
Genetic Variants in Smoking-Related Genes in Two Smoking Cessation Programs: A Cross-Sectional Study

 ,
,  , ,
, , 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Population Study
2.2. Samples Obtention
2.3. Single Nucleotide Variation Selection
2.4. Genotyping
2.5. Statistical Analysis
3. Results
3.1. Study Population
3.2. Genotype Frequencies
3.3. Metabolizer Classification by CYP2A6 Alleles
3.4. Matched Sub-Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- [ENCODAT] Instituto Nacional de Psiquiatría Ramón de la Fuente Muñiz; Instituto Nacional de Salud Pública, Comisión Nacional Contra las Adicciones, Secretaría de Salud. Encuesta Nacional de Consumo de Drogas, Alcohol y Tabaco 2016–2017: Reporte de Tabaco; Reynales-Shigematsu, L.M., Zavala-Arciniega, L., Paz-Ballesteros, W.C., Gutiérrez-Torres, D.S., García-Buendía, J.C., Rodriguez-Andrade, M.A., Gutiérrez-Reyes, J., Franco-Núñez, A., Romero-Martínez, M., y Mendoza-Alvarado, L., Eds.; INPRFM: Ciudad de México, México, 2017.
- Organización Panamericana de la Salud; Instituto Nacional de Salud Pública. Encuesta Global de Tabaquismo en Adultos; INSP/OPS: Cuernavaca, México, 2017.
- Ducci, F.; Goldman, D. The Genetic Basis of Addictive Disorders. Psychiatry Clin. N. Am. 2012, 35, 495–519. [Google Scholar] [CrossRef] [PubMed]
- Tyndale, R.F.; Zhu, A.Z.; George, T.P.; Cinciripini, P.; Hawk, L.W., Jr.; Schnoll, R.A.; Swan, G.E.; Benowitz, N.L.; Heitjan, D.F.; Lerman, C.; et al. Lack of associations of CHRNA5-A3-B4 genetic variants with smoking cessation treatment outcomes in Caucasian smokers despite associations with baseline smoking. PLoS ONE 2015, 10, e0128109. [Google Scholar] [CrossRef] [PubMed]
- Benowitz, N.L.; Hukkanen, J.; Jacob, P., 3rd. Nicotine chemistry metabolism kinetics and biomarkers. In Nicotine Psychopharmacology; Handbook of Experimental Pharmacology Series; Springer: Berlin/Heidelberg, Germany, 2009; pp. 29–60. [Google Scholar]
- Tyndale, R.F.; Mwenifumbo, J.C. Molecular genetics of nicotine metabolism. In Nicotine Psychopharmacology; Handbook of Experimental Pharmacology Series; Springer: Berlin/Heidelberg, Germany, 2009; Volume 192, pp. 235–259. [Google Scholar]
- Al Koudsi, N.; Hoffmann, E.B.; Assadzadeh, A.; Tyndale, R.F. Hepatic CYP2A6 levels and nicotine metabolism: Impact of genetic, physiological, environmental, and epigenetic factors. Eur. J. Clin. Pharmacol. 2010, 66, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Harari, O.; Martinez, M.; Zhang, X.; McDonald, S.A.; Murphy, S.E.; Goate, A. A compensatory effect upon splicing results in normal function of the CYP2A6*14 allele. Pharmacogenet. Genom. 