
Comparative Analysis of Bacillariophyceae Chloroplast Genomes Uncovers Extensive Genome Rearrangements Associated with Speciation



Abstract
:1. Introduction
2. Materials and Methods
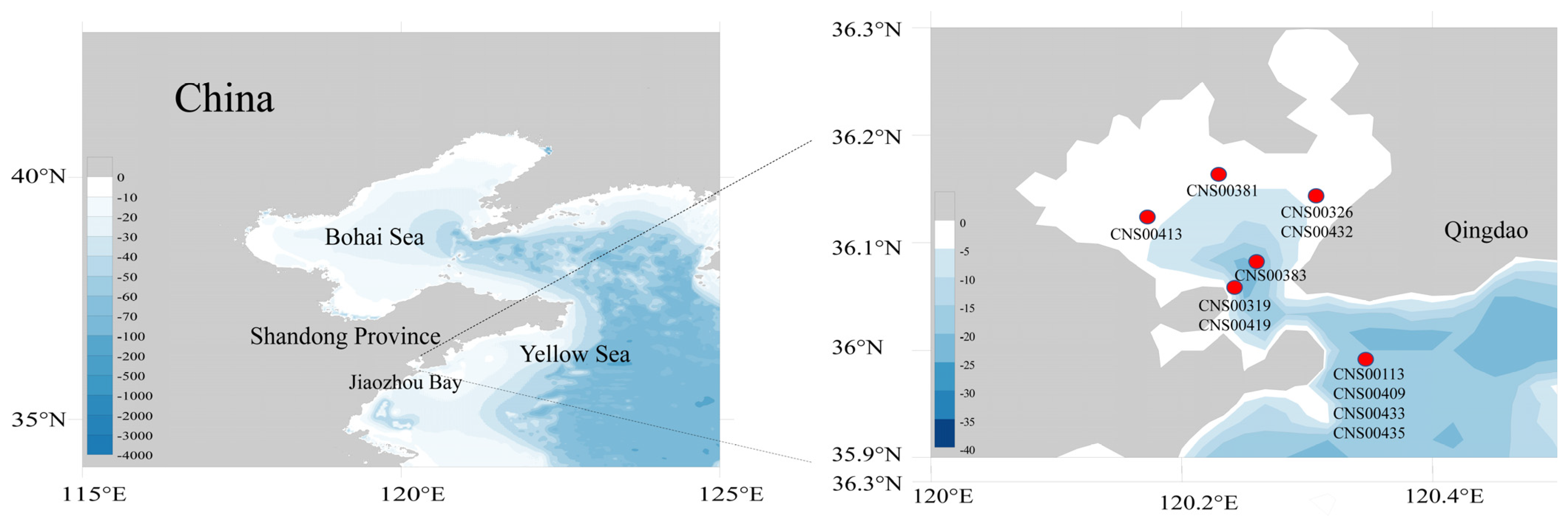
2.1. Strain Isolation and Culturing
2.2. DNA Library Preparation and Whole Genome Sequencing
2.3. Construction of Molecular Marker
2.4. Construction and Annotation of cpDNAs
2.5. Phylogenetic Analysis of cpDNAs
2.6. Synteny Analysis
2.7. Divergence Time Estimation
3. Results
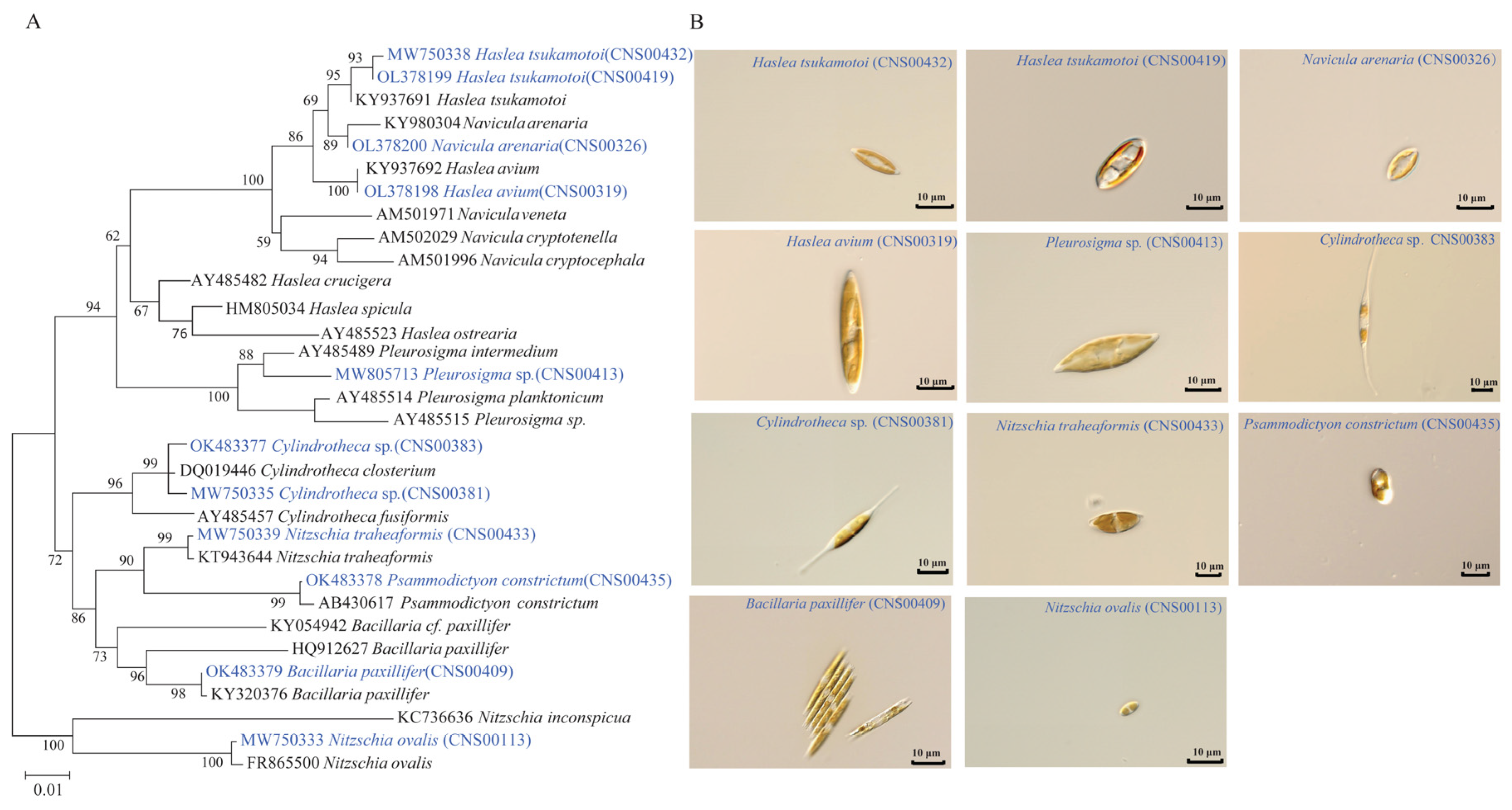
3.1. Molecular and Morphological Characterization of 11 Diatom Strains
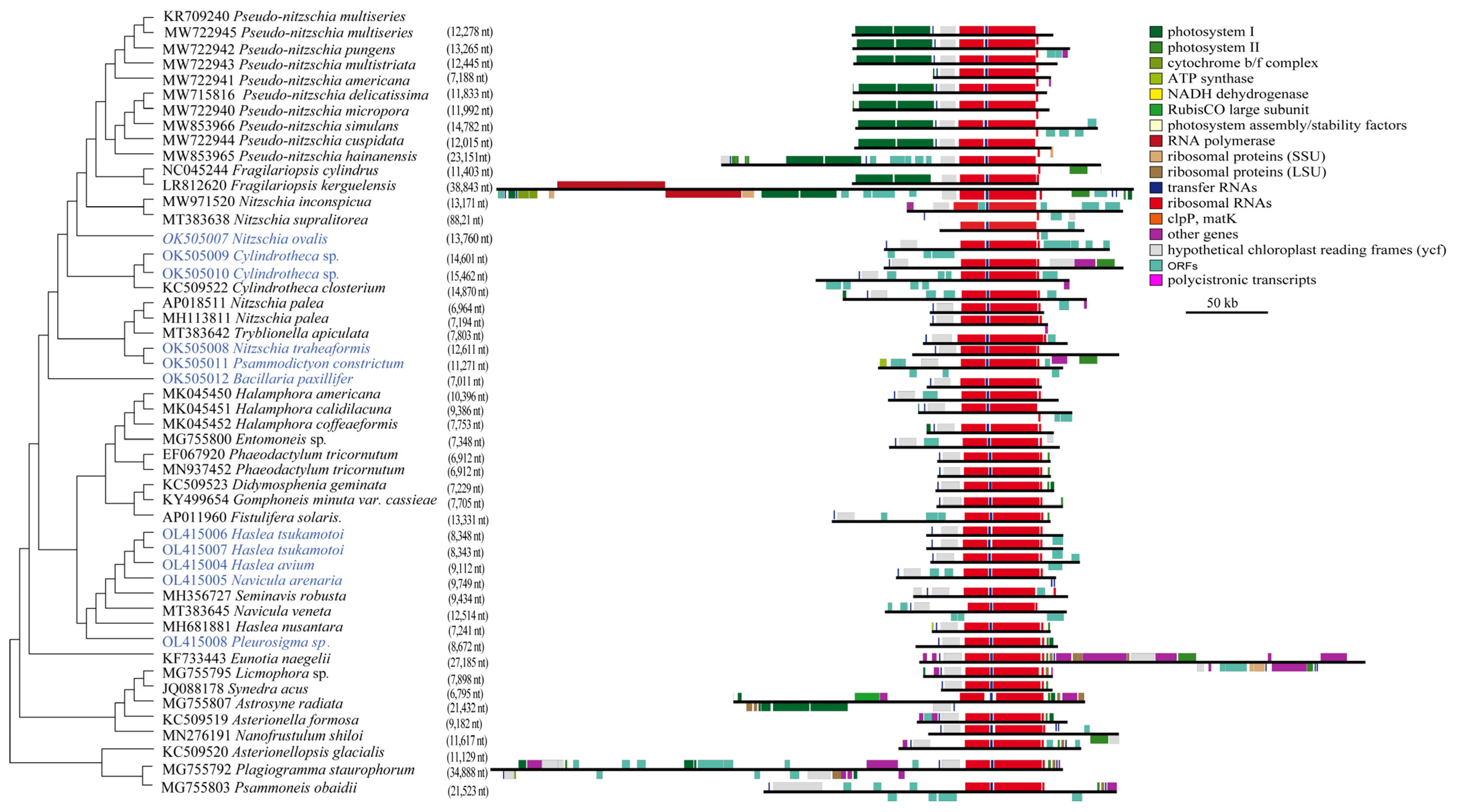
3.2. Construction and Comparative Analysis of cpDNAs
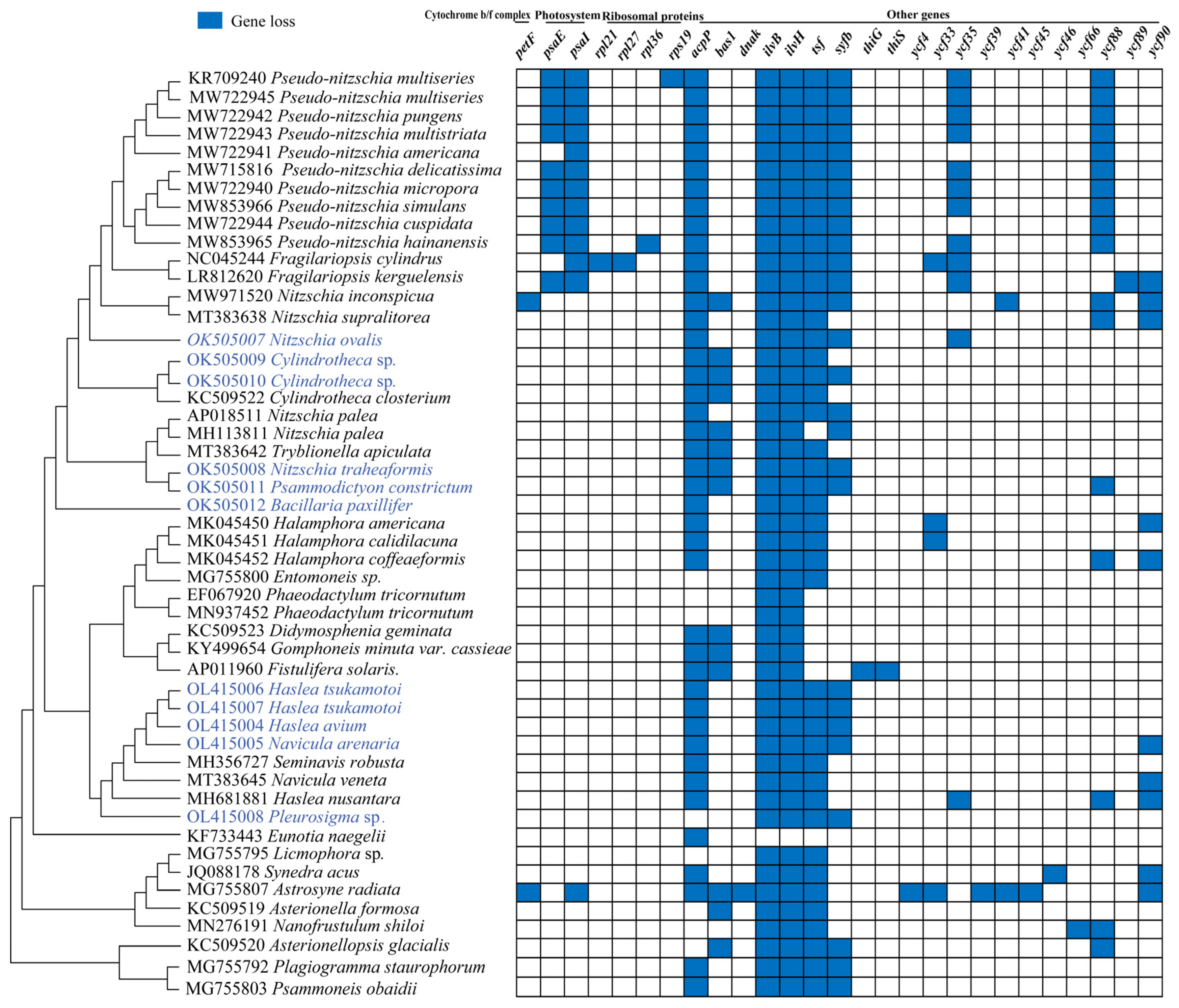
3.3. Plasticity of the Inverted Repeat Regions in Bacillariophyceae
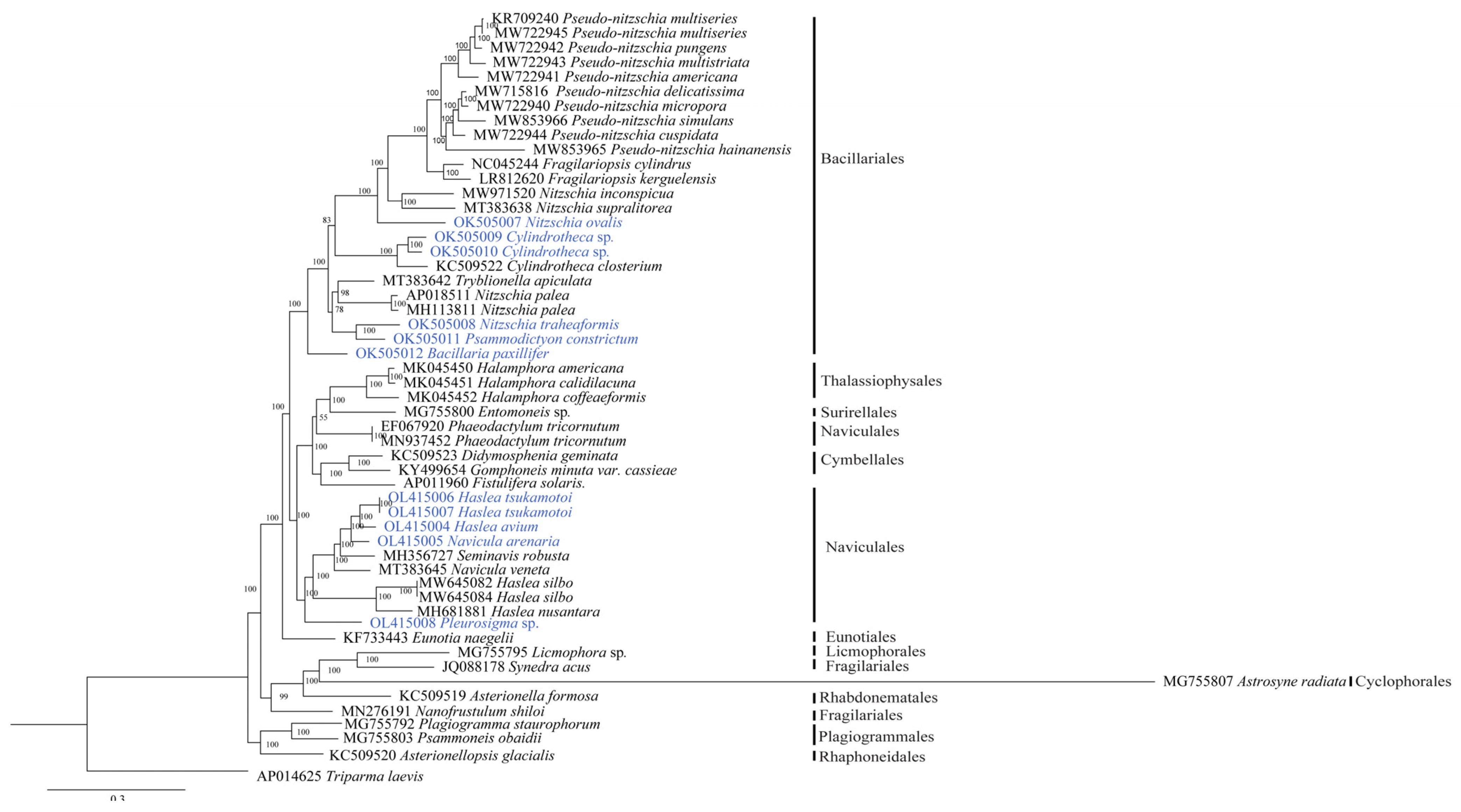
3.4. Phylogenetic Analysis of the Bacillariophyceae
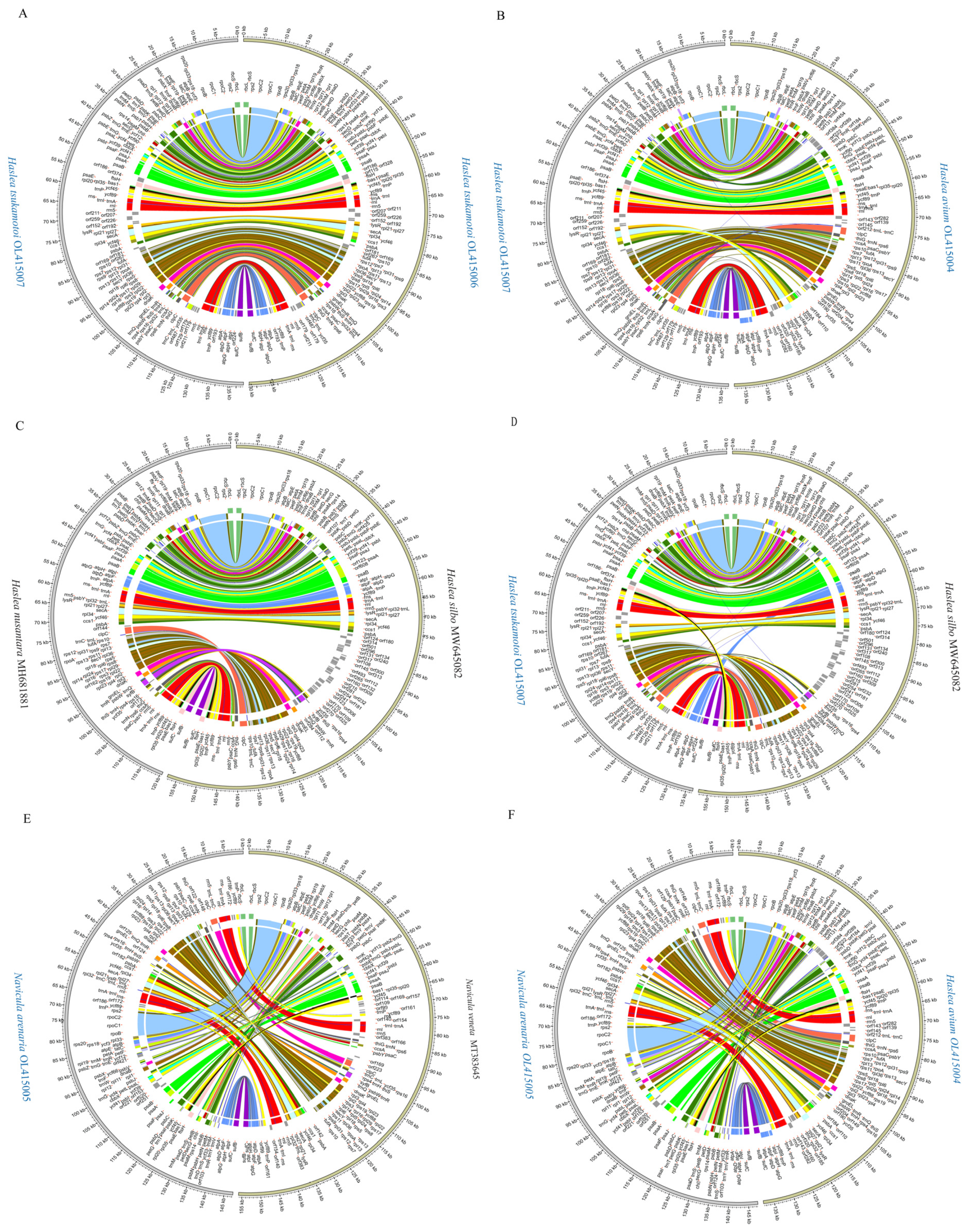
3.5. Inter- and Intra-Genus Synteny Analysis of cpDNAs
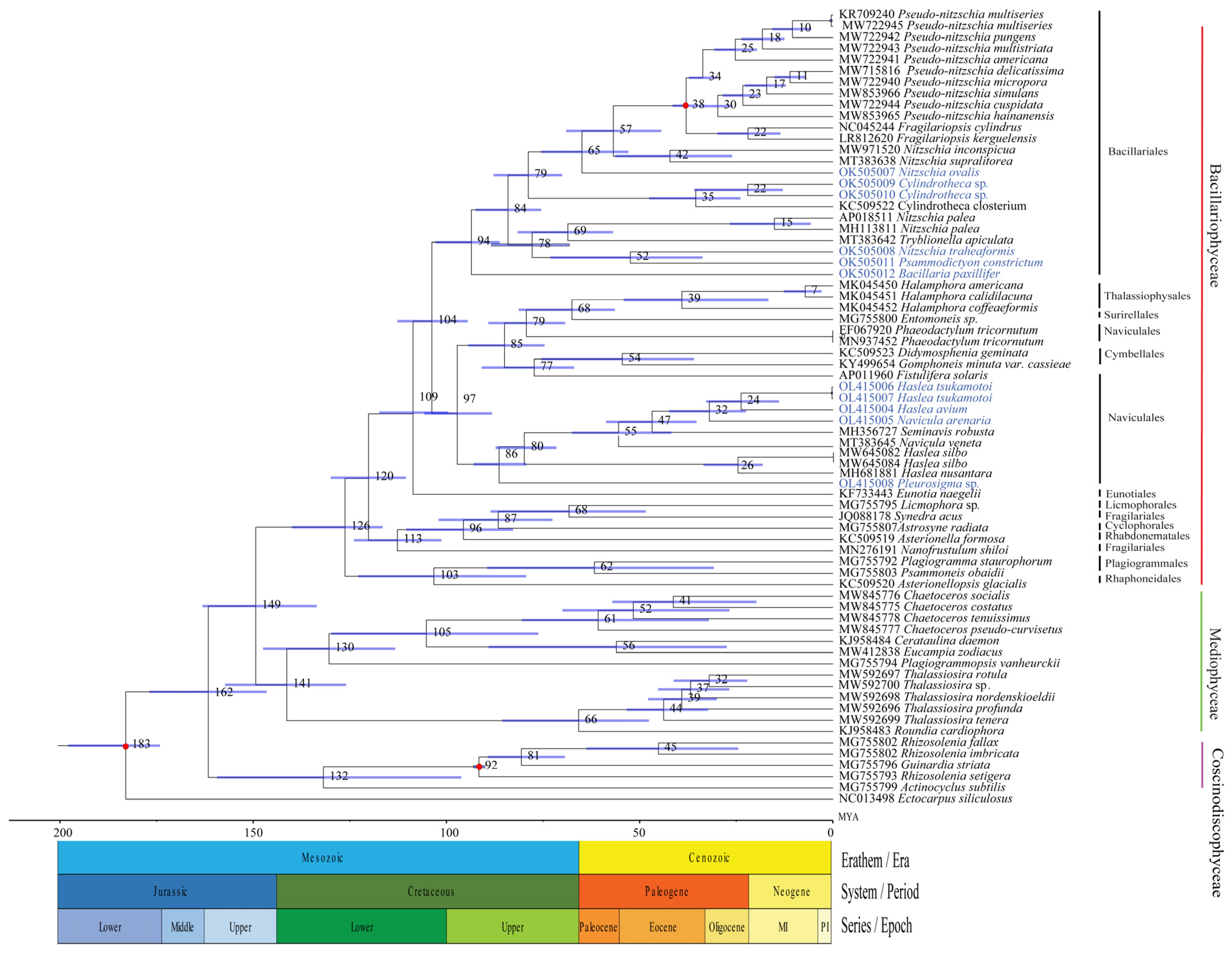
3.6. Divergence Time Estimation of Bacillariophyceae Species
4. Discussion
4.1. Chloroplast Genomes Are Useful Super Barcodes
4.2. Gene Loss and Plasticity of the IR Regions
4.3. Extensive Genome Arrangements between Bacillariophyceae Species
4.4. The Origin and Speciation of Bacillariophyceae Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smetacek, V. Biological oceanography-Diatoms and the silicate factor. Nature 1998, 391, 224–225. [Google Scholar] [CrossRef]
- Armbrust, E.V. The life of diatoms in the world’s oceans. Nature 2009, 459, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Falciatore, A.; Jaubert, M.; Bouly, J.P.; Bailleul, B.; Mock, T. Diatom Molecular Research Comes of Age: Model Species for Studying Phytoplankton Biology and Diversity (OPEN). Plant Cell 2020, 32, 547–572. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef]
- Karlusich, J.J.P.; Ibarbalz, F.M.; Bowler, C. Phytoplankton in the Tara Ocean. Ann. Rev. Mar. Sci. 2020, 12, 233–265. [Google Scholar] [CrossRef]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef]
- Nelson, D.M.; Treguer, P.; Brzezinski, M.A.; Leynaert, A.; Queguiner, B. Production and dissolution of biogenic silica in the ocean: Revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycles 1995, 9, 359–372. [Google Scholar] [CrossRef]
- Mann, D.G.; Crawford, R.M.; Round, F.E. Bacillariophyta. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 205–266. [Google Scholar] [CrossRef]
- Mann, D.G.; Trobajo, R.; Sato, S.; Li, C.; Witkowski, A.; Rimet, F.; Ashworth, M.P.; Hollands, R.M.; Theriot, E.C. Ripe for reassessment: A synthesis of available molecular data for the speciose diatom family Bacillariaceae. Mol. Phylogenetics Evol. 2021, 158, 106985. [Google Scholar] [CrossRef]
- Abdel Hamid, M.; Refaay, D.; Abdel-Mogib, M.; Azab, Y. Studies on biomass and lipid production of seven diatom species with special emphasis on lipid composition of Nitzschia palea (Bacillariophyceae) as reliable biodiesel feedstock. Algol. Stud. 2013, 143, 65–87. [Google Scholar] [CrossRef]
