


How Do Landscape Heterogeneity, Community Structure, and Topographical Factors Contribute to the Plant Diversity of Urban Remnant Vegetation at Different Scales?


Abstract
1. Introduction
2. Data and Methods
2.1. Study Area
2.2. Sampling and Vegetation Survey
2.3. Landscape Pattern Metrics Combined with Community Structure and Topographic Factors
2.4. Data Analysis
2.4.1. Data Analysis of Sampling and Vegetation Survey
2.4.2. Data Analysis of Landscape Pattern Metrics Combined with Community Structure and Topographic Factors
3. Results
3.1. Species Composition in Different Remnant Vegetation in Guangzhou City
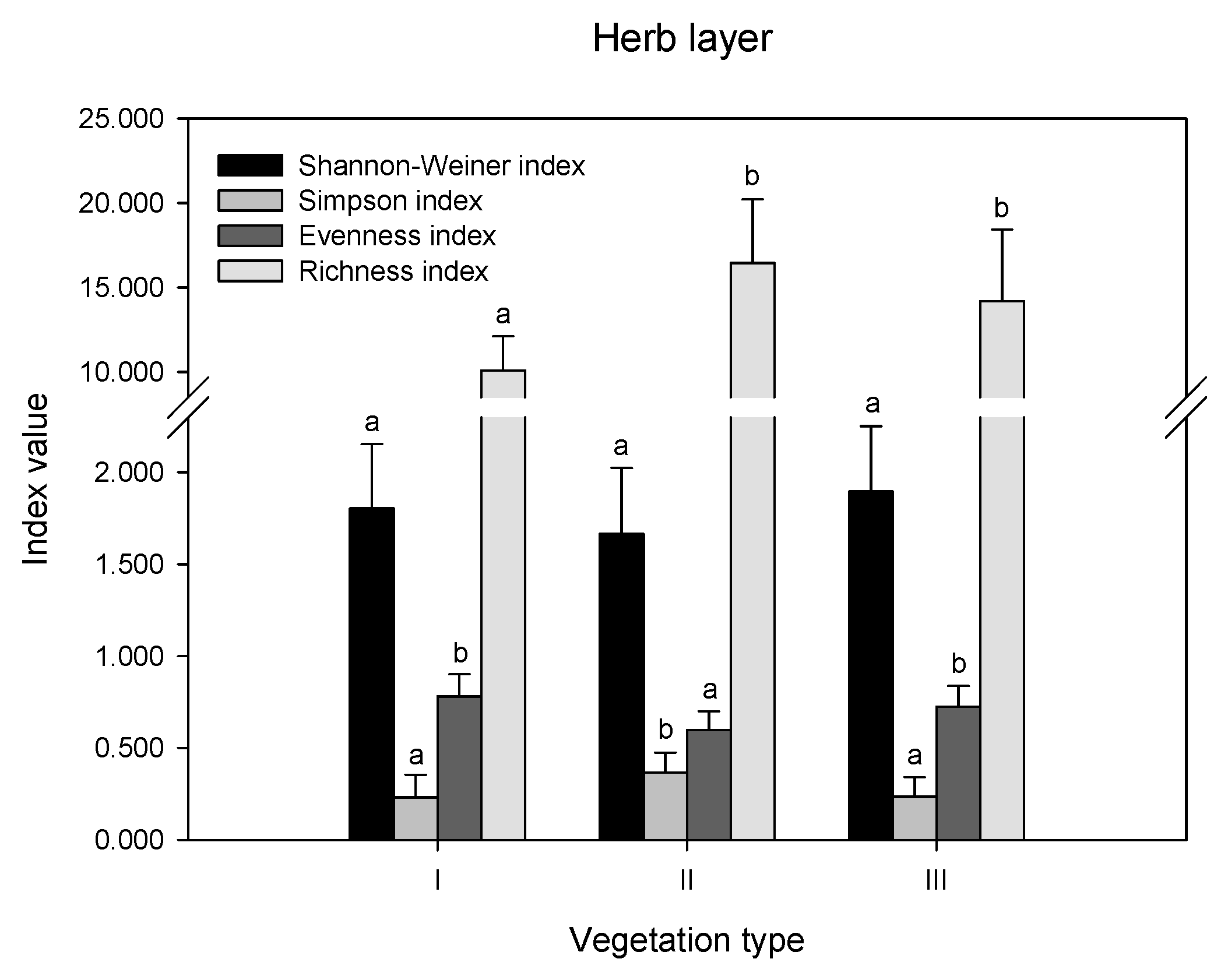
3.2. Plant Diversity in Different Layers of Remnant Vegetation in Guangzhou City
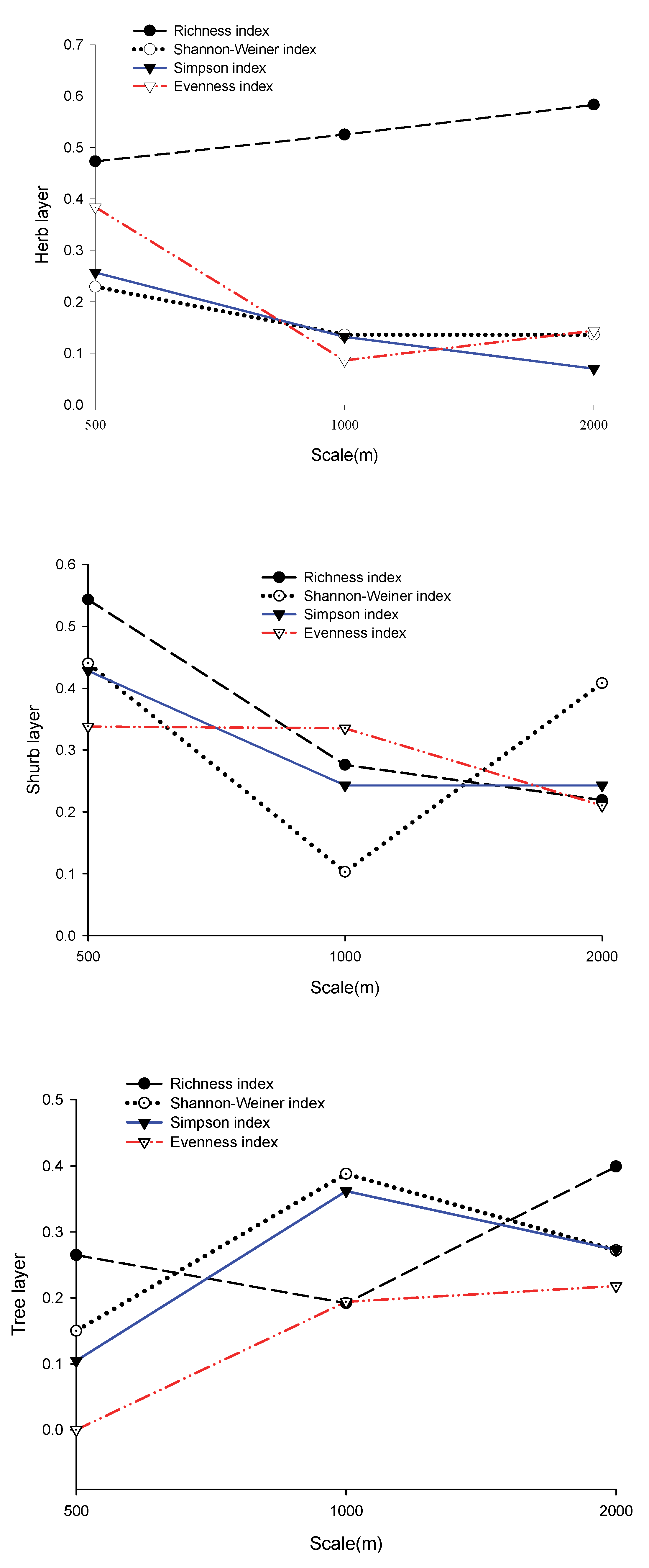
3.3. Relationships between Landscape Heterogeneity, Community Structure, Topographic Factors, and Plant Diversity Indices
3.4. The Explanatory Power of the Regression Model
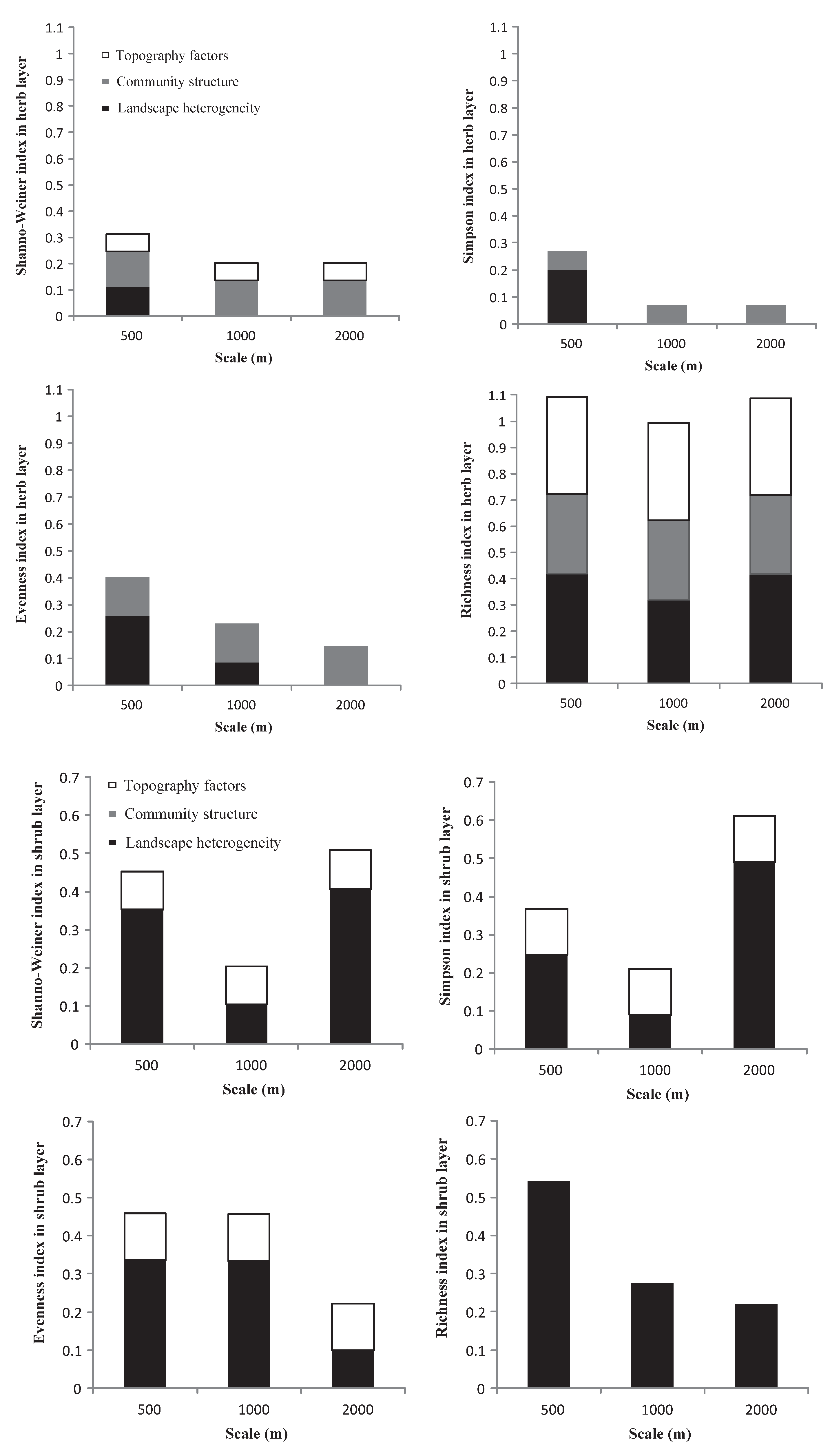
3.5. The Relative Importance of the Effects of Landscape Heterogeneity, Community Structure, and Topographic Factors on Diversity Indices and Richness Index
4. Discussion
4.1. The Response of the Diversity Index to the Impact Factors
4.2. The Relative Importance of Impact Factors on Species Biodiversity Varying with Buffer Radius
4.3. The Impact of Landscape Heterogeneity on Plant Diversity
5. Conclusions
- (1)
- The response mechanisms of the plant richness index and diversity indices in different layers under different buffer radii to impact factors were different. Compared to the biodiversity indices commonly used in the past, such as the Shannon diversity index (H’), evenness index (E), and dominance index (D’), the richness index in the herb layer was more direct and sensitive than the richness index in the tree and shrub layers and the diversity indices in the three layers to the impact factors.
- (2)
