
Mycobacterium tuberculosis-Induced Maternal Immune Activation Promotes Autism-Like Phenotype in Infected Mice Offspring



Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Mtb Infection
2.3. Social Interaction Test
2.4. Self-Grooming/Repetitive Behaviors
2.5. Blood Collection
2.6. Brain Tissue Collection
2.7. Relative mRNA Expression in the Cerebellum (Quantitative PCR)
2.8. Cytokine Analysis
2.9. Statistical Analysis
3. Results
3.1. The Effect of Prenatal Mtb Infection on Offspring Social Behaviors
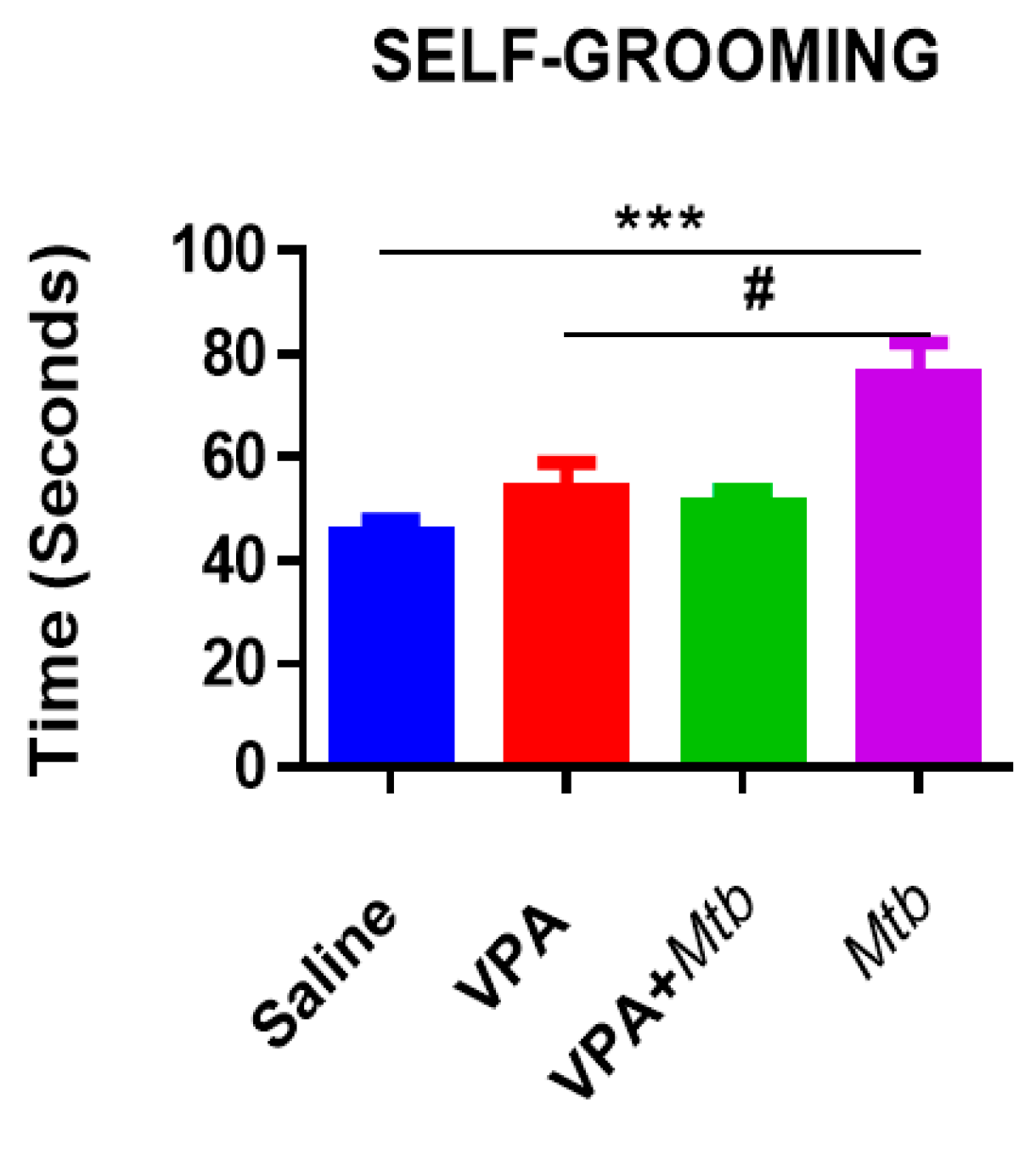
3.2. Mtb Infection Increases Self-Grooming Behavior
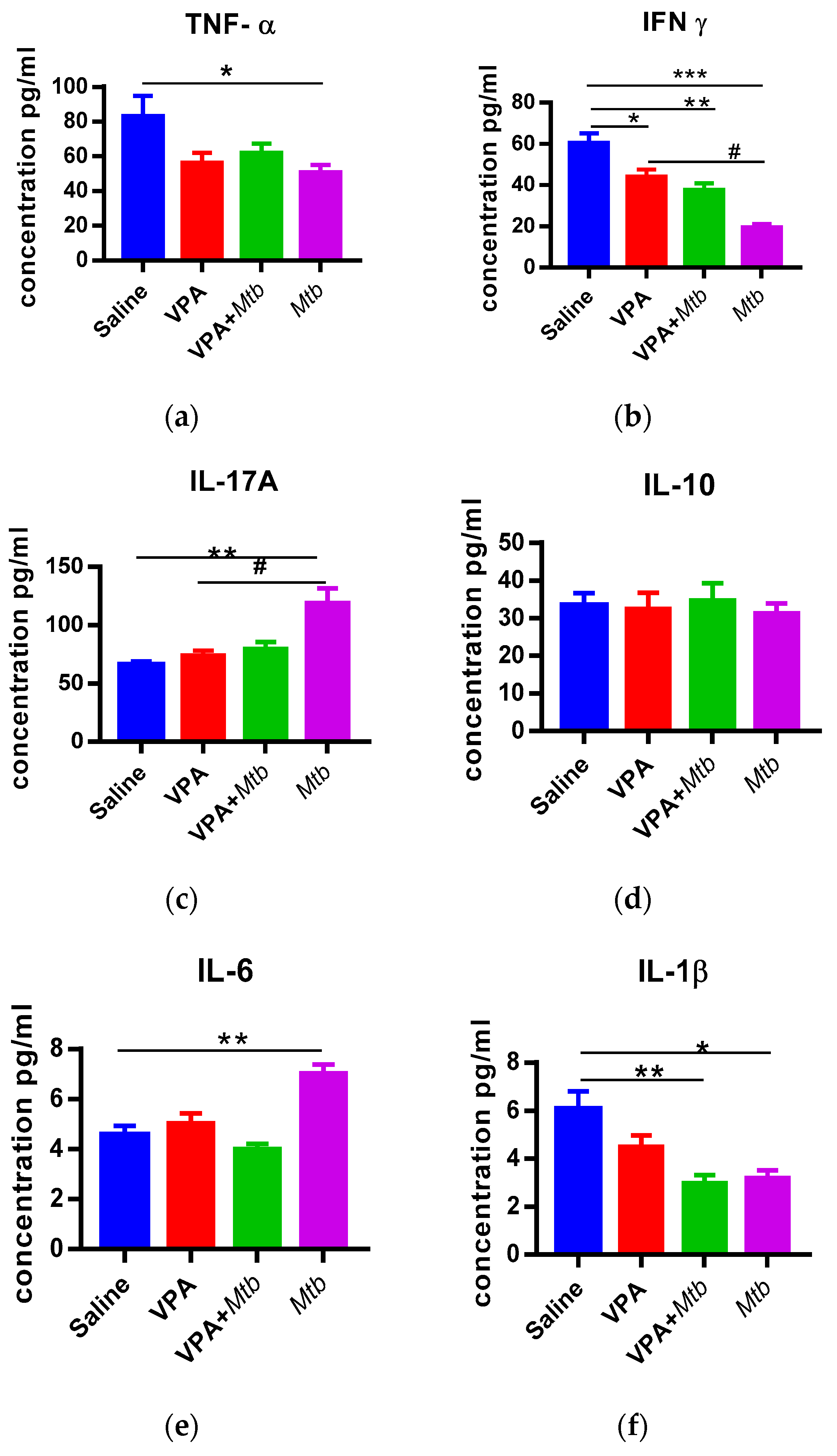
3.3. Prenatal Mtb Infection Causes Immune Dysregulation in Offspring
3.4. Mtb Infection Dysregulates Gene Expression of Synaptic Molecules in Offspring Cerebellum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | Mtb Concentration (pg/mL) | VPA + Mtb Concentration (pg/mL) |
|---|---|---|
| TNF-α | 50.7 + 11.21 * | 61.9 + 11.21 |
| IFN-γ | 19.6 + 4.78 *** | 37.7 + 4.78 |
| IL-17A | 119 + 10.35 a,# | 79.7 + 10.35 |
| Il-10 | 31.3 + 5.35 | 34.6 + 5.35 |
| Il-6 | 7.03 + 0.481 b,** | 4.00 + 0.481 |
| Il-1β | 3.20 + 0.701 * | 2.98 + 0.701 ** |
| Gene | Mtb Fold Change | VPA + Mtb Fold Change |
|---|---|---|
| NRXN1 | 4.31 + 0.971 *,a | 0.855 + 0.971 # |
| SHANK3 | 0.764 + 0.278 | 0.629 + 0.278 # |
| NLGN1 | 2.42 + 0.376 *,b | 1.35 + 0.376 |
| NRXN2 | 0.908 + 0.197 | 1.19 + 0.197 * |
| NLGN2 | 1.42 + 0.168 | 1.30 + 0.168 |
Appendix B
| ANOVA | DF | F (DFn,DFd) | p Value |
|---|---|---|---|
| Interaction | 3 | F (3, 88) = 14.12 | p < 0.0001 |
| Row FActor | 3 | F (3, 88) = 0.04163 | p = 0.9886 |
| Column Factor | 1 | F (1, 88) = 13.61 | p = 0.0004 |
| Residual | 88 |
| ANOVA | DF | F (DFn,DFd) | p Value |
|---|---|---|---|
| Interaction | 3 | F (3, 88) = 7.405 | p = 0.0002 |
| Row FActor | 3 | F (3, 88) = 4.065 | p = 0.0094 |
| Column Factor | 1 | F (1, 88) = 3.146 | P = 0,0796 |
| ANOVA | DF | F (DFn,DFd) | p Value |
|---|---|---|---|
| Treatment (between columns) | 3 | F (3, 44) = 11.29 | p < 0.0001 |
| CYTOKINES | DF | F (DFn,DFd) | p Value |
|---|---|---|---|
| TNF-α | 3 | F (3, 16) = 3.727 | p = 0.0332 |
| IFN-γ | 3 | F (3, 16) = 24.97 | p < 0.0001 |
| IL-17A | 3 | F (3, 16) = 10.47 | p = 0.0005 |
| IL-10 | 3 | F (3, 16) = 0.1383 | p = 0.9356 |