- Touliabah, H.E.; Abdel-Hamid, M.I.; Almutairi, A.W. Long-term monitoring of the biomass and production of lipids by Nitzschia palea for biodiesel production. Saudi. J. Biol. Sci. 2020, 27, 2038–2046. [Google Scholar] [CrossRef]
- Guasch, H.; Leira, M.; Montuelle, B.; Geiszinger, A.; Roulier, J.L.; Tornés, E.; Serra, A. Use of multivariate analyses to investigate the contribution of metal pollution to diatom species composition: Search for the most appropriate cases and explanatory variables. Hydrobiologia 2009, 627, 143–158. [Google Scholar] [CrossRef]
- Rimet, F. Recent views on river pollution and diatoms. Hydrobiologia 2012, 683, 1–24. [Google Scholar] [CrossRef]
- Tlili, A.; Corcoll, N.; Bonet, B.; Morin, S.; Montuelle, B.; Bérard, A.; Guasch, H. In situ spatio-temporal changes in pollution-induced community tolerance to zinc in autotrophic and heterotrophic biofilm communities. Ecotoxicology 2011, 20, 1823. [Google Scholar] [CrossRef]
- Zielinski, U.; Gersonde, R. Diatom distribution in Southern Ocean surface sediments (Atlantic sector): Implications for paleoenvironmental reconstructions. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 129, 213–250. [Google Scholar] [CrossRef]
- Lelong, A.; Hegaret, H.; Soudant, P.; Bates, S.S. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: Revisiting previous paradigms. Phycologia 2012, 51, 168–216. [Google Scholar] [CrossRef]
- Saeed, A.F.; Awan, S.A.; Ling, S.; Wang, R.; Wang, S. Domoic acid: Attributes, exposure risks, innovative detection techniques and therapeutics. Algal Res. 2017, 24, 97–110. [Google Scholar] [CrossRef]
- Gallardo, W.G.; Buen, S.M.A. Evaluation of mucus, Navicula, and mixed diatoms as larval settlement inducers for the tropical abalone Haliotis asinina. Aquaculture 2003, 221, 357–364. [Google Scholar] [CrossRef]
- Alallaf, A.; Kotab, M.; Shafik, H.; Elsayed, A. In vitro efficacy of biologically active compounds derived from Navicula arenaria against soil borne phytopathogenic Macrophomina phaseolina and Fusarium oxysporum. Alfarama J. Basic Appl. Sci. 2021, 2, 285–296. [Google Scholar] [CrossRef]
- Prasetiya, F.S.; Gastineau, R.; Poulin, M.; Lemieux, C.; Turmel, M.; Syakti, A.D.; Hardivillier, Y.; Widowati, I.; Risjani, Y.; Iskandar, I.; et al. Haslea nusantara (Bacillariophyceae), a new blue diatom from the Java Sea, Indonesia: Morphology, biometry and molecular characterization. Plant Ecol. Evol. 2019, 152, 188–202. [Google Scholar] [CrossRef]
- Pouvreau, J.-B.; Morançais, M.; Taran, F.; Rosa, P.; Dufossé, L.; Guérard, F.; Pin, S.; Fleurence, J.; Pondaven, P. Antioxidant and free radical scavenging properties of marennine, a blue-green polyphenolic pigment from the diatom Haslea ostrearia (Gaillon/Bory) Simonsen responsible for the natural greening of cultured oysters. J. Agric. Food Chem. 2008, 56, 6278–6286. [Google Scholar] [CrossRef]