2013, 23, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Tanner, J.A.; Tyndale, R.F. Variation in CYP2A6 activity and personalized medicine. J. Pers. Med. 2017, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-S.; Saccone, N.L.; Culverhouse, R.C.; Bracci, P.M.; Chen, C.H.; Dueker, N.; Han, Y.; Huang, H.; Jin, G.; Kohno, T.; et al. Smoking and Genetic Risk Variation Across Populations of European, Asian, and African American Ancestry—A Meta-Analysis of Chromosome 15q25. Genet. Epidemiol. 2012, 36, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Saccone, N.L.; Culverhouse, R.C.; Schwantes-An, T.-H.; Cannon, D.S.; Chen, X.; Cichon, S.; Giegling, I.; Han, S.; Han, Y.; Keskitalo-Vuokko, K.; et al. Multiple independent loci at chromosome 15q25.1 affect smoking quantity: A meta-analysis and comparison with lung cancer and COPD. PLoS Genet. 2010, 6, e1001053. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C.; Cruchaga, C.; Saccone, N.L.; Bertelsen, S.; Liu, P.; Budde, J.; Duan, W.; Fox, L.; Grucza, R.A.; Kern, J.; et al. Risk for nicotine dependence and lung cancer is conferred by mRNA expression levels and amino acid change in CHRNA5. Hum. Mol. Genet. 2009, 18, 3125–3135. [Google Scholar] [CrossRef] [PubMed]
- Bierut, L.J.; Tyndale, R.F. Preparing the Way: Exploiting Genomic Medicine to Stop Smoking. Trends Mol. Med. 2018, 24, 187–196. [Google Scholar] [CrossRef]
- Gaedigk, A.; Ingelman-Sundberg, M.; Miller, N.A.; Leeder, J.S.; Whirl-Carrillo, M.; Klein, T.E. The Pharmacogene Variation (PharmVar) Consortium: Incorporation of the Human Cytochrome P450 (CYP) Allele Nomenclature Database. Clin. Pharmacol. Ther. 2018, 103, 399–401. [Google Scholar] [CrossRef]
- Pérez-Rubio, G.; Perez-Rodriguez, M.E.; Fernández-López, J.C.; Ramirez-Venegas, A.; Garcia-Colunga, J.; Avila-Moreno, F.; Camarena, A.; Sansores, R.H.; Falfan-Valencia, R. SNPs in NRXN1 and CHRNA5 are associated to smoking and regulation of GABAergic and glutamatergic pathways. Pharmacogenomics 2016, 17, 1145–1158. [Google Scholar] [CrossRef]
- Pérez-Rubio, G.; López-Flores, L.A.; Ramírez-Venegas, A.; Noé-Díaz, V.; García-Gómez, L.; Ambrocio-Ortiz, E.; Sánchez-Romero, C.; Hernández-Zenteno, R.D.J.; Sansores, R.H.; Falfán-Valencia, R. Genetic polymorphisms in CYP2A6 are associated with a risk of cigarette smoking and predispose to smoking at younger ages. Gene 2017, 628, 205–210. [Google Scholar] [CrossRef]
- Public Health Information Department of the General Public Health Administration of the Health Consulate. EPIDAT, Program for Epidemiological Analysis of Tabulated Data, Version 3.1.; WHO: Geneva, Switzerland, 2006.