| Il-6 | 3 | F (3, 16) = 15.01 | p < 0.0001 |
| Il-1β | 3 | F (3, 16) = 8.232 | p = 0.0015 |
| GENE | DF | F (DFn,DFd) | p Value |
|---|---|---|---|
| NRXN1 | 3 | F (3, 16) = 5.51 | p = 0.0086 |
| SHANK3 | 3 | F (3, 16) = 3.258 | p = 0.0492 |
| NLGN1 | 3 | F (3, 16) = 7.215 | p = 0.0028 |
| NRXN2 | 3 | F (3, 16) = 3.29 | p = 0.0478 |
| NLGN2 | 3 | F (3, 16) = 1.013 | p = 0.4127 |
References
- Modabbernia, A.; Velthorst, E.; Reichenberg, A. Environmental risk factors for autism: An evidence-based review of systematic reviews and meta-analyses. Mol. Autism. 2017, 8, 13. [Google Scholar] [CrossRef]
- Brown, A.S.; Schaefer, C.A.; Wyatt, R.J.; Goetz, R.; Begg, M.D.; Gorman, J.M.; Susser, E.S. Maternal Exposure to Respiratory Infections and Adult Schizophrenia Spectrum Disorders: A Prospective Birth Cohort Study. Schizophr. Bull. 2000, 26, 287–295. [Google Scholar] [CrossRef]
- Fang, S.Y.; Wang, S.; Huang, N.; Yeh, H.H.; Chen, C.Y. Prenatal Infection and Autism Spectrum Disorders in Childhood: A Population-Based Case-Control Study in Taiwan. Paediatr. Perinat. Epidemiol. 2015, 29, 307–316. [Google Scholar] [CrossRef]
- Yasumatsu, K.; Nagao, J.I.; Arita-Morioka, K.I.; Narita, Y.; Tasaki, S.; Toyoda, K.; Ito, S.; Kido, H.; Tanaka, Y. Bacterial-induced maternal interleukin-17A pathway promotes autistic-like behaviors in mouse offspring. Exp. Anim. 2020, 69, 250–260. [Google Scholar] [CrossRef]
- Vargas, D.L.; Nascimbene, C.; Krishnan, C.; Zimmerman, A.W.; Pardo, C.A. Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann. Neurol. 2004, 57, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.; Van De Water, J. Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impaired behavioral outcome. Brain Behav. Immun. 2011, 25, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Urakubo, A.; Jarskog, L.; Lieberman, J.A.; Gilmore, J.H. Prenatal exposure to maternal infection alters cytokine expression in the placenta, amniotic fluid, and fetal brain. Schizophr. Res. 2001, 47, 27–36. [Google Scholar] [CrossRef]
- Offenbacher, S.; Riché, E.L.; Barros, S.P.; Bobetsis, Y.A.; Lin, D.; Beck, J.D. Effects of Maternal Campylobacter rectus Infection on Murine Placenta, Fetal and Neonatal Survival, and Brain Development. J. Periodontol. 2005, 76, 2133–2143. [Google Scholar] [CrossRef]