- Gastineau, R.; Hardivillier, Y.; Leignel, V.; Tekaya, N.; Morançais, M.; Fleurence, J.; Davidovich, N.; Jacquette, B.; Gaudin, P.; Hellio, C.; et al. Greening effect on oysters and biological activities of the blue pigments produced by the diatom Haslea karadagensis (Naviculaceae). Aquaculture 2012, 368–369, 61–67. [Google Scholar] [CrossRef]
- Tanaka, T.; Maeda, Y.; Veluchamy, A.; Tanaka, M.; Abida, H.; Marechal, E.; Bowler, C.; Muto, M.; Sunaga, Y.; Tanaka, M.; et al. Oil Accumulation by the Oleaginous Diatom Fistulifera solaris as Revealed by the Genome and Transcriptome. Plant Cell 2015, 27, 162–176. [Google Scholar] [CrossRef]
- Trobajo, R.; Rovira, L.; Ector, L.; Wetzel, C.E.; Kelly, M.; Mann, D.G. Morphology and identity of some ecologically important small Nitzschia species. Diatom Res. 2013, 28, 37–59. [Google Scholar] [CrossRef]
- Carballeira, R.; Trobajo, R.; Leira, M.; Benito, X.; Sato, S.; Mann, D.G. A combined morphological and molecular approach to Nitzschia varelae sp. nov., with discussion of symmetry in Bacillariaceae. Eur. J. Phycol. 2017, 52, 342–359. [Google Scholar] [CrossRef]
- Rimet, F.; Kermarrec, L.; Bouchez, A.; Hoffmann, L.; Ector, L.; Medlin, L.K. Molecular phylogeny of the family Bacillariaceae based on 18S rDNA sequences: Focus on freshwater Nitzschia of the section Lanceolatae. Diatom Res. 2011, 26, 273–291. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: From gene to genome. Biol. Rev. Camb. Philos Soc. 2015, 90, 157–166. [Google Scholar] [CrossRef]
- Fu, C.N.; Wu, C.S.; Ye, L.J.; Mo, Z.Q.; Liu, J.; Chang, Y.W.; Li, D.Z.; Chaw, S.M.; Gao, L.M. Prevalence of isomeric plastomes and effectiveness of plastome super-barcodes in yews (Taxus) worldwide. Sci. Rep. 2019, 9, 2773. [Google Scholar] [CrossRef]
- Dong, W.; Xu, C.; Wen, J.; Zhou, S. Evolutionary directions of single nucleotide substitutions and structural mutations in the chloroplast genomes of the family Calycanthaceae. BMC Evol. Biol. 2020, 20, 96. [Google Scholar] [CrossRef]
- Yu, M.; Ashworth, M.P.; Hajrah, N.H.; Khiyami, M.A.; Sabir, M.J.; Alhebshi, A.M.; Al-Malki, A.L.; Sabir, J.S.M.; Theriot, E.C.; Jansen, R.K. Evolution of the Plastid Genomes in Diatoms. In Advances in Botanical Research; Plastid Genome Evolution; Elsevier: Amsterdam, The Netherlands, 2018; pp. 129–155. [Google Scholar]
- Song, H.; Liu, F.; Li, Z.; Xu, Q.; Chen, Y.; Yu, Z.; Chen, N. Development of a high-resolution molecular marker for tracking Phaeocystis globosa genetic diversity through comparative analysis of chloroplast genomes. Harmful Algae 2020, 99, 101911. [Google Scholar] [CrossRef]
- Cui, L.; Leebens-Mack, J.; Wang, L.-S.; Tang, J.; Rymarquis, L.; Stern, D.B.; dePamphilis, C.W. Adaptive evolution of chloroplast genome structure inferred using a parametric bootstrap approach. BMC Evol. Biol. 2006, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Hamsher, S.E.; Keepers, K.G.; Pogoda, C.S.; Stepanek, J.G.; Kane, N.C.; Kociolek, J.P. Extensive chloroplast genome rearrangement amongst three closely related Halamphora spp. (Bacillariophyceae), and evidence for rapid evolution as compared to land plants. PLoS ONE 2019, 14, e0217824. [Google Scholar] [CrossRef] [PubMed]
