
Developmental Neurotoxicity of Environmentally Relevant Pharmaceuticals and Mixtures Thereof in a Zebrafish Embryo Behavioural Test

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Maintenance of Fish and Egg Spawning
2.3. Zebrafish Embryotoxicity Test (ZFET)
2.4. Light-Dark Transition Test
2.5. Mixture Design
2.6. RNA Isolation and Quantitative Real-Time PCR
2.7. Statistical Analysis
2.7.1. ZFET—Single Compounds
2.7.2. Light-Dark Transition Test—Single Compounds
2.7.3. Gene Expression—Single Compounds
2.7.4. Mixtures and Light-Dark Transition Test
3. Results
3.1. Zebrafish Embryotoxicity Test (ZFET)
3.2. Behaviour Testing
3.2.1. Single-Compound Dose–Response Analysis
3.2.2. Different Exposure Windows
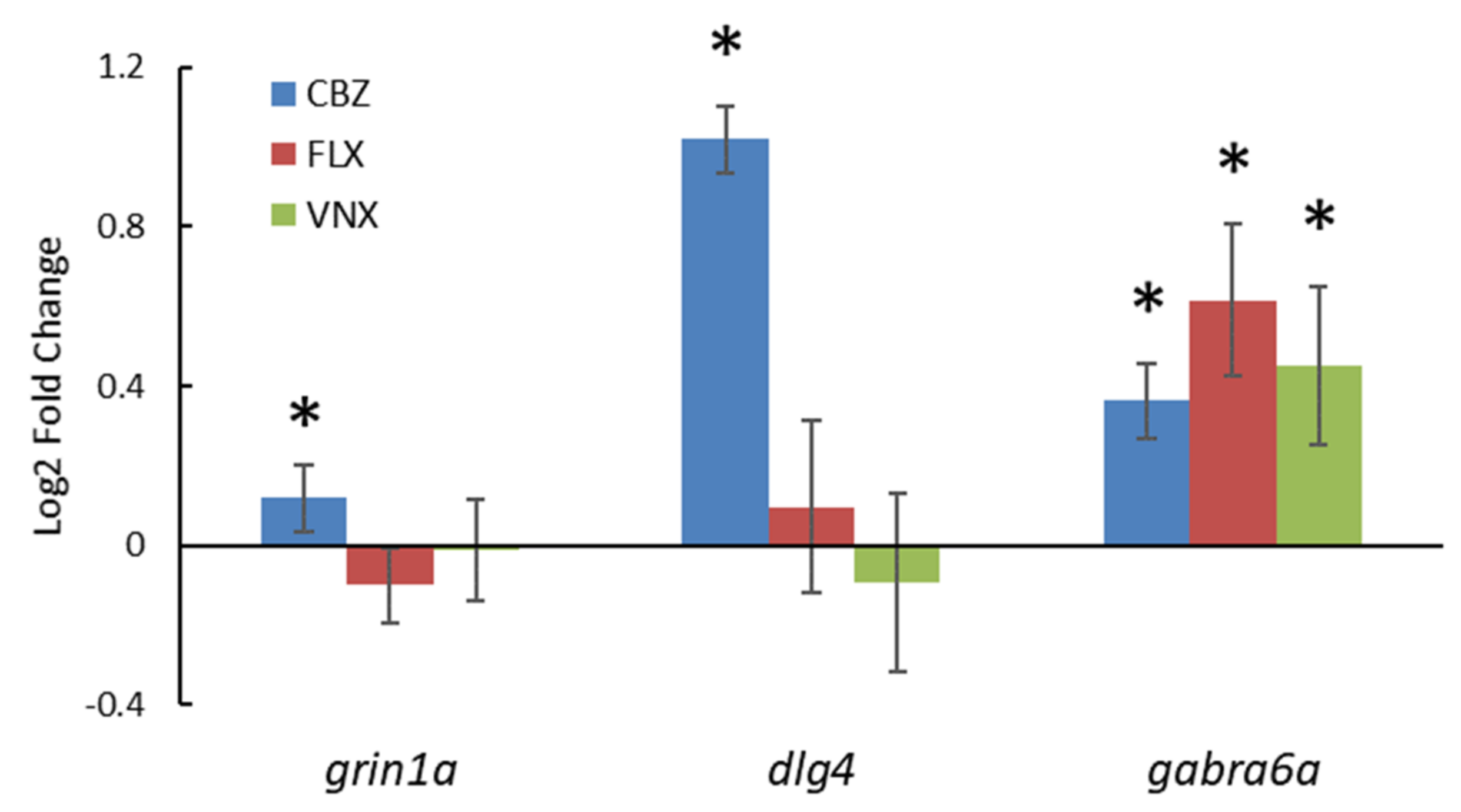
3.2.3. Gene Expression
3.2.4. RPF Estimation for Mixture Design
3.2.5. Mixture Results
4. Discussion
4.1. Prenatal Exposure to Psychoactive Compounds May Lead to Long-Term Neurobehavioural Outcomes
4.2. The Target Compounds Inhibit the Locomotor Activity of Zebrafish Embryos
4.3. MOA of the Target Compound, Potential Relation to DNT
4.4. The Contribution of Gene Expression to DNT Assessment
4.5. Risk Assessment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | CBZ | FLX | VNX | ref |
|---|---|---|---|---|
| NL | 0.075–1.5 µg/L 1 | 0.02–0.22 µg/L | [71] | |
| NL | 0.004–0.75 µg/L 2 | [72] | ||
| Germany | 1.28 µg/L | [73] | ||
| Spain | 0.12 μg/L | [74] | ||
| USA | 1.31 μg/L | [75] | ||
| PNEC 3 | 1520 µg/L | 10 µg/L | 20 µg/L |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smirnova, L.; Hogberg, H.T.; Leist, M.; Hartung, T. Developmental neurotoxicity—Challenges in the 21st century and in vitro opportunities. Altex 2014, 31, 129–156. [Google Scholar] [CrossRef]
- Hessel, E.V.S.; Staal, Y.C.M.; Piersma, A.H. Design and validation of an ontology-driven animal-free testing strategy for developmental neurotoxicity testing. Toxicol. Appl. Pharmacol. 2018, 354, 136–152. [Google Scholar] [CrossRef]
- Thapar, A.; Cooper, M.; Rutter, M. Neurodevelopmental disorders. Lancet Psychiatry 2017, 4, 339–346. [Google Scholar] [CrossRef]
- Rice, D.; Barone, S., Jr. Critical periods of vulnerability for the developing nervous system: Evidence from humans and animal models. Environ. Health Perspect 2000, 108 (Suppl. 3), 511–533. [Google Scholar] [CrossRef] [PubMed]
- Aschner, M.; Ceccatelli, S.; Daneshian, M.; Fritsche, E.; Hasiwa, N.; Hartung, T.; Hogberg, H.T.; Leist, M.; Li, A.; Mundi, W.R.; et al. Reference compounds for alternative test methods to indicate developmental neurotoxicity (DNT) potential of chemicals: Example lists and criteria for their selection and use. ALTEX 2017, 34, 49–74. [Google Scholar] [CrossRef]
