
Differentiating Endurance-and Speed-Adapted Types of Elite and World Class Milers According to Biomechanical, Pacing and Perceptual Responses during a Sprint Interval Session


 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design
2.3. Measures
2.3.1. 100 m Sprint Time and Maximal Speed
2.3.2. Biomechanical Variables
2.3.3. Ratings of Perceived Exertion (RPE)
2.3.4. Core Affect
2.4. Statistical Analyses
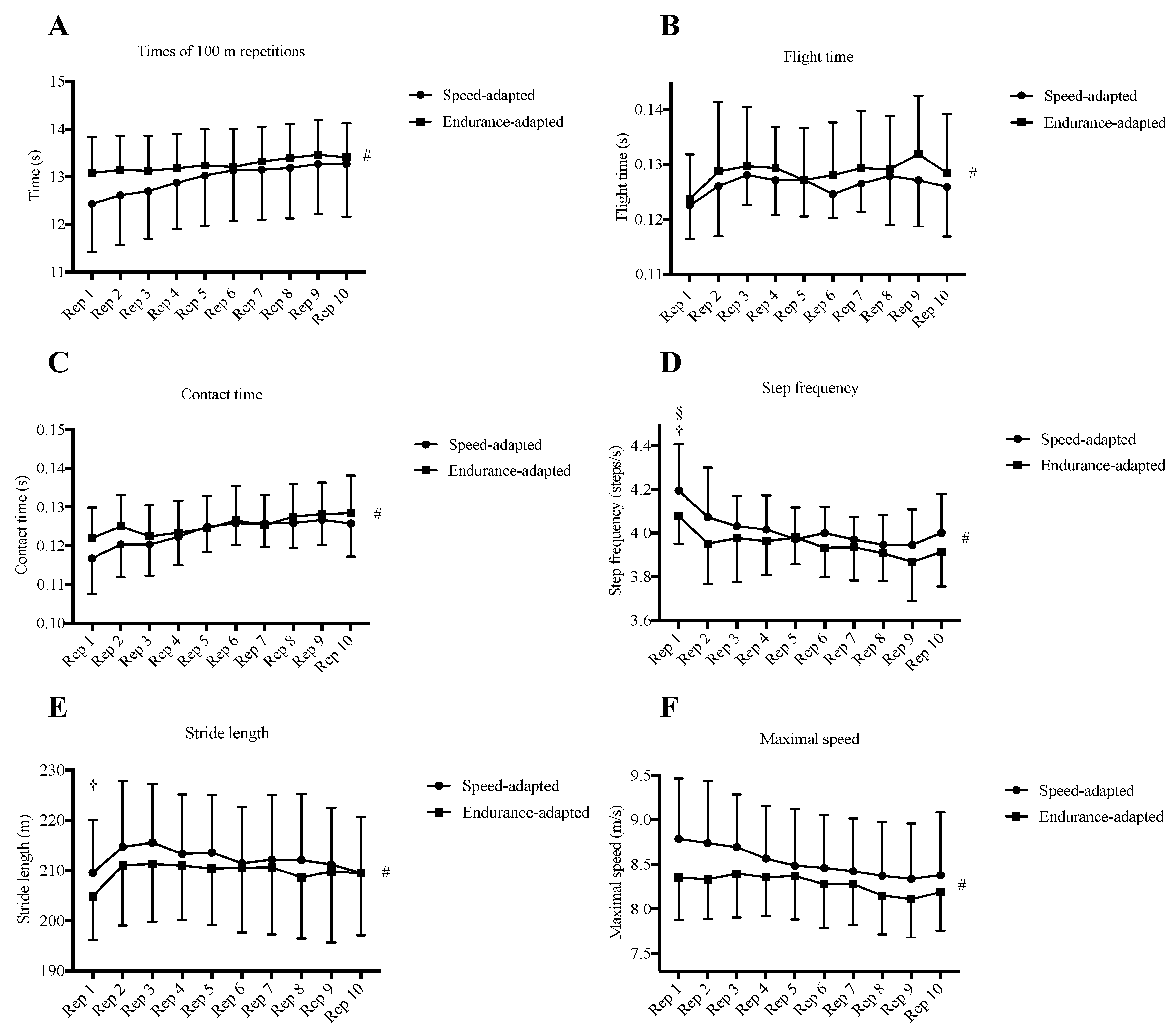
3. Results
4. Discussion
5. Conclusions and Practical Applications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Humphreys, J.; Holman, R. Focus on Middle Distance Running; Adam & Charles Black: London, UK, 1985. [Google Scholar]
- Spencer, M.R.; Gastin, P.B. Energy system contribution during 200- to 1500-m running in highly trained athletes. Med. Sci. Sports Exerc. 2001, 33, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Billat, L.V.; Koralsztein, J.P. Significance of the velocity at V̇O(2max) and time to exhaustion at this velocity. Sport Med. 1996, 22, 90–108. [Google Scholar] [CrossRef] [PubMed]
- Di Prampero, P.E.; Capelli, C.; Pagliaro, P.; Soule, R.G. Energetics of best performances in middle-distance running. J. Appl. Physiol. 1993, 74, 2318–2324. [Google Scholar] [CrossRef]
- Duffield, R.; Dawson, B. Energy system contribution in track running. New Stud. Athl. 2003, 18, 47–56. [Google Scholar]
- Kadono, H.; Enomoto, Y.; Ae, M. Change in the energetic of middle distance runners during race. J. Biomech. 2007, 40, S749. [Google Scholar] [CrossRef]
- Nummela, A.; Rusko, H. Time course of anaerobic and aerobic energy expenditure during short-term exhaustive running in athletes. Int. J. Sports Med. 1995, 16, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Hettinga, F.J.; Edwards, A.M.; Hanley, B. The Science Behind Competition and Winning in Athletics: Using World-Level Competition Data to Explore Pacing and Tactics. Front Sport Act. Living. 2019, 1. [Google Scholar] [CrossRef]
- Tucker, R.; Lambert, M.I.; Noakes, T.D. An analysis of pacing strategies during men’s world-record performances in track athletics. Int. J. Sports Physiol. Perform. 2006, 1, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, J.; Thorstensson, A. Ground reaction forces at different speeds of human walking and running. Acta Physiol. Scand. 1989, 136, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Cunninghan, R.; Hunter, I.; Seeley, M.; Feland, B. Variations in running technique between female sprinters, middle, and distance runners. Int. J. Exerc. 2013, 6, 43–51. [Google Scholar]
- Williams, K.R.; Cavanagh, P.R. Relationship between distance running mechanics, running economy, and performance. J. Appl. Physiol. 1987, 63, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Girard, O.; Brocherie, F.; Morin, J.B.; Degache, F.; Millet, G.P. Comparison of four sections for analyzing running mechanics alterations during repeated treadmill sprints. J. Appl. Biomech. 2015, 31, 389–395. [Google Scholar] [CrossRef]
