollution and remediation. Int. J. Environ. Res. Public Health 2022, 19, 10164. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, S.; Cheng, P.; Zhang, S.; Sun, Y. Effects of soil amendments on heavy metal immobilization and accumulation by maize grown in a multiple-metal-contaminated soil and their potential for safe crop production. Toxics 2020, 8, 102. [Google Scholar] [CrossRef]
- Sun, Y.; Zheng, F.; Wang, W.; Zhang, S.; Wang, F. Remediation of Cr(VI)-contaminated soil by nano-zero-valent iron in combination with biochar or humic acid and the consequences for plant performance. Toxics 2020, 8, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Lambers, H.; Wong, W.S.; Dixon, K.W.; Stevens, J.C.; Cross, A.T. Initiating pedogenesis of magnetite tailings using Lupinus angustifolius (narrow-leaf lupin) as an ecological engineer to promote native plant establishment. Sci. Total Environ. 2021, 788, 147622. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.H.; Wang, Y.T.; Lian, Z.H.; Yang, T.T.; Zeng, Q.W.; Feng, S.W.; Fang, Z.; Shu, W.S.; Huang, L.N.; Ye, Z.H.; et al. Revegetation approach and plant identity unequally affect structure, ecological network and function of soil microbial community in a highly acidified mine tailings pond. Sci. Total Environ. 2020, 744, 140793. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Y. Occurrence of arbuscular mycorrhizal fungi in mining-impacted sites and their contribution to ecological restoration: Mechanisms and applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Laghlimi, M.; Baghdad, B.; Hadi, H.E.; Boua Bd Li, A. Phytoremediation mechanisms of heavy metal contaminated soils: A review. Open J. Ecol. 2015, 5, 375–388. [Google Scholar] [CrossRef]
- He, L.; Su, R.; Chen, Y.; Zeng, P.; Du, L.; Cai, B.; Zhang, A.; Zhu, H. Integration of manganese accumulation, subcellular distribution, chemical forms, and physiological responses to understand manganese tolerance in Macleaya cordata. Environ. Sci. Pollut. Res. 2022, 29, 39017–39026. [Google Scholar] [CrossRef]
- Su, R.; Wang, Y.; Huang, S.; Chen, R.; Wang, J. Application for ecological restoration of contaminated soil: Phytoremediation. Int. J. Environ. Res. Public Health 2022, 19, 13124. [Google Scholar] [CrossRef]
- Dou, X.; Dai, H.; Skuza, L.; Wei, S. Cadmium removal potential of hyperaccumulator Solanum nigrum L. under two planting modes in three years continuous phytoremediation. Environ. Pollut. 2022, 307, 119493. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker AJ, M.; Jaffre, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef]
- Shen, X.; Dai, M.; Yang, J.; Sun, L.; Tan, X.; Peng, C.; Ali, I.; Naz, I. A critical review on the phytoremediation of heavy metals from environment: Performance and challenges. Chemosphere 2022, 291, 132979. [Google Scholar] [CrossRef]
- Oladoye, P.O.; Olowe, O.M.; Asemoloye, M.D. Phytoremediation technology and food security impacts of heavy metal contaminated soils: A review of literature. Chemosphere 2022, 288, 132555. [Google Scholar] [CrossRef]
- Jiang, Y.B.; Jiang, S.M.; Li, Z.B.; Yan, X.P.; Qin, Z.X.; Huang, R.Z. Field scale remediation of Cd and Pb contaminated paddy soil using three mulberry (Morus alba L.) cultivars. Ecol. Eng. 2019, 129, 38–44. [Google Scholar] [CrossRef]