- Kuegler, P. Markers of murine embryonic and neural stem cells, neurons and astrocytes: Reference points for developmental neurotoxicity testing. Altex 2010, 16–41. [Google Scholar] [CrossRef]
- Sheets, L.P.; Slikker, W., Jr. Animal/Human Concordance. In Handbook of Developmental Neurotoxicology, 2nd ed.; Slikker, W., Jr., Paule, M.G., Wang, C., Eds.; Academic Press: London, UK, 2018; pp. 527–538. [Google Scholar]
- Bal-Price, A.; Pistollato, F.; Sachana, M.; Bopp, S.K.; Munn, S.; Worth, A. Strategies to improve the regulatory assessment of developmental neurotoxicity (DNT) using in vitro methods. Toxicol. Appl. Pharmacol. 2018, 354, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Vinken, M. The adverse outcome pathway concept: A pragmatic tool in toxicology. Toxicology 2013, 312, 158–165. [Google Scholar] [CrossRef]
- Bal-Price, A.; Lein, P.J.; Keil, K.P.; Sethi, S.; Shafer, T.; Barenys, M.; Fritsche, E.; Sachana, M.; Meek, M.E.B. Developing and applying the adverse outcome pathway concept for understanding and predicting neurotoxicity. Neurotoxicology 2017, 59, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Tonk, E.C.; Pennings, J.L.; Piersma, A.H. An adverse outcome pathway framework for neural tube and axial defects mediated by modulation of retinoic acid homeostasis. Reprod. Toxicol. 2015, 55, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Echevarria, D.J.; Stewart, A.M. Gaining translational momentum: More zebrafish models for neuroscience research. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 55, 1–6. [Google Scholar] [CrossRef]
- Kaushik, G.; Xia, Y.; Pfau, J.C.; Thomas, M.A. Dysregulation of autism-associated synaptic proteins by psychoactive pharmaceuticals at environmental concentrations. Neurosci. Lett. 2017, 661, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, G.; Xia, Y.; Yang, L.; Thomas, M.A. Psychoactive pharmaceuticals at environmental concentrations induce in vitro gene expression associated with neurological disorders. BMC Genom. 2016, 17, 435. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.A.; Joshi, P.P.; Klaper, R.D. Gene-class analysis of expression patterns induced by psychoactive pharmaceutical exposure in fathead minnow (Pimephales promelas) indicates induction of neuronal systems. Comp. Biochem. Physiol. C Toxicol. Pharm. 2012, 155, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Cunha, V.; Rodrigues, P.; Santos, M.M.; Moradas-Ferreira, P.; Ferreira, M. Fluoxetine modulates the transcription of genes involved in serotonin, dopamine and adrenergic signalling in zebrafish embryos. Chemosphere 2018, 191, 954–961. [Google Scholar] [CrossRef]
- Ewing, G.; Tatarchuk, Y.; Appleby, D.; Schwartz, N.; Kim, D. Placental transfer of antidepressant medications: Implications for postnatal adaptation syndrome. Clin. Pharm. 2015, 54, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Rampono, J.; Simmer, K.; Ilett, K.F.; Hackett, L.P.; Doherty, D.A.; Elliot, R.; Kok, C.H.; Coenen, A.; Forman, T. Placental transfer of SSRI and SNRI antidepressants and effects on the neonate. Pharmacopsychiatry 2009, 42, 95–100. [Google Scholar] [CrossRef]