- Abram, M.; Schluter, D.; Vuckovic, D.; Wraber, B.; Doric, M.; Deckert, M. Murine model of pregnancy-associated Listeria monocytogenes infection. FEMS Immunol. Med. Microbiol. 2003, 35, 177–182. [Google Scholar] [CrossRef]
- Ashdown, H.; Dumont, Y.; Ng, M.; Poole, S.; Boksa, P.; Luheshi, G.N. The role of cytokines in mediating effects of prenatal infection on the fetus: Implications for schizophrenia. Mol. Psychiatry 2006, 11, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Huguet, G.; Bourgeron, T. Genetic Causes of Autism Spectrum Disorders; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 13–24. [Google Scholar]
- Wi, B.; Nowakowska, B.A. Genetics and epigenetics of autism spectrum disorder—current evidence in the field. J. Appl. Genet. 2019, 60, 37–47. [Google Scholar]
- Guang, S.; Pang, N.; Deng, X.; Yang, L.; He, F.; Wu, L.; Chen, C.; Yin, F.; Peng, J. Synaptopathology involved in autism spectrum disorder. Front. Cell. Neurosci. 2018, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Howard, L.; Gallagher, L.; Shen, S. Regulation and postsynaptic binding of neurexins—drug targets for neurodevelopmental and neuropsychiatric disorders. Front. Biol. 2015, 10, 239–251. [Google Scholar] [CrossRef]
- Lein, P.J. Overview of the Role of Environmental Factors in Neurodevelopmental Disorders. In Environmental Factors in Neurodevelopmental and Neurodegenerative Disorders; Academic Press: Cambridge, MA, USA, 2015; pp. 3–20. [Google Scholar]
- Kim, H.G.; Kishikawa, S.; Higgins, A.W.; Seong, I.S.; Donovan, D.J.; Shen, Y.; Lally, E.; Weiss, L.A.; Najm, J.; Kutsche, K.; et al. Disruption of neurexin 1 associated with autism spectrum disorder. Am. J. Hum. Genet. 2008, 82, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Almandil, N.B.; Alkuroud, D.N.; Abdulazeez, S.; Alsulaiman, A.; Elaissari, A.; Francis Borgio, J. Environmental and Genetic Factors in Autism Spectrum Disorders: Special Emphasis on Data from Arabian Studies. Int. J. Environ. Res. Public Health 2019, 16, 658. [Google Scholar] [CrossRef] [PubMed]