- The combined explanatory power of landscape heterogeneity, community structure, and topographic factors accounted for 43% of the species diversity indices, and 62% of the richness index at its peak.
- (3)
- The three impact factors that affect the species diversity indices and richness index of urban remnant vegetation rarely act alone, and often cause comprehensive cumulative effects and scale dependence.
- (4)
- Scale does matter in urbanization landscape studies. At a 500 m buffer radius, community structure combined with road disturbance indices was strongly related to diversity indices in herb and shrub layers. The stand age was negatively correlated with the tree layer richness index. As the scale increased, the diversity indices and richness index in the three layers decreased or increased under the influence of comprehensive factors.
- (5)
- Except for the herb layer, the interpretation of landscape heterogeneity for each plant diversity index was more stable than that for the other two factors. Road disturbance indices (AD and RD), farthest distance from the sample point to the forest edge (FD), area-edge indices (LPI), edge indices (ED), shape indices (SHAPE), and landscape diversity indices (SHDI and SIDI), a total of 8 indices, can well indicate species diversity and richness.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mckinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Rocha-Santos, L.; Pessoa, M.S.; Cassano, C.R.; Cassano, C.R.; Talora, D.C.; Orihuela, R.L.L.; Mariano-NetobJosé, E.; Morante-Filho, J.C.; Faria, D.; Cazetta, E. The shrinkage of a forest: Landscape-scale deforestation leading to overall changes in local forest structure. Biol. Conserv. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Li, D.; Stucky, B.J.; Deck, J.; Baiser, B.; Guralnick, R.P. The effect of urbanization on plant phenology depends on regional temperature. Nat. Ecol. Evol. 2019, 3, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Rusterholz, H.P.; Baur, B. Urbanisation and forest size affect the infestation rates of plant-galling arthropods and damage by herbivorous insects. Eur. J. Entomol. 2020, 117, 34–48. [Google Scholar] [CrossRef]