- Heckenroth, A.; Rabier, J.; Dutoit, T.; Torre, F.; Prudent, P.; Laffont-Schwob, I. Selection of native plants with phytoremediation potential for highly contaminated Mediterranean soil restoration: Tools for a non-destructive and integrative approach. J. Environ. Manag. 2016, 183, 850–863. [Google Scholar] [CrossRef]
- Zhang, X.; Lou, X.; Zhang, H.; Ren, W.; Tang, M. Effects of sodium sulfide application on the growth of Robinia pseudoacacia, heavy metal immobilization, and soil microbial activity in Pb-Zn polluted soil. Ecotoxicol. Environ. Saf. 2020, 197, 110563. [Google Scholar] [CrossRef]
- Kang, W.; Bao, J.; Zheng, J.; Xu, F.; Wang, L. Phytoremediation of heavy metal contaminated soil potential by woody plants on Tonglushan ancient copper spoil heap in China. Int. J. Phytoremediat. 2016, 20, 1–7. [Google Scholar] [CrossRef]
- Al-Lami, M.K.; Oustriere, N.; Gonzales, E.; Burken, J.G. Phytomanagement of Pb/Zn/Cu tailings using biosolids-biochar or -humus combinations: Enhancement of bioenergy crop production, substrate functionality, and ecosystem services. Sci. Total Environ. 2022, 836, 155676. [Google Scholar] [CrossRef]
- Liu, T.; Zhu, F.; Yan, W.; Chen, X.; Huang, X. Assessment of Koelreuteria paniculata seedling for phytroremediation of pyrene-contaminated soils. Water Air Soil Pollut. 2018, 229, 396. [Google Scholar] [CrossRef]
- Kiboi, M.N.; Ngetich, F.K.; Mucheru-Muna, M.W.; Diels, J.; Mugendi, D.N. Soil nutrients and crop yield response to conservation-effective management practices in the sub-humid highlands agro-ecologies of Kenya. Heliyon 2021, 7, e07156. [Google Scholar] [CrossRef]
- Tang, C.; Chen, Y.; Zhang, Q.; Li, J.; Zhang, F.; Liu, Z. Effects of peat on plant growth and lead and zinc phytostabilization from lead-zinc mine tailing in southern China: Screening plant species resisting and accumulating metals. Ecotoxicol. Environ. Saf. 2019, 176, 42–49. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, Y.; Du, L.; Zhang, M.; Han, L. Accumulation and subcellular distribution of heavy metal in Paulownia fortunei cultivated in lead-zinc slag amended with peat. Int. J. Phytoremediat. 2019, 21, 1153–1160. [Google Scholar] [CrossRef]
- Han, L.; Chen, Y.; Chen, M.; Wu, Y.; Su, R.; Du, L.; Liu, Z. Mushroom residue modification enhances phytoremediation potential of Paulownia fortunei to lead-zinc slag. Chemosphere 2020, 253, 126774. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Liu, Y.; Zeng, G.; Zheng, B.; Tan, X.; Liu, H.; Xie, J.; Gan, C.; Liu, W. Cadmium accumulation and tolerance of Macleaya cordata: A newly potential plant for sustainable phytoremediation in Cd-contaminated soil. Environ. Sci. Pollut. Res. 2016, 23, 10189–10199. [Google Scholar] [CrossRef] [PubMed]
- Dou, C.M.; Fu, X.P.; Chen, X.C.; Shi, J.Y.; Chen, Y.X. Accumulation and detoxification of manganese in hyperaccumulator Phytolacca americana. Plant Biol. 2010, 11, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Ghori, N.H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy metal stress and responses in plants. Int. J. Enivron. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- James, J.J.; Drenovsky, R.E. A basis for relative growth rate differences between native and invasive forb seedlings. Rangel. Ecol. Manag. 2007, 60, 395–400. [Google Scholar] [CrossRef]