- Deckmann, I.; Brum Schwingel, G.; Fontes-Dutra, M.; Bambini-Junior, V.; Gottfried, C. Neuroimmune Alterations in Autism: A Translational Analysis Focusing on the Animal Model of Autism Induced by Prenatal Exposure to Valproic Acid Keywords Autism spectrum disorder Neuroimmune alterations Cytokines Animal model Valproic acid. Neuroimmunomodulation 2018, 25, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Roullet, F.I.; Lai, J.K.; Foster, J.A. In utero exposure to valproic acid and autism—A current review of clinical and animal studies. Neurotoxicol. Teratol. 2013, 36, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Bambini-junior, V.; Baronio, D.; Mackenzie, J.; Zanatta, G.; Riesgo, S. Comprehensive Guide to Autism; Springer: New York, NY, USA, 2014. [Google Scholar]
- Campolongo, M.; Kazlauskas, N.; Falasco, G.; Urrutia, L.; Salgueiro, N.; Höcht, C.; Depino, A.M. Sociability deficits after prenatal exposure to valproic acid are rescued by early social enrichment. Mol. Autism. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Peralta, F.; Fuentealba, C.; Fiedler, J.; Aliaga, E. Prenatal valproate treatment produces autistic-like behavior and increases metabotropic glutamate receptor 1A-immunoreactivity in the hippocampus of juvenile rats. Mol. Med. Rep. 2016, 14, 2807–2814. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Przewłocki, R. Behavioral alterations in rats prenatally to valproic acid: Animal model of autism. Neuropsychopharmacology 2005, 30, 80–89. [Google Scholar] [CrossRef]
- Kazlauskas, N.; Seiffe, A.; Campolongo, M.; Zappala, C.; Depino, A.M. Sex-specific effects of prenatal valproic acid exposure on sociability and neuroinflammation: Relevance for susceptibility and resilience in autism. Psychoneuroendocrinology 2019, 110, 104441. [Google Scholar] [CrossRef]
- Jana, N.; Vasishta, K.; Jindal, S.K.; Khunnu, B.; Ghosh, K. Perinatal outcome tuberculosis in pregnancies complicated by pulmonary. Int. J. Gynecol. Obstet. 1994, 44, 119–124. [Google Scholar] [CrossRef]
- Awodele, O.; Patrick, E.B.; Agbaje, E.O.; Oremosu, A.A.; Gbotolorun, S.C. Assessing the Risk of Birth Defects Associated with Exposure to Fixed-Dose Combined Antituberculous Agents during Pregnancy in Rats. Sci. World J. 2012, 2012, 1–8. [Google Scholar] [CrossRef][Green Version]
- Tilahun, D.; Fekadu, A.; Tekola, B.; Araya, M.; Roth, I.; Davey, B.; Hanlon, C.; Hoekstra, R.A. Ethiopian community health workers ’ beliefs and attitudes towards children with autism: Impact of a brief training intervention. Autism 2017, 23, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Kaidanovich-Beilin, O.; Lipina, T.; Vukobradovic, I.; Roder, J.; Woodgett, J.R. Assessment of social interaction behaviors. J. Vis. Exp. 2011, 48, e2473. [Google Scholar] [CrossRef] [PubMed]