- Mao, Y.Y.; Chen, F.Q.; Guo, Y.; LiuM, J.X.; Jiang, F.; Li, X. Restoration dynamics of understory species diversity in Castanopsis hystrix plantations at different ages. Chin. J. Appl. Environ. Biol. 2021, 27, 930–937. [Google Scholar]
- Ye, Y.H.; Lin, S.S.; Wu, J.; Li, J.; Zou, J.F.; Yu, D.Y. Effect of rapid urbanization on plant species diversity in municipal parks, in a new Chinese city: Shenzhen. Acta Ecol. Sin. 2012, 32, 221–226. [Google Scholar] [CrossRef]
- Dos Santos, J.S.; Silva-Neto, C.M.; Silva, T.C.; Siqueira, K.N.; Ribeiro, M.C.; Collevatti, R.G. Landscape structure and local variables affect plant community diversity and structure in a Brazilian agricultural landscape. Biotropica 2021, 54, 239–250. [Google Scholar] [CrossRef]
- Nakagawa, M.; Momose, K.; Kishimoto, K.; Kamoi, T.; Tanaka, H.; Kaga, M.; Yamashita, S.; Itioka, T.; Nagamasu, H.; Sakai, S.; et al. Tree community structure, dynamics, and diversity partitioning in a Bornean tropical forested landscape. Biodivers. Conserv. 2012, 22, 127–140. [Google Scholar] [CrossRef]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef]
- Forman, R.T.T. Urban ecology principles: Are urban ecology and natural area ecology really different? Landsc. Ecol. 2016, 31, 1653–1662. [Google Scholar] [CrossRef]
- Huang, L.; Zhu, W.; Ren, H.; Chen, H.; Wang, J. Impact of atmospheric nitrogen deposition on soil properties and herb-layer diversity in remnant forests along an urban–rural gradient in Guangzhou, southern China. Plant Ecol. 2012, 213, 1187–1202. [Google Scholar] [CrossRef]
- UN: China’s Urban Population Will Increase by Another 255 Million by 2050. Available online: https://www.un.org/development/desa/zh/news/population/2018-world-urbanization-prospects.html (accessed on 22 October 2021).