- Wang, L.; Kang, Y.; Liang, S.; Chen, D.; Zhang, Q.; Zeng, L.; Luo, J.; Jiang, F. Synergistic effect of co-exposure to cadmium (II) and 4-n-nonylphenol on growth inhibition and oxidative stress of Chlorella sorokiniana. Ecotoxicol. Environ. Saf. 2018, 154, 145–153. [Google Scholar] [CrossRef]
- Maurya, P.; Kumari, R. Toxic metals distribution, seasonal variations and environmental risk assessment in surficial sediment and mangrove plants (A. marina), Gulf of Kachchh (India). J. Hazard. Mater. 2021, 413, 125345. [Google Scholar] [CrossRef]
- Ma, X.; Li, X.; Liu, J.; Cheng, Y.; Zhai, F.; Sun, Z.; Han, L. Enhancing Salix viminalis L.–mediated phytoremediation of polycyclic aromatic hydrocarbon–contaminated soil by inoculation with Crucibulum laeve (white-rot fungus). Environ. Sci. Pollut. Res. 2020, 27, 41326–41341. [Google Scholar] [CrossRef]
- Vaculik, M.; Konlechner, C.; Langer, I.; Adlassnig, W.; Puschenreiter, M.; Lux, A.; Hauser, M.T. Root anatomy and element distribution vary between two Salix caprea isolates with different Cd accumulation capacities. Environ. Pollut. 2012, 163, 117–126. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Xu, Z.; Wang, Y.; Teng, Z.; An, M.; Zhang, Y.; Zhu, W.; Xu, N.; Sun, G. Toxic effects of heavy metals Pb and Cd on mulberry (Morus alba L.) seedling leaves: Photosynthetic function and reactive oxygen species (ROS) metabolism responses. Ecotoxicol. Environ. Saf. 2020, 195, 110469. [Google Scholar]
- Zhou, Y.; Wang, Y.; Inyang, A.I. Ecophysiological differences between five mangrove seedlings under heavy metal stress. Mar. Pollut. Bull. 2021, 172, 112900. [Google Scholar] [CrossRef]
- Chen, H.; Lee, J.; Lee, J.-M.; Han, M.; Emonet, A.; Lee, J.; Jia, X.; Lee, Y. MSD2, an apoplastic Mn-SOD, contributes to root skotomorphogenic growth by modulating ROS distribution in Arabidopsis. Plant Sci. 2022, 317, 111192. [Google Scholar] [CrossRef]
- Mahdavian, K.; Ghaderian, S.M.; Schat, H. Pb accumulation, Pb tolerance, antioxidants, thiols, and organic acids in metallicolous and non-metallicolous Peganum harmala L. under Pb exposure. Environ. Exp. Bot. 2016, 126, 21–31. [Google Scholar] [CrossRef]
- Vallee, B.L.; Ulmer, D.D. Biochemical effects of mercury, cadmium, and lead. Annu. Rev. Biochem. 1972, 41, 91. [Google Scholar] [CrossRef]
- Peralta, J.R.; Gardea-Torresdey, J.L.; Tiemann, K.J.; Gomez, E.; Arteaga, S.; Rascon, E.; Parsons, J.G. Uptake and effects of five heavy metals on seed germination and plant growth in alfalfa (Medicago sativa L.). Bull. Environ. Contam. Toxicol. 2001, 66, 727–734. [Google Scholar] [CrossRef]
- Yang, P.; Li, Y.; He, C.; Yan, J.; Zhang, W.; Li, X.; Xiang, F.; Zuo, Z.; Li, X.; Zhu, Y.; et al. Phenotype and TMT-based quantitative proteomics analysis of Brassica napus reveals new insight into chlorophyll synthesis and chloroplast structure. J. Proteom. 2020, 214, 103621. [Google Scholar] [CrossRef]
- Shi, Y.; Mu, X.; Li, K.; Shao, H. Soil characterization and differential patterns of heavy metal accumulation in woody plants grown in coal gangue wastelands in Shaanxi, China. Environ. Sci. Pollut. Res. 2016, 23, 13489–13497. [Google Scholar]