- Ruck, E.C.; Nakov, T.; Jansen, R.K.; Theriot, E.C.; Alverson, A.J. Serial gene losses and foreign DNA underlie size and sequence variation in the plastid genomes of diatoms. Genome Biol. Evol. 2014, 6, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Sabir, J.S.; Yu, M.; Ashworth, M.P.; Baeshen, N.A.; Baeshen, M.N.; Bahieldin, A.; Theriot, E.C.; Jansen, R.K. Conserved gene order and expanded inverted repeats characterize plastid genomes of Thalassiosirales. PLoS ONE 2014, 9, e107854. [Google Scholar] [CrossRef]
- Lommer, M.; Roy, A.S.; Schilhabel, M.; Schreiber, S.; Rosenstiel, P.; LaRoche, J. Recent transfer of an iron-regulated gene from the plastid to the nuclear genome in an oceanic diatom adapted to chronic iron limitation. BMC Genom. 2010, 11, 718. [Google Scholar] [CrossRef]
- Brembu, T.; Winge, P.; Tooming-Klunderud, A.; Nederbragt, A.J.; Jakobsen, K.S.; Bones, A.M. The chloroplast genome of the diatom Seminavis robusta: New features introduced through multiple mechanisms of horizontal gene transfer. Mar. Genom. 2014, 16, 17–27. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Wang, J.; Liu, F.; Chen, N. Mitochondrial genome of the harmful algal bloom species Odontella regia (Mediophyceae, Bacillariophyta). J. Appl. Phycol. 2021, 33, 855–868. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence-limits on phylogenies-an approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Capella-Gutierrez, S.; Silla-Martinez, J.M.; Gabaldon, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlic, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Bui Quang, M.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Lam-Tung, N.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Matari, N.H.; Blair, J.E. A multilocus timescale for oomycete evolution estimated under three distinct molecular clock models. BMC Evol. Biol. 2014, 14, 101. [Google Scholar] [CrossRef]
- Damste, J.S.S.; Muyzer, G.; Abbas, B.; Rampen, S.W.; Masse, G.; Allard, W.G.; Belt, S.T.; Robert, J.M.; Rowland, S.J.; Moldowan, J.M.; et al. The rise of the rhizosolenid diatoms. Science 2004, 304, 584–587. [Google Scholar] [CrossRef]
- Gachon, C.M.M.; Heesch, S.; Küpper, F.C.; Achilles-Day, U.E.M.; Brennan, D.; Campbell, C.N.; Clarke, A.; Dorrell, R.G.; Field, J.; Gontarek, S.; et al. The CCAP KnowledgeBase: Linking protistan and cyanobacterial biological resources with taxonomic and molecular data. Syst. Biodivers. 2013, 11, 407–413. [Google Scholar] [CrossRef]
- Witkowski, A.; Li, C.; Zglobicka, I.; Yu, S.-X.; Ashworth, M.; Bek, P.D.; Qin, S.; Tang, C.; Krzywda, M.; Ruppel, M.; et al. Multigene Assessment of Biodiversity of Diatom (Bacillariophyceae) Assemblages from the Littoral Zone of the Bohai and Yellow Seas in Yantai Region of Northeast China with some Remarks on Ubiquitous Taxa. J. Coast. Res. 2016, 74, 166–195. [Google Scholar] [CrossRef]
- Li, H.; Yang, G.; Sun, Y.; Wu, S.; Zhang, X. Cylindrotheca closterium is a species complex as was evidenced by the variations of rbcL gene and SSU rDNA. J. Ocean. Univ. China 2007, 6, 167–174. [Google Scholar] [CrossRef]