- Stevens, M.H.H.; Carson, W.P. Resource quantity, not resource heterogeneity, maintains plant diversity. Ecol. Lett. 2002, 5, 420–426. [Google Scholar] [CrossRef]
- Pausas, J.G.; Carreras, J.; Ferré, A.; Font, X. Coarse-scale plant species richness in relation to environmental heterogeneity. J. Veg. Sci. 2003, 14, 661–668. [Google Scholar] [CrossRef]
- Dufour, A.; Gadallah, F.; Wagner, H.H.; Guisan, A.; Buttler, A. Plant species richness and environmental heterogeneity in a mountain landscape: Effects of variability and spatial configuration. Ecography 2006, 29, 573–584. [Google Scholar] [CrossRef]
- Malkinson, D.; Kopel, D.; Wittenberg, L. From rural-urban gradients to patch-matrix frameworks: Plant diversity patterns in urban landscapes. Landsc. Urban Plan. 2018, 169, 260–268. [Google Scholar] [CrossRef]
- Chávez, V.; Macdonald, S.E. The influence of canopy patch mosaics on understory plant community composition in boreal mixed wood forest. For. Ecol. Manag. 2010, 259, 1067–1075. [Google Scholar] [CrossRef]
- Redon, M.; Bergès, L.; Cordonnier, T.; Luque, S. Effects of increasing landscape heterogeneity on local plant species richness: How much is enough? Ecol. Evol. 2014, 29, 773–787. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, X.; Zhou, M.; Zhou, M.; Ai, D.; Wang, G.; Wang, Y.; Chu, C.; Lundholm, J. The effect of environmental heterogeneity on species richness depends on community position along the environmental gradient. Sci. Rep. 2015, 5, 15723. [Google Scholar] [CrossRef]
- Gastauer, M.; Mitre, S.K.; Carvalho, C.S.; Trevelin, L.C.; Sarmento, P.; Meira, N.J.; Caldeira, C.F.; Ramos, S.J.; Jaffé, R. Landscape heterogeneity and habitat amount drive plant diversity in Amazonian canga ecosystems. Landsc. Ecol. 2021, 36, 393–406. [Google Scholar] [CrossRef]
- Schindler, S.; Wehrden, H.V.; Poriazidis, K.; Wrbka, T.; Kati, V. Multiscale performance of landscape metrics as indicators of species richness of plants, insects and vertebrates. Ecol. Indic. 2013, 31, 41–48. [Google Scholar] [CrossRef]
- Priego-Santander, A.G.; Campos, M.; Bocco, G.; Ramirez-Sanchez, L.G. Relationship between landscape heterogeneity and plant species richness on the Mexican Pacific coast. Appl. Geogr. 2013, 40, 171–178. [Google Scholar] [CrossRef]
- Fan, M.; Wang, Q.H.; Mi, K.; Peng, Y. Scale-dependent effects of landscape pattern on plant diversity in Hunshandak Sandland. Biodivers. Conserv. 2017, 26, 2169–2185. [Google Scholar] [CrossRef]
- Kim, H.; Lee, C.B. On the relative importance of landscape variables to plant diversity and phylogenetic community structure on uninhabited islands, South Korea. Landsc. Ecol. 2020, 36, 209–211. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, W.T.; Lu, Y.T.; Dong, J.H.; Zhou, Y.Q.; Shang, J.X.; Li, X.; Mi, K. Multiscale influences of urbanized landscape metrics on the diversity of indigenous plant species: A case study in Shunyi District of Beijing, China. Chin. J. Appl. Environ. Biol. 2021, 31, 4058–4066. [Google Scholar]
- Joern, F.; David, B. Landscape Modification and Habitat Fragmentation:A Synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar]
- Peng, Y.; Fan, M.; Qing, F.T.; Xue, D.Y. Study Progresses on Effects of Landscape Metrics on Plant Diversity. Ecol. Environ. Conserv. 2016, 25, 1061–1068. [Google Scholar]
- Naidu, M.T.; Kumar, O.A. Tree diversity, stand structure, and community composition of tropical forests in Eastern Ghats of Andhra Pradesh, India. J. Asia-Pac. Biodivers. 2016, 9, 328–334. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G. Effect of tree species mixing on the size structure, density, and yield of forest stands. Eur. J. For. Res. 2016, 135, 1–22. [Google Scholar] [CrossRef]
- Walker, S.J.; Grimm, N.B.; Briggs, J.M.; Gries, C.; Dugan, L. Effects of urbanization on plant species diversity in central Arizona. Front. Ecol. Environ. 2009, 7, 465–470. [Google Scholar] [CrossRef]
- Zou, M.; Zhu, K.H.; Yin, J.Z.; Gu, B. Analysis on Slope Revegetation Diversity in Different Habitats. Procedia Earth Planet. Sci. Lett. 2012, 5, 180–187. [Google Scholar]
- Qian, S.H.; Qin, D.; Wu, X.; Hu, S.W.; Hu, L.Y.; Lin, D.M.; Zhao, L.; Shang, K.K.; Song, K.; Yang, Y.C. Urban growth and topographical factors shape patterns of spontaneous plant community diversity in a mountainous city in southwest China. Urban For. Urban Green. 2020, 55, 126814. [Google Scholar] [CrossRef]