- Venhorst, A.; Micklewright, D.P.; Noakes, T.D. The Psychophysiological Determinants of Pacing Behaviour and Performance During Prolonged Endurance Exercise: A Performance Level and Competition Outcome Comparison. Sport Med. 2018, 48, 2387–2400. [Google Scholar] [CrossRef]
- Renfree, A.; West, J.; Corbett, M.; Rhoden, C.; St Clair Gibson, A. Complex interplay between determinants of pacing and performance during 20-km cycle time trials. Int. J. Sports Physiol. Perform. 2012, 7, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Spiriev, A. IAAF Scoring Tables of Athletics: 2018 Revised Edition; IAAF: Monte Carlo, Monaco, 2019. [Google Scholar]
- Buchheit, M.; Laursen, P.B. High-intensity interval training, solutions to the programming puzzle: Part I: Cardiopulmonary emphasis. Sport Med. 2013, 43, 313–338. [Google Scholar] [CrossRef]
- Buchheit, M.; Laursen, P.B. High-intensity interval training, solutions to the programming puzzle: Part II: Anaerobic energy, neuromuscular load and practical applications. Sport Med. 2013, 43, 927–954. [Google Scholar] [CrossRef] [PubMed]
- Mero, A.; Komi, P.V.; Gregor, R.J. Biomechanics of Sprint Running: A Review. Sport Med. 1992, 13, 376–392. [Google Scholar] [CrossRef] [PubMed]
- Borg, G.A.V. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef]
- Pageaux, B. Perception of effort in Exercise Science: Definition, measurement and perspectives. Eur. J. Sport Sci. 2016, 16, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Hardy, C.J.; Rejeski, W.J. Not What, but How One Feels: The Measurement of Affect during Exercise. J. Sport Exerc. Psychol. 2016, 11, 304–317. [Google Scholar] [CrossRef]
- Svebak, S.; Murgatroyd, S. Metamotivational Dominance. A Multimethod Validation of Reversal Theory Constructs. J. Pers. Soc. Psychol. 1985, 48, 107–116. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioural Sciences/Jacob Cohen, 2nd ed.; Routeledge: Oxfordshire, UK, 1988. [Google Scholar]
- Hopkins, W.G.; Marshall, S.W.; Batterham, A.M.; Hanin, J. Progressive statistics for studies in sports medicine and exercise science. Med. Sci. Sports Exerc. 2009, 41, 3–12. [Google Scholar] [CrossRef]
- Casado, A.; Renfree, A. Fortune Favors the Brave. Tactical Behaviors in the Middle Distance Running Events at the 2017 IAAF World Championships. Int. J. Sports Physiol. Perform. 2018, 13, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Casado, A.; Hanley, B.; Jiménez-Reyes, P.; Renfree, A. Pacing profiles and tactical behaviors of elite runners. J. Sport Health Sci 2020. ahead of print. [Google Scholar] [CrossRef]