- Medlin, L.K.; Desdevises, Y. Phylogeny of ‘ARAPHID’ diatoms inferred from SSU & LSU rDNA, RBCL & PSBA sequences. Vie Et Milieu-Life Environ. 2016, 66, 129–154. [Google Scholar]
- An, S.M.; Choi, D.H.; Lee, J.H.; Lee, H.; Noh, J.H. Identification of benthic diatoms isolated from the eastern tidal flats of the Yellow Sea: Comparison between morphological and molecular approaches. PLoS ONE 2017, 12, e0179422. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; Sun, Z.; Xu, K. Taxonomy and molecular phylogeny of three marine benthic species of Haslea (Bacillariophyceae), with transfer of two species to Navicula. Diatom Res. 2017, 32, 451–463. [Google Scholar] [CrossRef]
- Cao, M.; Yuan, X.-L.; Bi, G. Complete sequence and analysis of plastid genomes of Pseudo-nitzschia multiseries (Bacillariophyta). Mitochondrial DNA Part A 2016, 27, 2897–2898. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Podell, S.; Pinowska, A.; Traller, J.C.; Smith, S.R.; McClure, R.; Beliaev, A.; Bohutskyi, P.; Hill, E.A.; Rabines, A.; et al. Diploid genomic architecture of Nitzschia inconspicua, an elite biomass production diatom. Sci. Rep. 2021, 11, 15592. [Google Scholar] [CrossRef] [PubMed]
- Gastineau, R.; Hamedi, C.; Baba Hamed, M.B.; Abi-Ayad, S.-M.E.-A.; Bąk, M.; Lemieux, C.; Turmel, M.; Dobosz, S.; Wróbel, R.J.; Kierzek, A.; et al. Morphological and molecular identification reveals that waters from an isolated oasis in Tamanrasset (extreme South of Algerian Sahara) are colonized by opportunistic and pollution-tolerant diatom species. Ecol. Indic. 2021, 121, 107104. [Google Scholar] [CrossRef]
- Guillory, W.X.; Onyshchenko, A.; Ruck, E.C.; Parks, M.; Nakov, T.; Wickett, N.J.; Alverson, A.J. Recurrent Loss, Horizontal Transfer, and the Obscure Origins of Mitochondrial Introns in Diatoms (Bacillariophyta). Genome Biol. Evol. 2018, 10, 1504–1515. [Google Scholar] [CrossRef]
- Mann, D.G.; Vanormelingen, P. An inordinate fondness? The number, distributions, and origins of diatom species. J. Eukaryot. Microbiol. 2013, 60, 414–420. [Google Scholar] [CrossRef]
- Amato, A.; Kooistra, W.H.C.F.; Ghiron, J.H.L.; Mann, D.G.; Proschold, T.; Montresor, M. Reproductive isolation among sympatric cryptic species in marine diatoms. Protist 2007, 158, 193–207. [Google Scholar] [CrossRef]
- Sarno, D.; Kooistra, W.; Medlin, L.K.; Percopo, I.; Zingone, A. Diversity in the genus Skeletonema (Bacillariophyceae). II. An assessment of the taxonomy of S-costatum-like species with the description of four new species. J. Phycol. 2005, 41, 151–176. [Google Scholar] [CrossRef]
- Xiong, A.S.; Peng, R.H.; Zhuang, J.; Gao, F.; Zhu, B.; Fu, X.Y.; Xue, Y.; Jin, X.F.; Tian, Y.S.; Zhao, W.; et al. Gene duplication, transfer, and evolution in the chloroplast genome. Biotechnol. Adv. 2009, 27, 340–347. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Mishra, A.K.; Khan, A.; Hashem, A.; Abd Allah, E.F.; Al-Harrasi, A. Gene Loss and Evolution of the Plastome. Genes 2020, 11, 1133. [Google Scholar] [CrossRef]