- Samren, E.B.; van Duijn, C.M.; Koch, S.; Hiilesmaa, V.K.; Klepel, H.; Bardy, A.H.; Mannagetta, G.B.; Deichl, A.W.; Gaily, E.; Granstrom, M.L.; et al. Maternal use of antiepileptic drugs and the risk of major congenital malformations: A joint European prospective study of human teratogenesis associated with maternal epilepsy. Epilepsia 1997, 38, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Alwan, S.; Friedman, J.M.; Chambers, C. Safety of Selective Serotonin Reuptake Inhibitors in Pregnancy: A Review of Current Evidence. CNS Drugs 2016, 30, 499–515. [Google Scholar] [CrossRef]
- Huybrechts, K.F.; Palmsten, K.; Avorn, J.; Cohen, L.S.; Holmes, L.B.; Franklin, J.M.; Mogun, H.; Levin, R.; Kowal, M.; Setoguchi, S.; et al. Antidepressant use in pregnancy and the risk of cardiac defects. N. Engl. J. Med. 2014, 370, 2397–2407. [Google Scholar] [CrossRef]
- Malm, H.; Artama, M.; Gissler, M.; Ritvanen, A. Selective serotonin reuptake inhibitors and risk for major congenital anomalies. Obs. Gynecol. 2011, 118, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Veroniki, A.A.; Cogo, E.; Rios, P.; Straus, S.E.; Finkelstein, Y.; Kealey, R.; Reynen, E.; Soobiah, C.; Thavorn, K.; Hutton, B.; et al. Comparative safety of anti-epileptic drugs during pregnancy: A systematic review and network meta-analysis of congenital malformations and prenatal outcomes. BMC Med. 2017, 15, 95. [Google Scholar] [CrossRef]
- Jentink, J.; Dolk, H.; Loane, M.A.; Morris, J.K.; Wellesley, D.; Garne, E.; de Jong-van den Berg, L.; Group, E.A.S.W. Intrauterine exposure to carbamazepine and specific congenital malformations: Systematic review and case-control study. BMJ 2010, 341, c6581. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, K.; Mitsuhashi, T.; Takahashi, T. Adverse effects of prenatal and early postnatal exposure to antiepileptic drugs: Validation from clinical and basic researches. Brain Dev. 2017, 39, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Veroniki, A.A.; Rios, P.; Cogo, E.; Straus, S.E.; Finkelstein, Y.; Kealey, R.; Reynen, E.; Soobiah, C.; Thavorn, K.; Hutton, B.; et al. Comparative safety of antiepileptic drugs for neurological development in children exposed during pregnancy and breast feeding: A systematic review and network meta-analysis. BMJ Open 2017, 7, e017248. [Google Scholar] [CrossRef] [PubMed]
- Beker van Woudenberg, A.; Snel, C.; Rijkmans, E.; de Groot, D.; Bouma, M.; Hermsen, S.; Piersma, A.; Menke, A.; Wolterbeek, A. Zebrafish embryotoxicity test for developmental (neuro)toxicity: Demo case of an integrated screening approach system using anti-epileptic drugs. Reprod. Toxicol. 2014, 49, 101–116. [Google Scholar] [CrossRef]