- Malkova, N.V.; Yu, C.Z.; Hsiao, E.Y.; Moore, M.J.; Patterson, P.H. Maternal immune activation yields offspring displaying mouse versions of the three core symptoms of autism. Brain Behav. Immun. 2012, 26, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Schwartzer, J.J.; Careaga, M.; Onore, C.E.; Rushakoff, J.A.; Berman, R.F.; Ashwood, P. Maternal immune activation and strain specific interactions in the development of autism-like behaviors in mice. Transl. Psychiatry 2013, 3, e240-9. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The maternal interleukin-17a pathway in mice promotes autism-like phenotypes in offspring. Science 2016, 351, 933–939. [Google Scholar] [CrossRef]
- Smith, S.E.P.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 2007, 27, 10695–10702. [Google Scholar] [CrossRef]
- Garay, P.A.; Hsiao, E.Y.; Patterson, P.H.; McAllister, A.K. Maternal immune activation causes age- and region-specific changes in brain cytokines in offspring throughout development. Brain Behav. Immun. 2013, 31, 54–68. [Google Scholar] [CrossRef]
- Hsueh, P.T.; Lin, H.H.; Wang, H.H.; Liu, C.L.; Ni, W.F.; Liu, J.K.; Chang, H.H.; Sun, D.S.; Chen, Y.S.; Chen, Y.L. Immune imbalance of global gene expression, and cytokine, chemokine and selectin levels in the brains of offspring with social deficits via maternal immune activation. Genes Brain Behav. 2018, 17, e12479. [Google Scholar] [CrossRef] [PubMed]
- Rose, D.R.; Careaga, M.; Van de Water, J.; McAllister, K.; Bauman, M.D.; Ashwood, P. Long-term altered immune responses following fetal priming in a non-human primate model of maternal immune activation. Brain Behav. Immun. 2017, 63, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, M.; Gangestad, S.W. Rethinking IL-6 and CRP: Why they are more than inflammatory biomarkers, and why it matters. Brain Behav. Immun. 2018, 70, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Schindler, R.; Mancilla, J.; Endres, S.; Ghorbani, R.; Clark, S.C.; Dinarello, C.A. Correlations and interactions in the production of interleukin-6 (IL- 6), IL-1, and tumor necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood 1990, 75, 40–47. [Google Scholar] [CrossRef]