- Douglas, S.E.; Penny, S.L. The plastid genome of the cryptophyte alga, Guillardia theta: Complete sequence and conserved synteny groups confirm its common ancestry with red algae. J. Mol. Evol. 1999, 48, 236–244. [Google Scholar] [CrossRef]
- Glockner, G.; Rosenthal, A.; Valentin, K. The structure and gene repertoire of an ancient red algal plastid genome. J. Mol. Evol. 2000, 51, 382–390. [Google Scholar] [CrossRef]
- Puerta, M.V.S.; Bachvaroff, T.R.; Delwiche, C.F. The complete plastid genome sequence of the haptophyte Emiliania huxleyi: A comparison to other plastid genomes. DNA Res. 2005, 12, 151–156. [Google Scholar] [CrossRef]
- Bendich, A.J. Circular chloroplast chromosomes: The grand illusion. Plant Cell 2004, 16, 1661–1666. [Google Scholar] [CrossRef]
- Kolodner, R.; Tewari, K.K. Inverted repeats in chloroplast DNA from higher plants. Proc. Natl. Acad. Sci. USA 1979, 76, 41–45. [Google Scholar] [CrossRef]
- Zhu, A.; Guo, W.; Gupta, S.; Fan, W.; Mower, J.P. Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss on substitution rates. New Phytol. 2016, 209, 1747–1756. [Google Scholar] [CrossRef]
- Wu, C.-S.; Lai, Y.-T.; Lin, C.-P.; Wang, Y.-N.; Chaw, S.-M. Evolution of reduced and compact chloroplast genomes (cpDNAs) in gnetophytes: Selection toward a lower-cost strategy. Mol. Phylogenetics Evol. 2009, 52, 115–124. [Google Scholar] [CrossRef]
- Wolf, P.G.; Der, J.P.; Duffy, A.M.; Davidson, J.B.; Grusz, A.L.; Pryer, K.M. The evolution of chloroplast genes and genomes in ferns. Plant Mol. Biol. 2011, 76, 251–261. [Google Scholar] [CrossRef]
- Strauss, S.H.; Palmer, J.D.; Howe, G.T.; Doerksen, A.H. Chloroplast genomes of two conifers lack a large inverted repeat and are extensively rearranged. Proc. Natl. Acad. Sci. USA 1988, 85, 3898–3902. [Google Scholar] [CrossRef]
- Wolfe, K.H. The site of deletion of the inverted repeat in pea chloroplast DNA contains duplicated gene fragments. Curr. Genet. 1988, 13, 97–99. [Google Scholar] [CrossRef]
- Liu, S.; Xu, Q.; Liu, K.; Zhao, Y.; Chen, N. Chloroplast Genomes for Five Skeletonema Species: Comparative and Phylogenetic Analysis. Front. Plant Sci. 2021, 12, 2962. [Google Scholar] [CrossRef]
- Liu, K.; Chen, Y.; Cui, Z.; Liu, S.; Xu, Q.; Chen, N. Comparative Analysis of Chloroplast Genomes of Thalassiosira Species. Front. Mar. Sci. 2021, 8, 21. [Google Scholar] [CrossRef]
- Xu, Q.; Cui, Z.; Chen, N. Comparative Analysis of Chloroplast Genomes of Seven Chaetoceros Species Revealed Variation Hotspots and Speciation Time. Front. Microbiol. 2021, 12, 742554. [Google Scholar] [CrossRef]
- Vanormelingen, P.; Vanelslander, B.; Sato, S.; Gillard, J.; Trobajo, R.; Sabbe, K.; Vyverman, W. Heterothallic sexual reproduction in the model diatom Cylindrotheca. Eur. J. Phycol. 2013, 48, 93–105. [Google Scholar] [CrossRef]
- Stock, W.; Vanelslander, B.; Rudiger, F.; Sabbe, K.; Vyverman, W.; Karsten, U. Thermal Niche Differentiation in the Benthic Diatom Cylindrotheca closterium (Bacillariophyceae) Complex. Front. Microbiol. 2019, 10, 1395. [Google Scholar] [CrossRef]