- Cupertino, A.P.; Cartujano-Barrera, F.; Ramírez, M.; Rodríguez-Bolaños, R.; Thrasher, J.F.; Pérez-Rubio, G.; Falfán-Valencia, R.; Ellerbeck, E.F.; Reynales-Shigematsu, L.M. A mobile smoking cessation intervention for Mexico (vive sin tabaco… ¡decídete!): Single-arm pilot study. J. Med. Internet Res. 2019, 7, e12482. [Google Scholar] [CrossRef]
- Dempsey, D.; Tutka, P.; Iii, P.J.P.; Allen, F.; Schoedel, K.; Tyndale, R.; Benowitz, N.L. Nicotine metabolite ratio as an index of cytochrome P450 2A6 metabolic activity. Clin. Pharmacol. Ther. 2004, 76, 64–72. [Google Scholar] [CrossRef]
- Pérez-Rubio, G.; Ramírez-Venegas, A.; Díaz, V.N.; Gómez, L.G.; Fabián, K.E.; Carmona, S.G.; López-Flores, L.A.; Ambrocio-Ortiz, E.; Romero, R.C.; Alcantar-Ayala, N.; et al. Polymorphisms in HTR2A and DRD4 Predispose to Smoking and Smoking Quantity. PLoS ONE 2017, 12, e0170019. [Google Scholar]
- Moreno-Coutiño, A.; Villalobos-Gallegos, L. Psychometric Properties of the Fagerström Test for Nicotine Dependence in a Sample of Mexican Smokers. J. Addict. Nurs. 2017, 28, 27–33. [Google Scholar] [CrossRef]
- Pérez-Rubio, G.; Silva-Zolezzi, I.; Fernández-López, J.C.; Camarena, Á.; Velázquez-Uncal, M.; Morales-Mandujano, F.; Hernández-Zenteno, R.D.J.; Flores-Trujillo, F.; Sánchez-Romero, C.; Velázquez-Montero, A.; et al. Genetic Variants in IL6R and ADAM19 are Associated with COPD Severity in a Mexican Mestizo Population. COPD J. Chronic Obstr. Pulm. Dis. 2016, 13, 610–615. [Google Scholar] [CrossRef]
- Vujkovic, M.; Bellamy, S.L.; Zuppa, A.F.; Gastonguay, M.R.; Moorthy, G.S.; Ratshaa, B.; Han, X.; Steenhoff, A.P.; Mosepele, M.; Strom, B.L.; et al. Polymorphisms in cytochrome P450 are associated with extensive efavirenz pharmacokinetics and CNS toxicities in an HIV cohort in Botswana. Pharmacogenom. J. 2018, 18, 678–688. [Google Scholar] [CrossRef]
- Glatard, A.; Dobrinas, M.; Gholamrezaee, M.; Lubomirov, R.; Cornuz, J.; Csajka, C.; Eap, C.B. Association of nicotine metabolism and sex with relapse following varenicline and nicotine replacement therapy. Exp. Clin. Psychopharmacol. 2017, 25, 353–362. [Google Scholar] [CrossRef]
- Svyryd, Y.; Ramírez-Venegas, A.; Hernandez, B.S.; Aguayo-Gómez, A.; Luna-Muñoz, L.; Arteaga-Vazquez, J.; Regalado-Piñeda, J.; Mutchinick, O.M. Genetic Risk Determinants for Cigarette Smoking Dependence in Mexican Mestizo Families. Nicotine Tob. Res. 2016, 18, 620–625. [Google Scholar] [CrossRef]