- Molloy, C.A.; Morrow, A.L.; Meinzen-Derr, J.; Schleifer, K.; Dienger, K.; Manning-Courtney, P.; Altaye, M.; Wills-Karp, M. Elevated cytokine levels in children with autism spectrum disorder. J. Neuroimmunol. 2006, 172, 198–205. [Google Scholar] [CrossRef]
- Jyonouchi, H. Immunological Abnormalities in Autism Spectrum Disorders. Adv. Neuroimmune Biol. 2013, 4, 141–159. [Google Scholar] [CrossRef]
- Ten Donkelaar, H.J.; Lammens, M.; Wesseling, P.; Thijssen, H.O.; Renier, W.O. Development and developmentaldisorders of the human cerebellum. J. Neurol. 2003, 250, 1025–1036. [Google Scholar] [CrossRef]
- Zhang, C.; Atasoy, D.; Araç, D.; Yang, X.; Fucillo, M.V.; Robison, A.J.; Ko, J.; Brunger, A.T.; Südhof, T.C. Neurexins physically and functionally interact with GABA(A) receptors. Neuron 2010, 66, 403–416. [Google Scholar] [CrossRef]
- Hines, R.M.; Wu, L.; Hines, D.J.; Steenland, H.; Mansour, S.; Dahlhaus, R.; Singaraja, R.R.; Cao, X.; Sammler, E.; Hormuzdi, S.G.; et al. Synaptic imbalance, stereotypies, and impaired social interactions in mice with altered neuroligin 2 expression. J. Neurosci. 2008, 28, 6055–6067. [Google Scholar] [CrossRef]
- Berridge, K.C.; Whishaw, I.Q. Cortex, striatum and cerebellum: Control of serial order in a grooming sequence. Exp. Brain Res. 1992, 90, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lim, C.S.; Kaang, B.K. Neuronal mechanisms and circuits underlying repetitive behaviors in mouse models of autism spectrum disorder. Behav. Brain Funct. 2016, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Sharova, L.; Sura, P.; Smith, B.J.; Gogal, R.M., Jr.; Sharov, A.A.; Ward, D.L.; Holladay, S.D. Nonspecific stimulation of the maternal immune system. II. Effects on gene expression in the fetus. Teratology 2000, 62, 420–428. [Google Scholar] [CrossRef]
- Holladay, S.D.; Sharova, L.; Smith, B.J.; Gogal, R.M.; Ward, D.L.; Blaylock, B.L. Nonspecific stimulation of the maternal immune system. I. Effects on teratogen-induced fetal malformations. Teratology 2000, 62, 413–419. [Google Scholar] [CrossRef]
- Holladay, S.D.; Sharova, L.V.; Punareewattana, K.; Hrubec, T.C.; Gogal, R.M., Jr.; Prater, M.R.; Sharov, A.A. Maternal immune stimulation in mice decreases fetal malformations caused by teratogens. Int. Immunopharmacol. 2002, 2, 325–332. [Google Scholar] [CrossRef]