- Lee, S.H.; Kang, J.W.; Lin, T.; Lee, J.E.; Jin, D.I. Teratogenic potential of antiepileptic drugs in the zebrafish model. Biomed. Res. Int. 2013, 2013, 726478. [Google Scholar] [CrossRef]
- Weichert, F.G.; Floeter, C.; Meza Artmann, A.S.; Kammann, U. Assessing the ecotoxicity of potentially neurotoxic substances—Evaluation of a behavioural parameter in the embryogenesis of Danio rerio. Chemosphere 2017, 186, 43–50. [Google Scholar] [CrossRef]
- Wong, R.Y.; Oxendine, S.E.; Godwin, J. Behavioral and neurogenomic transcriptome changes in wild-derived zebrafish with fluoxetine treatment. BMC Genom. 2013, 14, 348. [Google Scholar] [CrossRef]
- Airhart, M.J.; Lee, D.H.; Wilson, T.D.; Miller, B.E.; Miller, M.N.; Skalko, R.G. Movement disorders and neurochemical changes in zebrafish larvae after bath exposure to fluoxetine (PROZAC). Neurotoxicol. Teratol. 2007, 29, 652–664. [Google Scholar] [CrossRef]
- Thompson, W.A.; Arnold, V.I.; Vijayan, M.M. Venlafaxine in Embryos Stimulates Neurogenesis and Disrupts Larval Behavior in Zebrafish. Environ. Sci. Technol. 2017, 51, 12889–12897. [Google Scholar] [CrossRef]
- Thompson, W.A.; Vijayan, M.M. Zygotic Venlafaxine Exposure Impacts Behavioral Programming by Disrupting Brain Serotonin in Zebrafish. Environ. Sci. Technol. 2020, 54, 14578–14588. [Google Scholar] [CrossRef]
- Thompson, W.A.; Vijayan, M.M. Zygotic exposure to venlafaxine disrupts cortisol stress axis activity in multiple generations of zebrafish. Environ. Pollut. 2021, 274, 116535. [Google Scholar] [CrossRef]
- Al Shuraiqi, A.; Al-Habsi, A.; Barry, M.J. Time-, dose- and transgenerational effects of fluoxetine on the behavioural responses of zebrafish to a conspecific alarm substance. Environ. Pollut. 2021, 270, 116164. [Google Scholar] [CrossRef] [PubMed]
- Vera-Chang, M.N.; St-Jacques, A.D.; Gagne, R.; Martyniuk, C.J.; Yauk, C.L.; Moon, T.W.; Trudeau, V.L. Transgenerational hypocortisolism and behavioral disruption are induced by the antidepressant fluoxetine in male zebrafish Danio rerio. Proc. Natl. Acad. Sci. USA 2018, 115, E12435–E12442. [Google Scholar] [CrossRef] [PubMed]
- De Jongh, C.M.; Kooij, P.J.; de Voogt, P.; ter Laak, T.L. Screening and human health risk assessment of pharmaceuticals and their transformation products in Dutch surface waters and drinking water. Sci. Total Environ. 2012, 427–428, 70–77. [Google Scholar] [CrossRef]
- Moermond, C.T.; Smit, C.E.; van Leerdam, R.C.; Van der Aa, N.G.; Montforts, M.H. Geneesmiddelen en waterkwaliteit. In RIVM Briefrapport 2016-0111; RIVM: Bilthoven, The Netherlands, 2016; Available online: https://www.rivm.nl/bibliotheek/rapporten/2016-0111.pdf (accessed on 20 May 2021).