- Gmelin, J.F. Caroli a Linn´e Systema Naturae Per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis, 13th ed.; Vermes Infusoria; Georg Emanuel Beer: Leipzig, Gremany, 1788; Volume 1, pp. 3021–3909. [Google Scholar]
- Gastineau, R.; Hansen, G.; Poulin, M.; Lemieux, C.; Turmel, M.; Bardeau, J.F.; Leignel, V.; Hardivillier, Y.; Morancais, M.; Fleurence, J.; et al. Haslea silbo, A Novel Cosmopolitan Species of Blue Diatoms. Biology 2021, 10, 328. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Sims, P.A.; Mann, D.G.; Medlin, L.K. Evolution of the diatoms: Insights from fossil, biological and molecular data. Phycologia 2006, 45, 361–402. [Google Scholar] [CrossRef]
- Lewitus, E.; Bittner, L.; Malviya, S.; Bowler, C.; Morlon, H. Clade-specific diversification dynamics of marine diatoms since the Jurassic. Nat. Ecol. Evol. 2018, 2, 1715–1723. [Google Scholar] [CrossRef]
- Nakov, T.; Beaulieu, J.M.; Alverson, A.J. Insights into global planktonic diatom diversity: The importance of comparisons between phylogenetically equivalent units that account for time. ISME J. 2018, 12, 2807–2810. [Google Scholar] [CrossRef]
- Nakov, T.; Beaulieu, J.M.; Alverson, A.J. Accelerated diversification is related to life history and locomotion in a hyperdiverse lineage of microbial eukaryotes (Diatoms, Bacillariophyta). New Phytol. 2018, 219, 462–473. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Katz, M.E.; Milligan, A.J.; Fennel, K.; Cramer, B.S.; Aubry, M.P.; Berner, R.A.; Novacek, M.J.; Zapol, W.M. The rise of oxygen over the past 205 million years and the evolution of large placental mammals. Science 2005, 309, 2202–2204. [Google Scholar] [CrossRef] [PubMed]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.G.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Katz, M.E.; Knoll, A.H.; Quigg, A.; Raven, J.A.; Schofield, O.; Taylor, F.J.R. The evolution of modern eukaryotic phytoplankton. Science 2004, 305, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Cermeno, P.; Falkowski, P.G.; Romero, O.E.; Schaller, M.F.; Vallina, S.M. Continental erosion and the Cenozoic rise of marine diatoms. Proc. Natl. Acad. Sci. USA 2015, 112, 4239–4244. [Google Scholar] [CrossRef]
- Barker, P.F. Scotia Sea regional tectonic evolution: Implications for mantle flow and palaeocirculation. Earth-Sci. Rev. 2001, 55, 1–39. [Google Scholar] [CrossRef]
- Scher, H.D.; Martin, E.E. Timing and climatic consequences of the opening of Drake Passage. Science 2006, 312, 428–430. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Genes |
|---|---|
| Photosystem I | psaA, psaB, psaC, psaD, psaF, psaM, psaJ, psaL |
| Photosystem II | psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbN, psbT, psbV, psbX, psbY, psbZ |
| Cytochrome b/f complex | petA, petB, petD, petG, petL, petM, petN |
| ATP synthase | atpA, atpB, atpD, atpE, atpF, atpG, atpH, atpI |
| RubisCO subunit | rbcL, rbcS |
| RNA polymerase | rpoA, rpoB, rpoC1, rpoC2 |