- Hrubec, T.C.; Prater, M.R.; Mallela, M.K.; Gogal, R.M.; Guo, T.L.; Holladay, S.D. Diverse ability of maternal immune stimulation to reduce birth defects in mice exposed to teratogens: A review. J. Dev. Orig. Health Dis. 2012, 3, 132–139. [Google Scholar] [CrossRef]





| Group | Treatment and Dosage | Route of Administration |
|---|---|---|
| Saline | 0.2 mL of saline | Single i.p. |
| VPA | 0.2 mL of 500 mg/kg valproic acid | Single i.p. |
| VPA + Mtb | 0.2 mL of 350 mg/kg valproic acid + 10 mL of 1 × 108 CFU (OD600 = 1) | Single i.p. + aerosol infection |
| Mtb | 10 mL of 1 × 108 CFU (OD600 = 1) | Single aerosol infection |
| Genes | Forward Primer | Reverse Primer | Amplicon Size |
|---|---|---|---|
| NRXN1 | AGGTTCCGTGTGTCACTTGC | TCCTGTGTGTGTCTGGGGAT | 452 |
| NRXN2 | GCAGGGATTGGACACGCTAT | GAACTGTGACTGCCTACCCC | 464 |
| NLGN1 | GGGGATGAGGTTCCCTATGT | GGTTGGGTTTGGTATGGATG | 190 |
| NLGN2 | TTTCCGTCCTCCCCATCCAAT | TAGGAGCCGCCGTGTAGAAA | 923 |
| SHANK3 | GGCCATTTCAACAGAAGCCC | TGCGCCTTCGATCTCATGG | 119 |
| GAPDH | CCCTTAAGAGGGATGCTGCC | ACTGTGCCGTTGAATTTGCC | 118 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manjeese, W.; Mvubu, N.E.; Steyn, A.J.C.; Mpofana, T. Mycobacterium tuberculosis-Induced Maternal Immune Activation Promotes Autism-Like Phenotype in Infected Mice Offspring. Int. J. Environ. Res. Public Health 2021, 18, 4513. https://doi.org/10.3390/ijerph18094513
Manjeese W, Mvubu NE, Steyn AJC, Mpofana T. Mycobacterium tuberculosis-Induced Maternal Immune Activation Promotes Autism-Like Phenotype in Infected Mice Offspring. International Journal of Environmental Research and Public Health. 2021; 18(9):4513. https://doi.org/10.3390/ijerph18094513
Chicago/Turabian StyleManjeese, Wadzanai, Nontobeko E. Mvubu, Adrie J. C. Steyn, and Thabisile Mpofana. 2021. "Mycobacterium tuberculosis-Induced Maternal Immune Activation Promotes Autism-Like Phenotype in Infected Mice Offspring" International Journal of Environmental Research and Public Health 18, no. 9: 4513. https://doi.org/10.3390/ijerph18094513
APA StyleManjeese, W., Mvubu, N. E., Steyn, A. J. C., & Mpofana, T. (2021). Mycobacterium tuberculosis-Induced Maternal Immune Activation Promotes Autism-Like Phenotype in Infected Mice Offspring. International Journal of Environmental Research and Public Health, 18(9), 4513. https://doi.org/10.3390/ijerph18094513