- Casado, A.; García-Manso, J.M.; Romero-Franco, N.; Martínez-Patiño, M.J. Pacing strategies during male 1500 m world record performances. Res. Sports Med. 2021. ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Brughelli, M.; Cronin, J.; Chaouachi, A. Effects of running velocity on running kinetics and kinematics. J. Strength Cond. Res. 2011, 25, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Brocherie, F.; Millet, G.P.; Girard, O. Neuro-mechanical and metabolic adjustments to the repeated anaerobic sprint test in professional football players. Eur. J. Appl. Physiol. 2015, 115, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Reyes, P.; Pareja-Blanco, F.; Cuadrado-Peñafiel, V.; Morcillo, J.A.; Párraga, J.A.; González-Badillo, J.J. Mechanical, Metabolic and Perceptual Response during Sprint Training. Int. J. Sports Med. 2016, 37, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Ingham, S.A.; Whyte, G.P.; Pedlar, C.; Bailey, D.M.; Dunman, N.; Nevill, A.M. Determinants of 800-m and 1500-m running performance using allometric models. Med. Sci. Sports Exerc. 2008, 40, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Girard, O.; Brocherie, F.; Tomazin, K.; Farooq, A.; Morin, J.B. Changes in running mechanics over 100-m, 200-m and 400-m treadmill sprints. J. Biomech. 2016, 49, 1490–1497. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Reyes, P.; Cross, M.; Ross, A.; Samozino, P.; Brughelli, M.; Gill, N.; Morin, J.-B. Changes in mechanical properties of sprinting during repeated sprint in elite rugby sevens athletes. Eur. J. Sport Sci. 2019, 19, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Watson, D. Mood and Temperament; The Guilford Press: New York, NY, USA, 2000. [Google Scholar]
- Casado, A.; Moreno-Pérez, D.; Larrosa, M.; Renfree, A. Different psychophysiological responses to a high-intensity repetition session performed alone or in a group by elite middle-distance runners. Eur. J. Sport Sci. 2019, 19, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, M.; Kraemer, R.; Bartholomew, J.; Acevedo, E.; Jarreau, D. Affective responses to exercise are dependent on intensity rather than total work. Med. Sci. Sports Exerc. 2007, 39, 1417–1422. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Performance, Biomechanical and Perceptual Variables | Mean ± SD | ES | p | 95% CI | |
|---|---|---|---|---|---|
| Speed-Adapted | Endurance-Adapted | ||||
| Performance (IAAF) | 977.8 ± 132.65 | 980.9 ± 110.68 | 0.03 | 0.86 | −118.14–111.94 |
| Other perform (IAAF) | 944.4 ± 164.98 | 1003.5 ± 127.6 | 0.4 | 0.38 | −197.67–79.47 |
| Times of 100 m rep (s) | 12.97 ± 1.03 | 13.26 ± 0.72 | 0.33 | 0.47 | −1.12–0.54 |
| Maximal speed (m/s) | 8.53 ± 0.62 | 8.28 ± 0.45 | 0.45 | 0.32 | −0.27–0.76 |
| Flight time (s) | 0.126 ± 0.006 | 0.129 ± 0.01 | 0.31 | 0.49 | −0.01–0.005 |
| Contact time (s) | 0.12 ± 0.007 | 0.13 ± 0.008 | 0.21 | 0.64 | −0.09–0.005 |
| Frequency (steps/s) | 4.02 ± 0.13 | 3.95 ± 0.14 | 0.45 | 0.32 | −0.07–0.19 |