| Ribosomal proteins (SSU) | rps2, rps3, rps4, rps5, rps7, rps8, rps9, rps10, rps11, rps12, rps13, rps14, rps16, rps17, rps18, rps20 |
| Ribosomal proteins (LSU) | rpl1, rpl2, rpl3, rpl4, rpl5, rpl6, rpl11, rpl12, rpl13, rpl14, rpl16, rpl18, rpl20, rpl23, rpl24, rpl29, rpl31, rpl32, rpl33, rpl34, rpl35 |
| Other genes | cbbX, ccs1, ccsA, chlI, clpC, dnaB, dnaK, ftsH, groEL, lysR, secA, secG, secY, sufB, sufC, tatC, tufA, thiG, thiS, ycf3, ycf4, ycf12, ycf39, ycf41, ycf46, ycf66 |
| Family | Species | Accession Number | Size (bp) | LSC Length (bp) | SSC Length (bp) | IR Length (bp) | AT Content | Core Genes PCGs/rRNAs |
|---|---|---|---|---|---|---|---|---|
| Bacillariaceae | Nitzschia ovalis | OK505007 | 140,703 | 67,862 | 45,741 | 13,760 | 68.7% | 127/6 |
| Bacillariaceae | Nitzschia traheaformis | OK505008 | 166,822 | 73,101 | 68,919 | 12,611 | 69.4% | 129/6 |
| Bacillariaceae | Cylindrotheca sp. | OK505009 | 188,592 | 98,331 | 61,479 | 14,601 | 70.0% | 137/6 |
| Bacillariaceae | Cylindrotheca sp. | OK505010 | 178,349 | 81,389 | 66,456 | 15,462 | 69.9% | 134/6 |
| Bacillariaceae | Bacillaria paxillifer | OK505012 | 119,449 | 63,955 | 41,892 | 7011 | 70.0% | 129/6 |
| Bacillariaceae | Psammodictyon constrictum | OK505011 | 186,015 | 90,740 | 73,153 | 11,271 | 69.2% | 127/6 |
| Naviculaceae | Haslea tsukamotoi | OL415006 | 136,784 | 70,894 | 49,194 | 8348 | 68.6% | 127/6 |
| Naviculaceae | Haslea tsukamotoi | OL415007 | 136,746 | 70,012 | 50,048 | 8343 | 68.6% | 128/6 |
| Naviculaceae | Haslea avium | OL415004 | 137,691 | 70,273 | 49,194 | 9112 | 68.7% | 128/6 |
| Naviculaceae | Navicula arenaria | OL415005 | 147,331 | 80,134 | 47,699 | 9749 | 69.3% | 127/6 |
| Pleurosigma | Pleurosigma sp. | OL415008 | 174,382 | 96,638 | 60,400 | 8672 | 70.4% | 121/6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, J.; Chen, Y.; Liu, S.; Zhao, Y.; Chen, N. Comparative Analysis of Bacillariophyceae Chloroplast Genomes Uncovers Extensive Genome Rearrangements Associated with Speciation. Int. J. Environ. Res. Public Health 2022, 19, 10024. https://doi.org/10.3390/ijerph191610024
Wang Y, Wang J, Chen Y, Liu S, Zhao Y, Chen N. Comparative Analysis of Bacillariophyceae Chloroplast Genomes Uncovers Extensive Genome Rearrangements Associated with Speciation. International Journal of Environmental Research and Public Health. 2022; 19(16):10024. https://doi.org/10.3390/ijerph191610024
Chicago/Turabian StyleWang, Yichao, Jing Wang, Yang Chen, Shuya Liu, Yongfang Zhao, and Nansheng Chen. 2022. "Comparative Analysis of Bacillariophyceae Chloroplast Genomes Uncovers Extensive Genome Rearrangements Associated with Speciation" International Journal of Environmental Research and Public Health 19, no. 16: 10024. https://doi.org/10.3390/ijerph191610024
APA StyleWang, Y., Wang, J., Chen, Y., Liu, S., Zhao, Y., & Chen, N. (2022). Comparative Analysis of Bacillariophyceae Chloroplast Genomes Uncovers Extensive Genome Rearrangements Associated with Speciation. International Journal of Environmental Research and Public Health, 19(16), 10024. https://doi.org/10.3390/ijerph191610024