- Li, M.D.; Xu, Q.; Lou, X.-Y.; Payne, T.J.; Niu, T.; Ma, J.Z. Association and interaction analysis of variants in CHRNA5/CHRNA3/CHRNB4 gene cluster with nicotine dependence in African and European Americans. Am. J. Med. Genet. B. Neuropsychiatr. Genet. 2010, 153B, 745–756. [Google Scholar]
- Evans, D.E.; MacQueen, D.A.; Jentink, K.G.; Park, J.Y.; Lin, H.-Y.; Drobes, D.J. CHRNA5 variants moderate the effect of nicotine deprivation on a neural index of cognitive control. Genes. Brain. Behav. 2014, 13, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.M.; Xiao, Y.; Rice, T.; Bracci, P.M.; Wrensch, M.R.; Sison, J.D.; Chang, J.S.; Smirnov, I.V.; Patoka, J.; Seldin, M.F.; et al. Fine mapping of chromosome 15q25.1 lung cancer susceptibility in African-Americans. Hum. Mol. Genet. 2010, 19, 3652–3661. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.J.; Fanous, A.H.; Chen, J.; Kendler, K.S.; Chen, X. Variants in the 15q25 gene cluster are associated with risk for schizophrenia and bipolar disorder. Psychiatr. Genet. 2013, 23, 20–28. [Google Scholar] [CrossRef] [PubMed]


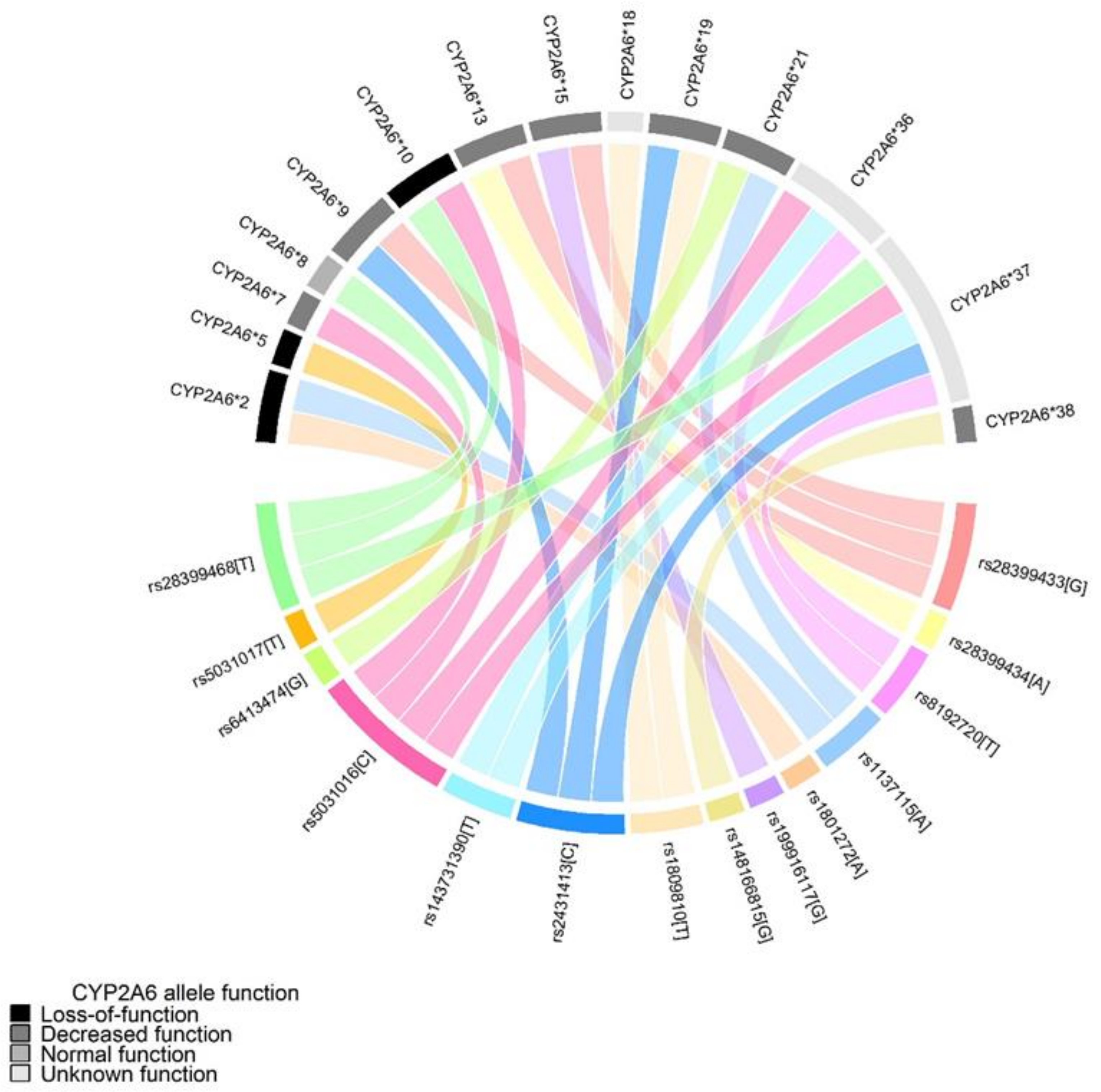{kind=link}
| SNP | Chr Position | Allele Change | MAF (GnomAD) | Consequence/Gene Location | Residue | |
|---|---|---|---|---|---|---|
| Change | Position | |||||
| rs28399433 | chr19:40850474 | A > C | C = 0.10323 | 2KB Upstream Variant | NA | NA |
| rs28399434 | chr19:40850414 | C > T | T = 0.00003 | Missense Variant | G [Gly] ⇒ R [Arg] | 5 |
| rs8192720 | chr19:40850405 | G > A | A = 0.02214 | Synonymous Variant | L [Leu] ⇒ L [Leu] | 8 |
| rs1137115 | chr19:40850376 | T > C | T = 0.24442 | Synonymous Variant | V [Val] ⇒ V [Val] | 17 |
| rs1801272 | chr19:40848628 | A > T | T = 0.01974 | Missense Variant | L [Leu] ⇒ H [His] | 160 |
| rs199916117 | chr19:40848293 | T > C | C = 0.00058 | Missense Variant | K [Lys] ⇒ E [Glu] | 194 |
| rs148166815 | chr19:40845404 | A > G | G = 0.00008 | Missense Variant | Y [Tyr] ⇒ H [His] | 351 |
| rs1809810 | chr19:40844759 | T > A | A = 0.01259 | Missense Variant | Y [Tyr] ⇒ F [Phe] | 392 |
| rs2431413 | chr19:40844073 | A > G | G = 0.12103 | Intron Variant | NA | NA |
| rs143731390 | chr19:40843969 | T > A | A = 0.1020 | Missense Variant | N [Asn] ⇒ Y [Tyr] | 438 |
| rs5031016 | chr19:40843869 | A > G | G = 0.00775 | Missense Variant | I [Ile] ⇒ T [Thr] | 471 |
| rs6413474 | chr19:40843854 | T > C | C = 0.01235 | Missense Variant | K [Lys] ⇒ R [Arg] | 476 |
| rs5031017 | chr19:40843845 | C > A | A = 0.00049 | Missense Variant | G [Gly] ⇒ V [Val] | 479 |
| rs28399468 | chr19:40843827 | C > A | A = 0.00165 | Missense Variant | R [Arg] ⇒ P [Pro] | 485 |