- Zhao, X.; Chen, B.; Huo, X.; Zhao, R.; Hua, X. Effects of exogenous selenium on physiological characteristics and absorption of lead and selenium in Chinese Cabbage under lead stress. J. Irrig. Drain. 2018, 37, 6. [Google Scholar]
- Liu, Y.; Sun, X.; Li, S.; Li, S.; Zhou, W.; Ma, Q.; Zhang, J. Influence of green waste compost on Pb-polluted soil remediation, soil quality improvement, and uptake by Pakchoi cabbage (Brassica campestris L. ssp.). Environ. Sci. Pollut. Res. 2020, 27, 7693–7701. [Google Scholar] [CrossRef]
- Sharma, P.; Kumar, S. Bioremediation of heavy metals from industrial effluents by endophytes and their metabolic activity: Recent advances. Bioresour. Technol. 2021, 339, 125589. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, F.; Liu, J.-L.; Wu, H.-T.; Yang, H.; Shi, Y.; Liu, J.; Zhang, Y.-F.; Luo, Y.-R.; Chen, K.-M. Heavy metal transporters: Functional mechanisms, regulation, and application in phytoremediation. Sci. Total Environ. 2022, 809, 151099. [Google Scholar] [CrossRef] [PubMed]
- Abdelkrim, S.; Jebara, S.H.; Saadani, O.; Chiboub, M.; Abid, G.; Mannai, K.; Jebara, M. Heavy metal accumulation in Lathyrus sativus growing in contaminated soils and identification of symbiotic resistant bacteria. Arch. Microbiol. 2019, 201, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Yan, W.; Zhang, H.; Xiao, Z.; Zheng, L. Subcellular distribution and chemical forms involved in manganese accumulation and detoxification for Xanthium strumarium L. Chemosphere 2019, 237, 124531. [Google Scholar] [CrossRef] [PubMed]
- Cosio, C. Hyperaccumulation of cadmium and zinc in Thlaspi caerulescens and Arabidopsis halleri at the leaf cellular level. Plant Physiol. 2004, 134, 716–725. [Google Scholar] [CrossRef]
- Li, Y.; Shao, M.; Huang, M.; Sang, W.; Zheng, S.; Jiang, N.; Gao, Y. Enhanced remediation of heavy metals contaminated soils with EK-PRB using β-CD/hydrothermal biochar by waste cotton as reactive barrier. Chemosphere 2022, 286, 131470. [Google Scholar] [CrossRef]
- Bosch, A.; Serra, D.; Prieto, C.; Schmitt, J.; Naumann, D.; Yantorno, O. Characterization of Bordetella pertussis growing as biofilm by chemical analysis and FT-IR spectroscopy. Appl. Microbiol. Biotechnol. 2006, 71, 736–747. [Google Scholar] [CrossRef]
- Xue, C.; He, C.Q.; Shi, Z.; Chen, X.; Muo, Q. Effect of spent mushroom substrate on strengthening the phytoremediation potential of Ricinus communis to Cd- and Zn-polluted soil. Int. J. Phytoremediat. 2019, 20, 1389–1399. [Google Scholar]
- Chandrasekhar, C.; Ray, J.G. Lead accumulation, growth responses and biochemical changes of three plant species exposed to soil amended with different concentrations of lead nitrate. Ecotoxicol. Environ. Saf. 2019, 171, 26–36. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Average Height (cm) | Total Biomass (g) | Root Biomass (g) | Stem Biomass (g) | Leaf Biomass (g) |
|---|---|---|---|---|---|
| CK | 42.54 ± 0.67 c | 9.71 ± 0.71 b | 4.98 ± 0.33 a | 2.64 ± 0.12 c | 2.08 ± 0.1 a |
| A | 52.67 ± 0.85 a | 11.05 ± 1.045 a | 5.28 ± 0.21 a | 3.60 ± 0.1 a | 2.17 ± 0.06 a |
| B | 47.95 ± 1.95 b | 10.14 ± 0.45 ab | 5.15 ± 0.38 a | 3.33 ± 0.16 b | 1.65 ± 0.11 a |
| C | 38.40 ± 1.01 d | 8.50 ± 0.54 c | 4.49 ± 0.16 b | 2.66 ± 0.15 c | 1.34 ± 0.17 a |