- Arteaga, M.A.; Delgado, J.D.; Otto, R.; Fernández-Palacios, J.M.; Arévalo, J.R. How do alien plants distribute along roads on oceanic islands? A casestudy in Tenerife, Canary Islands. Biol. Invasions 2009, 11, 1071–1086. [Google Scholar] [CrossRef]
- Schlesinger, M.D.; Manley, P.N.; Holyoak, M. Distinguishing stressors acting on land bird communities in an urbanizing environment. Ecology 2008, 89, 2302–2314. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Ma, K.; Wu, J.; Tang, R.G.; Luo, S.H.; Zhang, Y.X.; Bao, L. Distribution pattern of allergenic plants in the Beijing metropolitan region. Aerobiologia 2013, 29, 217–231. [Google Scholar] [CrossRef]
- Krishnamoorthy, M.A.; Webala, P.W.; Kingston, T. Baobab fruiting is driven by scale-dependent mediation of plant size and landscape features. Landsc. Ecol. 2022, 37, 226–250. [Google Scholar] [CrossRef]
- Ryser, R.; Hirt, M.R.; Häussler, J.; Gravel, D.; Brose, U. Landscape heterogeneity buffers biodiversity of simulated meta-food-webs under global change through rescue and drainage effects. Nat. Commun. 2021, 12, 4716. [Google Scholar] [CrossRef]
- Sultana, M.; Müller, M.; Meyer, M.; Storch, I. Neighboring Green Network and Landscape Metrics Explain Biodiversity within Small Urban Green Areas-A Case Study on Birds. Sustainability 2022, 14, 6394. [Google Scholar] [CrossRef]
- Zeleny, D.; Li, C.F.; Chytry, M. Pattern of local plant species richness along a gradient of landscape topographical heterogeneity: Result of spatial mass effect or environmental shift? Ecography 2010, 33, 578–589. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Syrbe, R.U.; Michel, E.; Walz, U. Structural indicators for the assessment of biodiversity and their connection to the richness of avifauna. Ecol. Indic. 2013, 31, 89–98. [Google Scholar] [CrossRef]
- Amici, V.; Rocchini, D.; Filibeck, G.; Bacaro, G.; Santi, E.; Grei, F.; Landi, S.; Scoppola, A.; Chiarucci, A. Landscape structure effects on forest plant diversity at local scale: Exploring the role of spatial extent. Ecol. Complex. 2015, 21, 44–52. [Google Scholar] [CrossRef]
- Güneralp, B.; Perlstein, A.S.; Seto, K.C. Balancing urban growth and ecological conservation: A challenge for planning and governance in China. Ambio 2015, 44, 532–543. [Google Scholar] [CrossRef] [PubMed]
- China Statistics Press. Statistics Bureau of Guangzhou Municipality. In Guangzhou Statistical Yearbook; China Statistics Press: Beijing, China, 2018; pp. 6–214. [Google Scholar]
- Luo, Z. The Study of Plant Diversity of Green Space in Urban Built-Up Area of Guangzhou City. Master’s Thesis, Zhongkai University of Agriculture and Engineering, Guangzhou, China, 2013. [Google Scholar]
- Zhang, X.C.; Pan, Q. Research on spatial structure changes of urban land use. GIS and Remote Sensing in Hydrology. Water Environ. Res. 2004, 289, 141–150. [Google Scholar]
- Interpretation of Relevant Greening Statistical Data of Guangzhou Forestry and Garden Bureau in 2017. Available online: http://lyylj.gz.gov.cn/zwgk/sjfb/content/post_3038151.html (accessed on 10 April 2022).
- Tian, Y.; Jim, C.Y.; Liu, Y. Using a Spatial Interaction Model to Assess the Accessibility of District Parks in Hong Kong. Sustainability 2017, 9, 1924. [Google Scholar] [CrossRef]
- Li, G.; Li, J.S.; Guan, X.; Wu, X.P.; Zhao, Z.P. Shengwu Duoyangxing Jiance Jishu Shouce; China Environmental Science Press: Beijing, China, 2014; pp. 15–45. [Google Scholar]
- Wu, W.C.; Ning, J.Z. The morphological characteristics and classification of the main Asteraceae weeds in Guangzhou. J. South China Agric. Univ. 1993, 2, 32–46. [Google Scholar]
- Yu, S.H.; Mao, R.B.; Tang, W.; Ma, W. Study of the areal types of seed weeds in China. Southwest China J. Agric. Sci. 2008, 4, 1189–1192. [Google Scholar]
- Wang, J.H.; Wang, J.W.; Wang, D.X.; Christensen, S.; Zhang, Y.W.; Lu, D.Y.; Chen, K.R. An Eco-Field Simulation Analysis on Normal Patch Distribution of Annual Weed. Guizhou Sci. 2012, 30, 1–2. [Google Scholar]
- Zeng, X.L.; Liao, F.L.; Wen, G.R.; Li, W.D. Investigation of the distribution and growth of major energy grasses in Meizhou city. Guangdong Agric. Sci. 2008, 7, 25–28. [Google Scholar]
- Crist, T.O.; Veech, J.A. Aditive partitioning of rarefaction curves and species-area relationships: Unifying α-, β- and γ-diversity with sample size and habitat area. Ecol. Lett. 2006, 9, 923–932. [Google Scholar] [CrossRef]