- Thomas, M.A.; Klaper, R.D. Psychoactive pharmaceuticals induce fish gene expression profiles associated with human idiopathic autism. PLoS ONE 2012, 7, e32917. [Google Scholar] [CrossRef] [PubMed]
- Hermsen, S.A.; van den Brandhof, E.J.; van der Ven, L.T.; Piersma, A.H. Relative embryotoxicity of two classes of chemicals in a modified zebrafish embryotoxicity test and comparison with their in vivo potencies. Toxicol. Vitr. 2011, 25, 745–753. [Google Scholar] [CrossRef]
- Bilotta, J.; Saszik, S.; Givin, C.M.; Hardesty, H.R.; Sutherland, S.E. Effects of embryonic exposure to ethanol on zebrafish visual function. Neurotoxicol. Teratol. 2002, 24, 759–766. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef]
- Staal, Y.C.M.; Meijer, J.; van der Kris, R.J.C.; de Bruijn, A.C.; Boersma, A.Y.; Gremmer, E.R.; Zwart, E.P.; Beekhof, P.K.; Slob, W.; van der Ven, L.T.M. Head skeleton malformations in zebrafish (Danio rerio) to assess adverse effects of mixtures of compounds. Arch. Toxicol. 2018, 92, 3549–3564. [Google Scholar] [CrossRef] [PubMed]
- Slob, W. Dose-response modeling of continuous endpoints. Toxicol. Sci. 2002, 66, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Chiarotti, F.; Venerosi, A. Epidemiology of Autism Spectrum Disorders: A Review of Worldwide Prevalence Estimates Since 2014. Brain Sci. 2020, 10, 274. [Google Scholar] [CrossRef]
- Perez-Crespo, L.; Prats-Uribe, A.; Tobias, A.; Duran-Tauleria, E.; Coronado, R.; Hervas, A.; Guxens, M. Temporal and Geographical Variability of Prevalence and Incidence of Autism Spectrum Disorder Diagnoses in Children in Catalonia, Spain. Autism Res. 2019, 12, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- Stuhec, M.; Svab, V.; Locatelli, I. Prevalence and incidence of attention-deficit/hyperactivity disorder in Slovenian children and adolescents: A database study from a national perspective. Croat. Med. J. 2015, 56, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Gould, E. Childhood lead poisoning: Conservative estimates of the social and economic benefits of lead hazard control. Environ. Health Perspect 2009, 117, 1162–1167. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Developmental neurotoxicity of industrial chemicals. Lancet 2006, 368, 2167–2178. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef]
- Sehonova, P.; Hodkovicova, N.; Urbanova, M.; Orn, S.; Blahova, J.; Svobodova, Z.; Faldyna, M.; Chloupek, P.; Briedikova, K.; Carlsson, G. Effects of antidepressants with different modes of action on early life stages of fish and amphibians. Environ. Pollut. 2019, 254, 112999. [Google Scholar] [CrossRef]
- Calcagno, E.; Durando, P.; Valdes, M.E.; Franchioni, L.; Bistoni Mde, L. Effects of carbamazepine on cortisol levels and behavioral responses to stress in the fish Jenynsia multidentata. Physiol. Behav. 2016, 158, 68–75. [Google Scholar] [CrossRef]
- Nassef, M.; Matsumoto, S.; Seki, M.; Khalil, F.; Kang, I.J.; Shimasaki, Y.; Oshima, Y.; Honjo, T. Acute effects of triclosan, diclofenac and carbamazepine on feeding performance of Japanese medaka fish (Oryzias latipes). Chemosphere 2010, 80, 1095–1100. [Google Scholar] [CrossRef]
- De Farias, N.O.; Oliveira, R.; Sousa-Moura, D.; de Oliveira, R.C.S.; Rodrigues, M.A.C.; Andrade, T.S.; Domingues, I.; Camargo, N.S.; Muehlmann, L.A.; Grisolia, C.K. Exposure to low concentration of fluoxetine affects development, behaviour and acetylcholinesterase activity of zebrafish embryos. Comp. Biochem. Physiol. C Toxicol. Pharm. 2019, 215, 1–8. [Google Scholar] [CrossRef]
- Huang, I.J.; Sirotkin, H.I.; McElroy, A.E. Varying the exposure period and duration of neuroactive pharmaceuticals and their metabolites modulates effects on the visual motor response in zebrafish (Danio rerio) larvae. Neurotoxicol. Teratol. 2019, 72, 39–48. [Google Scholar] [CrossRef]