| D | 37.60 ± 2.0 d | 7.04 ± 0.59 d | 3.93 ± 0.14 c | 2.03 ± 0.15 d | 1.08 ± 5.26 a |
| E | 37.20 ± 0.85 d | 6.60 ± 0.37 de | 4.07 ± 0.3 c | 1.61 ± 0.04 e | 0.92 ± 0.4 a |
| F | 31.80 ± 1.39 e | 5.70 ± 0.3 e | 3.39 ± 0.28 d | 1.45 ± 0.78 e | 0.85 ± 0.18 a |
| Treatment Group | Total Root Length (cm) | Total Root Surface Area (cm2) | Average Root Diameter (mm) | Number of Fine Roots |
|---|---|---|---|---|
| CK | 3079.01 ± 169.49 b | 576.04 ± 115.39 a | 1.32 ± 0.80 e | 1621 ± 117.20 ab |
| A | 3883.7 ± 429.61 a | 665.97 ± 98.48 a | 1.21 ± 1.10 e | 1803 ± 187.19 a |
| B | 2705.99 ± 122.29 b | 558.89 ± 46.63 a | 1.46 ± 1.50 de | 1413 ± 188.56 b |
| C | 1621.44 ± 179.88 c | 396.78 ± 20.36 b | 1.73 ± 0.29 cd | 995 ± 169.14 c |
| D | 1040.36 ± 247.13 d | 330.42 ± 54.04 bc | 2.02 ± 0.08 bc | 641 ± 107.53 d |
| E | 872.65 ± 123.18 de | 308.16 ± 51.49 bc | 2.25 ± 0.37 b | 582 ± 27.78 de |
| F | 514.46 ± 83.001 e | 215.84 ± 24.76 c | 2.67 ± 0.27 a | 365 ± 50.10 e |
| Treatment Group | Pb Content (mg/kg) | BCF | TF | Total Accumulation Amount (Average Value, mg) | ||
|---|---|---|---|---|---|---|
| Root | Stem | Leaf | ||||
| A | 540.67 ± 13.05 e | 59.88 ± 5.38 e | 20.85 ± 4.49 f | 0.5 ± 0.014 b | 0.07 ± 0.003 c | 3.12 |
| B | 743.54 ± 55.48 e | 106.76 ± 1.75 d | 64.157 ± 5.49 e | 0.65 ± 0.013 a | 0.13 ± 0.01 a | 4.29 |
| C | 1143.25 ± 186.31 d | 119.24 ± 19.86 d | 93.21 ± 3.29 d | 0.46 ± 0.05 b | 0.09 ± 0.01 b | 5.58 |
| D | 1738.04 ± 192.28 c | 165.84 ± 7.81 c | 143.88 ± 17.16 c | 0.4 ± 0.02 c | 0.08 ± 0.005 bc | 7.32 |
| E | 2338.69 ± 151.27 b | 280.78 ± 16.7 b | 188.22 ± 24.12 b | 0.36 ± 0.05 c | 0.08 ± 0.012 bc | 10.14 |
| F | 3187.87 ± 251.77 a | 389.46 ± 21.7 a | 253.11 ± 7.81 a | 0.36 ± 0.12 c | 0.07 ± 0.003 c | 11.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, R.; Xie, T.; Yao, H.; Chen, Y.; Wang, H.; Dai, X.; Wang, Y.; Shi, L.; Luo, Y. Lead Responses and Tolerance Mechanisms of Koelreuteria paniculata: A Newly Potential Plant for Sustainable Phytoremediation of Pb-Contaminated Soil. Int. J. Environ. Res. Public Health 2022, 19, 14968. https://doi.org/10.3390/ijerph192214968
Su R, Xie T, Yao H, Chen Y, Wang H, Dai X, Wang Y, Shi L, Luo Y. Lead Responses and Tolerance Mechanisms of Koelreuteria paniculata: A Newly Potential Plant for Sustainable Phytoremediation of Pb-Contaminated Soil. International Journal of Environmental Research and Public Health. 2022; 19(22):14968. https://doi.org/10.3390/ijerph192214968
Chicago/Turabian StyleSu, Rongkui, Tianzhi Xie, Haisong Yao, Yonghua Chen, Hanqing Wang, Xiangrong Dai, Yangyang Wang, Lei Shi, and Yiting Luo. 2022. "Lead Responses and Tolerance Mechanisms of Koelreuteria paniculata: A Newly Potential Plant for Sustainable Phytoremediation of Pb-Contaminated Soil" International Journal of Environmental Research and Public Health 19, no. 22: 14968. https://doi.org/10.3390/ijerph192214968
APA StyleSu, R., Xie, T., Yao, H., Chen, Y., Wang, H., Dai, X., Wang, Y., Shi, L., & Luo, Y. (2022). Lead Responses and Tolerance Mechanisms of Koelreuteria paniculata: A Newly Potential Plant for Sustainable Phytoremediation of Pb-Contaminated Soil. International Journal of Environmental Research and Public Health, 19(22), 14968. https://doi.org/10.3390/ijerph192214968