- Turtureanu, P.D.; Palpurina, S.; Becker, T.; Dolnik, C.; Ruprecht, E.; Sutcliffe, L.M.E.; Szabó, A.; Dengler, J. Scale- and taxon-dependent biodiversity patterns of dry grassland vegetation in Transylvania. Agric. Ecosyst. Environ. 2014, 182, 15–24. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtssonet, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Hautier, Y.; Tilman, D.; Isbell, F.; Seabloom, E.W.; Borer, E.T.; Reich, P.B. Anthropogenic environmental changes affect ecosystem stability via biodiversity. Science 2015, 348, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Bender, D.J.; Contreras, T.A.; Fahrig, L. Habitat loss and population decline: A meta-analysis of the patch size effect. Ecology 1998, 79, 517–533. [Google Scholar] [CrossRef]
- Hanski, I.; Ovaskainen, O. The metapopulation capacity of a fragmented landscape. Nature 2000, 404, 755–758. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Lomba, A.; Bunce, R.G.H.; Jongman, R.H.G.; Moreira, F.; Honrado, J. Interactions between abiotic filters, landscape structure and species traits as determinants of dairy farmland plant diversity. Landsc. Urban Plan. 2011, 99, 248–258. [Google Scholar] [CrossRef]
- Talaga, S.; Petitclerc, F.; Jean-François, C.; Dézerald, O.; Leroy, C.; Cereghino, R.; Dejean, A. Environmental drivers of community diversity in a neotropical urban landscape: A multi-scale analysis. Landsc. Ecol. 2017, 32, 1805–1818. [Google Scholar] [CrossRef]
- Yongjiu, F.; Yang, L.; Xiaohua, T. Spatiotemporal variation of landscape patterns and their spatial determinants in Shanghai, China. Ecol. Indic. 2018, 87, 22–32. [Google Scholar]
- Stiles, A.; Scheiner, S.M. A multi-scale analysis of fragmentation effects on remnant plant species richness in Phoenix, Arizona. J. Biogeogr. 2010, 37, 1721–1729. [Google Scholar] [CrossRef]
- Dong, C.F.; Liang, G.F.; Ding, S.Y.; Lu, X.L.; Tang, Q.; Li, D.K. Multi-scale effects for landscape metrics and species diversity under the different disturbance: A case study of Gongyi City. Acta Ecol. Sin. 2014, 34, 3444–3451. [Google Scholar]
- Wang, Y.T.; Ding, S.Y.; Liang, G.F. Multi-scale effects analysis for landscape structure and biodiversity of seminatural habitats and cropland in a typical agricultural landscape. Prog. Phys. Geogr. 2014, 33, 1704–1716. [Google Scholar]
- Monteiro, A.T.; Fava, F.; Goncalves, J.; Huete, A.; Gusmeroli, F.; Parolo, G.; Spano, D.; Bocchi, S. Landscape context determinants to plant diversity in the permanent meadows of Southern European Alps. Biodivers. Conserv. 2013, 22, 937–958. [Google Scholar] [CrossRef]
- Siefert, A. Spatial patterns of functional divergence in old-field plant communities. Oikos 2012, 121, 907–914. [Google Scholar] [CrossRef]
- Reitalu, T.; Helm, A.; Pärtel, M.; Bengtsson, K.; Gerhold, P.; Rosén, E.; Takkis, K.; Znamenskiy, S.; Prentice, H.C. Determinants of fine-scale plant diversity in dry calcareous grasslands within the Baltic Sea region. Agric. Ecosyst. Environ. 2014, 182, 59–68. [Google Scholar] [CrossRef]
- Rocchini, D.; Delucchi, L.; Bacaro, G.; Cavallini, P.; Feilhauer, H.; Foody, G.M.; He, K.S.; Nagendra, H.; Porta, C.; Ricotta, C.; et al. Calculating landscape diversity with information-theory based indices:A GRASS GIS solution. Ecol. Inform. 2013, 17, 82–93. [Google Scholar] [CrossRef]
- Hernandez-Stefanoni, J.L. The Role of Landscape Patterns of Habitat Types on Plant Species Diversity of a Tropical Forest in Mexico. Biodivers. Conserv. 2006, 15, 1441–1457. [Google Scholar] [CrossRef]
- Krauss, J.; Steffan-Dewenter, I.; Tscharntke, T. Landscape occupancy and local population size depends on host plant distribution in the butterfly Cupido minimus. Biol. Conserv. 2004, 120, 355–361. [Google Scholar] [CrossRef]
- Saura, S.; Torras, O.; Gil-Tena, A.; Pascual-Hortal, L. Shape Irregularity as an Indicator of Forest Biodiversity and Guidelines for Metric Selection. In Patterns and Processes in Forest Landscapes; Lafortezza, R., Sanesi, G., Chen, J., Crow, T.R., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 167–189. [Google Scholar]
- Mitchell, M.S.; Rutzmoser, S.H.; Wigley, T.B.; Loehle, C.; Gerwin, J.A.; Keyser, P.D.; Lancia, R.A.; Perry, R.W.; Reynolds, C.J.; Thill, R.E.; et al. Relationships between avian richness and landscape structure at multiple scales using multiple landscapes. For. Ecol. Manag. 2006, 221, 155–169. [Google Scholar] [CrossRef]
- Wellstein, C.; Otte, A.; Waldhardt, R. Seed bank diversity in mesic grasslands in relation to vegetation type, management and site conditions. J. Veg. Sci. 2007, 18, 153–162. [Google Scholar] [CrossRef]