- Bertilsson, L.; Tomson, T. Clinical pharmacokinetics and pharmacological effects of carbamazepine and carbamazepine-10,11-epoxide. An update. Clin. Pharm. 1986, 11, 177–198. [Google Scholar] [CrossRef]
- Tufi, S.; Leonards, P.; Lamoree, M.; de Boer, J.; Legler, J.; Legradi, J. Changes in Neurotransmitter Profiles during Early Zebrafish (Danio rerio) Development and after Pesticide Exposure. Environ. Sci. Technol. 2016, 50, 3222–3230. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Cheng, J.; Yi, J.; Rotchell, J.M.; Zhu, X.; Zhou, J. Environmental concentration of carbamazepine accelerates fish embryonic development and disturbs larvae behavior. Ecotoxicology 2016, 25, 1426–1437. [Google Scholar] [CrossRef]
- Monesson-Olson, B.; McClain, J.J.; Case, A.E.; Dorman, H.E.; Turkewitz, D.R.; Steiner, A.B.; Downes, G.B. Expression of the eight GABAA receptor alpha subunits in the developing zebrafish central nervous system. PLoS ONE 2018, 13, e0196083. [Google Scholar] [CrossRef] [PubMed]
- Prieto, M.G.; Carreno Gutiérrez, H.; Arévalo Arévalo, R.; Chiaramoni, N.S.; Alonso, S.D.V. Effect of Risperidone and Fluoxetine on the Movement and Neurochemical Changes of Zebrafish. Open J. Med. Chem. 2012, 2, 129–138. [Google Scholar] [CrossRef]
- Pinna, G.; Costa, E.; Guidotti, A. Fluoxetine and norfluoxetine stereospecifically and selectively increase brain neurosteroid content at doses that are inactive on 5-HT reuptake. Psychopharmacology 2006, 186, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Uzunova, V.; Sheline, Y.; Davis, J.M.; Rasmusson, A.; Uzunov, D.P.; Costa, E.; Guidotti, A. Increase in the cerebrospinal fluid content of neurosteroids in patients with unipolar major depression who are receiving fluoxetine or fluvoxamine. Proc. Natl. Acad. Sci. USA 1998, 95, 3239–3244. [Google Scholar] [CrossRef] [PubMed]
- Bisesi, J.H., Jr.; Bridges, W.; Klaine, S.J. Effects of the antidepressant venlafaxine on fish brain serotonin and predation behavior. Aquat. Toxicol. 2014, 148, 130–138. [Google Scholar] [CrossRef]
- Melnyk-Lamont, N.; Best, C.; Gesto, M.; Vijayan, M.M. The antidepressant venlafaxine disrupts brain monoamine levels and neuroendocrine responses to stress in rainbow trout. Environ. Sci. Technol. 2014, 48, 13434–13442. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.A.; Vijayan, M.M. Environmental levels of venlafaxine impact larval behavioural performance in fathead minnows. Chemosphere 2020, 259, 127437. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shin, Y.; Shi, M.; Kilpatrick, D.L. Temporal control of a dendritogenesis-linked gene via REST-dependent regulation of nuclear factor I occupancy. Mol. Biol. Cell 2011, 22, 868–879. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Freeman, J.L. Zebrafish Get Connected: Investigating Neurotransmission Targets and Alterations in Chemical Toxicity. Toxics 2016, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.P.; Trimmer, J.S.; Gilthorpe, J.D.; Smith, S.J. Characterization of zebrafish PSD-95 gene family members. J. Neurobiol. 2005, 63, 91–105. [Google Scholar] [CrossRef]
- Niell, C.M.; Meyer, M.P.; Smith, S.J. In vivo imaging of synapse formation on a growing dendritic arbor. Nat. Neurosci. 2004, 7, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Moermond, C.T.A.; Montforts, M.H.M.M.; Roex, E.W.M.; Venhuis, B.J. Medicijnresten en waterkwaliteit: Een update. In RIVM-briefrapport 2020-0088; Rijksinstituut voor Volksgezondheid en Milieu RIVM: Bilthoven, The Netherlands, 2020. [Google Scholar] [CrossRef]
- RIWA-Rijn, 2011. Jaarrapport 2010—De Rijn. RIWA-Rijn, Nieuwegein. Jaarrapport 2010 De Rijn. Available online: https://www.riwa-rijn.org/wp-content/uploads/2015/05/RIWA_Jaarrapport_2010_internet.pdf (accessed on 20 May 2021).