| Stride length (m) | 212.28 ± 11.54 | 209.75 ± 11.79 | 0.22 | 0.63 | −8.43–13.48 |
| RPE | 16.14 ± 1.86 | 15.48 ± 1.85 | 0.36 | 0.44 | −1.08–2.4 |
| Valence | −1.12 ± 1.7 | 0.29 ± 1.96 | 0.77 | 0.09 | −3.13–0.31 |
| Felt arousal | 3.64 ± 0.87 | 3.37 ± 0.98 | 0.29 | 0.53 | −0.61–1.13 |
| Halves difference (s) | 2.36 ± 0.93 | 1.03 ± 1.48 | 1.07 | 0.03 | 0.17–2.49 |
| Performance, Biomechanical and Perceptual Variables | Mean ± SD | ES | p | 95% CI | |
|---|---|---|---|---|---|
| Speed-Adapted | Endurance-Adapted | ||||
| Times of 100 m rep (%) | 2.47 ± 0.74 | 1.64 ± 0.7 | 1.15 | 0.02 | 0.15–1.5 |
| Maximal speed (%) | 2.66 ± 0.44 | 2.45 ± 0.63 | 0.39 | 0.4 | −0.3–0.72 |
| Flight time (%) | 0.13 ± 0.006 | 0.13 ± 0.009 | 0.4 | 0.38 | −0.79–1.97 |
| Contact time (%) | 3.79 ± 0.82 | 2.66 ± 0.93 | 1.29 | 0.01 | 0.31–1.95 |
| Step frequency (%) | 3.79 ± 0.97 | 2.83 ± 1.35 | 0.82 | 0.08 | −0.14–2.06 |
| Stride length (%) | 2.29 ± 0.55 | 2 ± 0.62 | 0.5 | 0.28 | −0.26–0.85 |
| RPE (%) | 13.32 ± 3.56 | 12.1 ± 6.32 | 0.24 | 0.6 | −3.61–6.03 |
| Valence (%) | 67.03 ± 35.16 | 30 ± 20.67 | 1.28 | 0.01 | 9.93–64.13 |
| Felt arousal (%) | 28.35 ± 14.04 | 20.26 ± 14.43 | 0.57 | 0.22 | −5.29–21.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casado, A.; Renfree, A.; Jaenes-Sánchez, J.C.; Cuadrado-Peñafiel, V.; Jiménez-Reyes, P. Differentiating Endurance-and Speed-Adapted Types of Elite and World Class Milers According to Biomechanical, Pacing and Perceptual Responses during a Sprint Interval Session. Int. J. Environ. Res. Public Health 2021, 18, 2448. https://doi.org/10.3390/ijerph18052448
Casado A, Renfree A, Jaenes-Sánchez JC, Cuadrado-Peñafiel V, Jiménez-Reyes P. Differentiating Endurance-and Speed-Adapted Types of Elite and World Class Milers According to Biomechanical, Pacing and Perceptual Responses during a Sprint Interval Session. International Journal of Environmental Research and Public Health. 2021; 18(5):2448. https://doi.org/10.3390/ijerph18052448
Chicago/Turabian StyleCasado, Arturo, Andrew Renfree, José Carlos Jaenes-Sánchez, Víctor Cuadrado-Peñafiel, and Pedro Jiménez-Reyes. 2021. "Differentiating Endurance-and Speed-Adapted Types of Elite and World Class Milers According to Biomechanical, Pacing and Perceptual Responses during a Sprint Interval Session" International Journal of Environmental Research and Public Health 18, no. 5: 2448. https://doi.org/10.3390/ijerph18052448
APA StyleCasado, A., Renfree, A., Jaenes-Sánchez, J. C., Cuadrado-Peñafiel, V., & Jiménez-Reyes, P. (2021). Differentiating Endurance-and Speed-Adapted Types of Elite and World Class Milers According to Biomechanical, Pacing and Perceptual Responses during a Sprint Interval Session. International Journal of Environmental Research and Public Health, 18(5), 2448. https://doi.org/10.3390/ijerph18052448