- Wrbka, T.; Schindler, S.; Pollheimer, M.; Schmitzberger, I.; Peterseil, J. Impact of the Austrian Agri-environmental scheme on diversity of landscapes, plants and birds. Community Ecol. 2008, 9, 217–227. [Google Scholar] [CrossRef]
- Honnay, O.; Piessens, K.; Van Landuyt, W.; Hermy, M.; Gulinck, H. Satellite based land use and landscape complexity indices as predictors for regional plant species diversity. Landsc. Urban Plan. 2003, 63, 241–250. [Google Scholar] [CrossRef]
- Janisova, M.; Michalcova, D.; Bacaro, G.; Ghisla, A. Landscape effects on diversity of semi-natural grasslands. Agric. Ecosyst. Environ. 2014, 182, 47–58. [Google Scholar] [CrossRef]
- Wang, R.X.; Yang, X.J. Nestedness theory suggests wetland fragments with large areas and macrophyte diversity benefit waterbirds. Ecol. Evol. 2021, 11, 12651–12664. [Google Scholar] [CrossRef] [PubMed]
- Horak, J.; Peltanova, A.; Podavkova, A.; Safarova, L.; Bogusch, P.; Romportl, D.; Zasadil, P. Biodiversity responses to land use in traditional fruit orchards of a rural agricultural landscape. Agric. Ecosyst. Environ. 2013, 178, 71–77. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Isaacs, R. Larger patches of diverse floral resources increase insect pollinator density, diversity, and their pollination of native wildflowers. Basic Appl. Ecol. 2014, 15, 701–711. [Google Scholar] [CrossRef]
- Matthies, S.A.; Ruter, S.; Prasse, R.; Schaarschmidt, F. Factors driving the vascular plant species richness in urban green spaces: Using a multivariable approach. Landsc. Urban Plan. 2015, 134, 177–187. [Google Scholar] [CrossRef]
- Tulloch, A.I.T.; Barnes, M.D.; Ringma, J.; Fuller, R.A.; Watson, J.E.M. Understanding the importance of small patches of habitat for conservation. J. Appl. Ecol. 2016, 53, 418–429. [Google Scholar] [CrossRef]
- Stejskalova, D.; Karasek, P.; Tlapakova, L.; Podhrazska, J. Sinuosity and Edge Effect-Important Factors of Landscape Pattern and Diversity. Pol. J. Environ. Stud. 2013, 22, 1177–1184. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Sample Sites | Longitude and Latitude | Altitude (m) | Slope (°) | Aspect (°) | Dominant Species | Age of Stand (Year) | Distance from the Road (m) |
|---|---|---|---|---|---|---|---|---|
| Grassland | JH | 113°17′09″, 23°14′31″ | 13 | - | - | Neyraudia reynaudiana | ≈10 | 50 |
| XG | 113°16′29″, 23°12′03″ | 11 | - | - | Neyraudia reynaudiana | ≈10 | 50 | |
| BY | 113°16′08″, 23°11′34″ | 20 | - | - | Neyraudia reynaudiana | ≈10 | 50 | |
| HSQ | 113°12′38″, 23°07′52″ | 9 | - | - | Neyraudia reynaudiana | ≈10 | 50 | |
| Secondary coniferous and broad leaved mixed forest | DS | 113°15′53″, 23°09′11″ | 59 | 20 | 280 | Pinus massoniana -Celtis sinensis -Ottochloa nodosa | 40–60 | 250 |
| YX | 113°15′28″, 23°08′32″ | 28 | 10 | 220 | Pinus massoniana- Cinnamomum burmanni -Piper sarmentosum | ≈60 | 50 | |
| TH | 113°21′42″, 23°07′50″ | 60 | 8 | 250 | Pinus massoniana +Psychotria rubra +Ottochloa nodosa | ≈60 | 40 | |
| Secondary broad leaved forest | BYST | 113°16′10″, 23°11′33″ | 180 | 45 | 13 | Schima superba -Psychotria rubra -Lophatherum gracile | 40–60 | 50 |
| BYSZ | 113°18′05″, 23°10′52″ | 255 | 3 | 60 | Schima superba -Psychotria rubra -Lophatherum gracile | 40–60 | 50 | |
| LYD | 113°21′12″, 23°12′29″ | 60 | 25 | 230 | Schima superba -Psychotria rubra -Lophatherum gracile | ≈60 | 300 | |
| PG | 113°21′45″, 23°11′26″ | 34 | 5 | 200 | Schima superba -Psychotria rubra -Lophatherum gracile | 60–80 | 50 | |
| HN | 113°21′15″, 23°09′30″ | 60 | 5 | 210 | Schima superba -Cinnamomum burmanni +Lophatherum gracile | 60–80 | 250 | |
| CL | 113°28′17″, 23°12′24″ | 58 | 20 | 290 | Castanea henryi -Castanea henryi -Cibotium barometz | 60–100 | 200 | |
| DL | 113°28′58″, 23°10′21″ | 50 | 2 | 100 | Schima superba -Psychotria rubra -Lophatherum gracile | >150 | 300 | |
| ZS | 113°29′06″, 23°08′27″ | 45 | 30 | 330 | Schima superba -Psychotria rubra -Adiantum capillus-veneris | 60–100 | 300 | |
| WC | 113°28′00″, 23°06′12″ | 60 | 30 | 210 | Schima superba -Rhaphiolepis indica -Dianella ensifolia | 60–100 | 150 |
| Types | Subtypes | Number of Indices | Indices Name |
|---|---|---|---|
| Landscape heterogeneity characteristics | Matrix indices | 1 | Vegetation coverage (Cv) |
| Road disturbance indices | 2 | Road density (RD) Average distance from sample point to road (AD) | |
| Distance from the edge of the forest | 2 | Shortest distance from sample point to forest edge (SD) Farthest distance from sample point to forest edge (FD) | |