- Ternes, T.A. Occurrence of drugs in German sewage treatment plants and rivers. Water Res. 1998, 32, 3245–3260. [Google Scholar] [CrossRef]
- Gros, M.; Petrovic, M.; Ginebreda, A.; Barcelo, D. Removal of pharmaceuticals during wastewater treatment and environmental risk assessment using hazard indexes. Environ. Int. 2010, 36, 15–26. [Google Scholar] [CrossRef]
- Schultz, M.M.; Furlong, E.T. Trace analysis of antidepressant pharmaceuticals and their select degradates in aquatic matrixes by LC/ESI/MS/MS. Anal. Chem. 2008, 80, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Kawai, H.; Iwadate, R.; Ishibashi, T.; Kudo, N.; Kawashima, Y.; Mitsumoto, A. Antidepressants with different mechanisms of action show different chronopharmacological profiles in the tail suspension test in mice. Chronobiol. Int. 2019, 36, 1194–1207. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, P.; Kus, K.; Skurzynska, M.; Nowakowska, E. The influence of aripiprazole and venlafaxine on the antidepressant-like effect observed in prenatally stressed rats (animal model of depression). Hum. Exp. Toxicol. 2018, 37, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Moermond, C.T.; Kase, R.; Korkaric, M.; Agerstrand, M. CRED: Criteria for reporting and evaluating ecotoxicity data. Environ. Toxicol. Chem. 2016, 35, 1297–1309. [Google Scholar] [CrossRef]
- EFSA-PPR-Panel. Scientific Opinion on the Relevance of Dissimilar Mode of Action and Its Appropriate Application for Cumulative Risk Assessment of Pesticides Residues in Food; EFSA-PPR-Panel: Parma, Italy, 2013; pp. 3472–3512. [Google Scholar]
- Parolini, M.; Ghilardi, A.; De Felice, B.; Del Giacco, L. Environmental concentration of fluoxetine disturbs larvae behavior and increases the defense response at molecular level in zebrafish (Danio rerio). Environ. Sci. Pollut. Res. Int. 2019, 26, 34943–34952. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, C.D.; Chu, S.; Judt, C.; Li, H.; Oakes, K.D.; Servos, M.R.; Andrews, D.M. Antidepressants and their metabolites in municipal wastewater, and downstream exposure in an urban watershed. Environ. Toxicol. Chem. 2010, 29, 79–89. [Google Scholar] [CrossRef]
- Lajeunesse, A.; Blais, M.; Barbeau, B.; Sauve, S.; Gagnon, C. Ozone oxidation of antidepressants in wastewater -Treatment evaluation and characterization of new by-products by LC-QToFMS. Chem. Cent. J. 2013, 7, 15. [Google Scholar] [CrossRef]
- Gurke, R.; Rossmann, J.; Schubert, S.; Sandmann, T.; Rossler, M.; Oertel, R.; Fauler, J. Development of a SPE-HPLC-MS/MS method for the determination of most prescribed pharmaceuticals and related metabolites in urban sewage samples. J. Chromatogr B Anal. Technol. Biomed. Life Sci. 2015, 990, 23–30. [Google Scholar] [CrossRef]
- Alygizakis, N.A.; Gago-Ferrero, P.; Borova, V.L.; Pavlidou, A.; Hatzianestis, I.; Thomaidis, N.S. Occurrence and spatial distribution of 158 pharmaceuticals, drugs of abuse and related metabolites in offshore seawater. Sci. Total Environ. 2016, 541, 1097–1105. [Google Scholar] [CrossRef] [PubMed]





| Tested Dose Ranges (µM) | ||
|---|---|---|
| Experiments Environmental Levels | Dose–Response Experiments | |