| Clustering indices | 4 | Number of patches (NP) Patch density (PD) Largest patch index (LPI) Contagion index (CONTA) | |
| Area-edge indices | 1 | Landscape shape index (LSI) | |
| Edge indices | 2 | Edge density (ED) Total edge length (TE) | |
| Shape indices | 2 | Shape Index (SHAPE) Fractal dimension index (FRAC) | |
| Landscape diversity indices | 3 | Shannon’s diversity index (SHDI) Simpson’s diversity index (SIDI) Shannon’s Evenness index (SHE) | |
| Community structure | 4 | Coverage of the herb layer (Ch) Height of the herb layer (Hh) Coverage of the tree layer (Ct) Age of stand (Age) | |
| Topographic | 3 | Elevation, Slope, Aspect |
| Types of Index | Subtypes of Index | Abbreviations | Formula | Description |
|---|---|---|---|---|
| Species diversity index | Shannon-Wiener index | H’ | H’ = −∑PilnPi | Pi is the relative abundance of the ith species at each plot, ln is the natural log, and H describes the species richness and the equitability of individual distribution within species. |
| Simpson’s index | D | D = 1−∑Pi2 | Pi is the proportion of the individuals in species i, and D reflects the dominance in the community. | |
| Evenness | J | J = H’/H’max | H’ is Shannon-Wiener’s biodiversity index, and H’max is the maximum of H’. | |
| Species richness index | Patrick Richness | R | R = S | S is the number of species in the sample plot. |
| Vegetation Type | Code | Dominant Herb-Layer Species (Importance Value >5%) | Dominant Shrub-Layer Species (Importance Value >5%) | Dominant Tree-Layer Species (Importance Value >5%) |
|---|---|---|---|---|
| Urban weeds | I | Bidens Pilosa (24%) Neyraudia reynaudiana (22%) Themeda villosa (6%) | - | - |
| Secondary coniferous and broad-leaved mixed forest | II | Eriachne pallescens (51%) Pteris semipinnata (5%) | Psychotria rubra (25%) Cinnamomum burmanni (10%) Celtis sinensis (6%) Ilex asprella var. asprella (6%) Desmos chinensis (6%) Trema cannabina (5%) | Pinus massoniana (34%) Cinnamomum burmanni (11%) Celtis sinensis (10%) Cinnamomum camphora (6%) |
| Secondary broad-leaved forest | III | Lophatherum gracile (22%) Dicranopteris dichotoma (19%) Eriachne pallescens (8%) Adiantum capillus-veneris (7%) | Psychotria rubra (31%) Desmos chinensis (5%) | Schima superba (45%) |
| Layer | Biodiversity Indices | Buffer Radius/m | ||
|---|---|---|---|---|
| 500 | 1000 | 2000 | ||
| Herb | Shannon index | -Ch *, -AD | -Ch * | -Ch * |
| Simpson index | AD *, -PD, Ch | Ch, RD | Ch | |
| Evenness | -AD *, -SLOPE *, PD *, LPI | -RD | -Ct *, Ch | |
| Richness | -SHAPE *, Ct * | SLOPE *, ELEVATION *, -LPI *, -ED | SLOPE, -SHAPE *, Ct *, SHDI *, AD | |
| Shrub | Shannon index | -RD *, Ct | -NP | -LPI *, -LSI *, PD |
| Simpson index | RD *, -Ct *, FD | -SLOPE *, -Ct | -SLOPE *, -Ct | |
| Evenness | Ct *, -FD, SLOPE | Cv *, -SHDI *, -FD | Ct *, SLOPE | |
| Richness | -RD *, CONTAG | AD *, -FD | -FD * | |
| Tree | Shannon index | Ct | Ct *, -PD *, -AD * | AD,Ct |
| Simpson index | -Ct | PD *, AD, -Ct | -AD, -CONTAG | |
| Evenness | - | -FD, -NP | -SIDI, AD | |
| Richness | -AGE *, NP | -AGE* | -AGE *, AD *, Cv | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Yang, G.; Que, Q.; Wang, Q.; Zhang, Z.; Huang, L. How Do Landscape Heterogeneity, Community Structure, and Topographical Factors Contribute to the Plant Diversity of Urban Remnant Vegetation at Different Scales? Int. J. Environ. Res. Public Health 2022, 19, 14302. https://doi.org/10.3390/ijerph192114302
Liu X, Yang G, Que Q, Wang Q, Zhang Z, Huang L. How Do Landscape Heterogeneity, Community Structure, and Topographical Factors Contribute to the Plant Diversity of Urban Remnant Vegetation at Different Scales? International Journal of Environmental Research and Public Health. 2022; 19(21):14302. https://doi.org/10.3390/ijerph192114302
Chicago/Turabian StyleLiu, Xingzhao, Guimei Yang, Qingmin Que, Qi Wang, Zengke Zhang, and Liujing Huang. 2022. "How Do Landscape Heterogeneity, Community Structure, and Topographical Factors Contribute to the Plant Diversity of Urban Remnant Vegetation at Different Scales?" International Journal of Environmental Research and Public Health 19, no. 21: 14302. https://doi.org/10.3390/ijerph192114302
APA StyleLiu, X., Yang, G., Que, Q., Wang, Q., Zhang, Z., & Huang, L. (2022). How Do Landscape Heterogeneity, Community Structure, and Topographical Factors Contribute to the Plant Diversity of Urban Remnant Vegetation at Different Scales? International Journal of Environmental Research and Public Health, 19(21), 14302. https://doi.org/10.3390/ijerph192114302