| Carbamazepine | 0.0032 a...100 | 0.3…200 |
| Carbamazepine 10,11 -epoxide | - | 0.03…30 |
| Fluoxetine | 0.00001…1 | 0.03…10 |
| Norfluoxetine | - | 0.03…10 |
| Phenytoin | - | 0.3…300 |
| Venlafaxine | 0.00032…100 | 0.3…300 |
| Desvenlafaxine | - | 0.3…300 |
| Reference Compound | Second Compound | RPF * |
|---|---|---|
| Carbamazepine | Fluoxetine | 50.50 |
| Carbamazepine | Venlafaxine | 12.65 |
| Venlafaxine | Fluoxetine | 10.26 |
| BMC05 a (µM) | BMC05-CI b (µM) | BMC05 (mg/L) | BMC05-CI(mg/L) | |
|---|---|---|---|---|
| Carbamazepine | 51.55–82.25 | 38.8–103 | 19–12 | 9.16–24.33 |
| Carbamazepine 10,11-epoxide | - | - | - | - |
| Fluoxetine | 0.17–0.65 | 0.06–1.42 | 0.05–0.2 | 0.02–0.44 |
| Norfluoxetine | 2.21 | 0.2–6.03 | 0.65 | 0.06–1.78 |
| Phenytoin | 45.13 | 4.14–52.5 | 11.38 | 1.04–13.24 |
| Venlafaxine | 0.26–1.34 | 0.10–2.87 | 0.07–0.37 | 0.03–0.8 |
| Desvenlafaxine | 6.06 | 0.69–26.7 | 1.6 | 0.18–7.03 |
| BMC50 a (µM) | BMC50-CI b (µM) | BMC50 (mg/L) | BMC50-CI(mg/L) | |
| Carbamazepine | 116–185.1 | 103–216 | 27.4–43.73 | 19–51.03 |
| Fluoxetine | 1.41–5.40 | 1.01–6.77 | 0.44–1.67 | 0.31–2.09 |
| Venlafaxine | 9.53–48.45 | 6.19–63.9 | 2.64–13.44 | 1.72–17.73 |
| Reference Compound | Second Compound | RPF | ||||
|---|---|---|---|---|---|---|
| Single Compounds | Compounds and Mix | |||||
| Lowest | Highest | Lowest | Highest | Ratio of Overlap a | ||
| Carbamazepine | Fluoxetine | 51.7 | 63 | 47.5 | 61 | 0.85 |
| Carbamazepine | Venlafaxine | 4.26 | 5.91 | 5.11 | 9.3 | 0.86 |
| Venlafaxine | Fluoxetine | 9.67 | 13.1 | 5.25 | 12.9 | 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atzei, A.; Jense, I.; Zwart, E.P.; Legradi, J.; Venhuis, B.J.; van der Ven, L.T.M.; Heusinkveld, H.J.; Hessel, E.V.S. Developmental Neurotoxicity of Environmentally Relevant Pharmaceuticals and Mixtures Thereof in a Zebrafish Embryo Behavioural Test. Int. J. Environ. Res. Public Health 2021, 18, 6717. https://doi.org/10.3390/ijerph18136717
Atzei A, Jense I, Zwart EP, Legradi J, Venhuis BJ, van der Ven LTM, Heusinkveld HJ, Hessel EVS. Developmental Neurotoxicity of Environmentally Relevant Pharmaceuticals and Mixtures Thereof in a Zebrafish Embryo Behavioural Test. International Journal of Environmental Research and Public Health. 2021; 18(13):6717. https://doi.org/10.3390/ijerph18136717
Chicago/Turabian StyleAtzei, Alessandro, Ingrid Jense, Edwin P. Zwart, Jessica Legradi, Bastiaan J. Venhuis, Leo T.M. van der Ven, Harm J. Heusinkveld, and Ellen V.S. Hessel. 2021. "Developmental Neurotoxicity of Environmentally Relevant Pharmaceuticals and Mixtures Thereof in a Zebrafish Embryo Behavioural Test" International Journal of Environmental Research and Public Health 18, no. 13: 6717. https://doi.org/10.3390/ijerph18136717
APA StyleAtzei, A., Jense, I., Zwart, E. P., Legradi, J., Venhuis, B. J., van der Ven, L. T. M., Heusinkveld, H. J., & Hessel, E. V. S. (2021). Developmental Neurotoxicity of Environmentally Relevant Pharmaceuticals and Mixtures Thereof in a Zebrafish Embryo Behavioural Test. International Journal of Environmental Research and Public Health, 18(13), 6717. https://doi.org/10.3390/ijerph18136717