| SNP | Gene | Chr Position | Allele Change | MAF (GnomAD) | Consequence/Gene Location |
|---|---|---|---|---|---|
| rs10189159 | NRXN1 | chr2:50487433 | T > C | C = 0.3286 | Intron Variant |
| rs985919 | chr2:50459875 | C > A | A = 0.4957 | Intron Variant | |
| rs10865246 | chr2:50443116 | C > A | C = 0.4950 | Intron Variant | |
| rs1882296 | chr2:50039707 | T > C | C = 0.3900 | Intron Variant | |
| rs1800955 | DRD4 | chr11:636784 | T > C | C = 0.406 | 2KB Upstream Variant |
| rs6311 | HTR2A | chr13:46897343 | C > T | T = 0.3970 | 2KB Upstream Variant |
| rs6313 | chr13:46895805 | G > A | A = 0.40701 | Synonymous Variant | |
| rs12914385 | CHRNA3 | chr15:78606381 | C > A | T = 0.3138 | Intron Variant |
| rs1317286 | chr15:78603787 | A > G | G = 0.2974 | Intron Variant | |
| rs6495307 | chr15:78597979 | C > T | T = 0.310 | Intron Variant | |
| rs615470 | chr15:78593646 | T > C | T = 0.3688 | Intron Variant | |
| rs16969968 | CHRNA5 | chr15:78590583 | G > A | A = 0.26553 | Missense Variant |
| rs17408276 | chr15:78589276 | T > C | C = 0.2881 | Intron Variant | |
| rs951266 | chr15:78586199 | G > A | A = 0.2522 | Intron Variant | |
| rs680244 | chr15:78578946 | T > C | T = 0.4098 | Intron Variant | |
| rs17486278 | chr15:78575140 | A > C | C = 0.3167 | Intron Variant | |
| rs4105144 | CYP2A6 near | chr19:40852719 | T > C | C = 0.4412 | Intergene |
| Variable | e-Dec (n = 38) | On-Site (n = 94) | p * |
|---|---|---|---|
| Age (years) | 35 (20–60) | 36 (20–40) | 0.700 |
| Male/Female n, (%) | 24/14 (63.2/36.8) | 57/37 (60.6/39.4) | 0.942 |
| Cigarettes per day | 9 (2–30) | 10 (1–60) | 0.064 |
| Years of smoking | 12 (2–44) | 16 (1–43) | 0.049 |
| Onset smoking age | 20.5 (12–50) | 17.5 (9–50) | 0.004 |
| Tobacco index | 6 (0.6–66.0) | 9.8 (0.1–38.7) | 0.015 |
| SNP Genotype/Allele | e-Dec | On-Site | p * | ||
|---|---|---|---|---|---|
| n = 38 | GF/AF % | n = 94 | GF/AF % | ||
| rs28399433 | |||||
| TT | 30 | 78.95 | 32 | 34.04 | <0.0001 |
| TG | 7 | 18.42 | 59 | 62.77 | |
| GG | 1 | 2.63 | 3 | 3.19 | |
| T | 67 | 88.16 | 123 | 65.43 | 0.0003 |
| G | 9 | 11.84 | 65 | 34.57 | |
| rs28399434 | |||||
| GG | 38 | 100.00 | 88 | 93.62 | NS |
| GA | 0 | 0.00 | 6 | 6.38 | |
| AA | 0 | 0.00 | 0 | 0 | |
| G | 76 | 100 | 182 | 96.81 | NS |
| A | 0 | 0 | 6 | 3.19 | |
| rs8192720 | |||||
| CC | 37 | 97.37 | 94 | 100 | NS |
| CT | 1 | 2.63 | 0 | 0 | |
| TT | 0 | 0.00 | 0 | 0 | |
| C | 75 | 98.68 | 188 | 100.00 | NS |
| T | 1 | 1.32 | 0 | 0.00 | |
| rs1137115 | |||||
| GG | 27 | 71.05 | 69 | 73.4 | NS |
| GA | 10 | 26.32 | 23 | 24.47 | |
| AA | 1 | 2.63 | 2 | 2.13 | |
| G | 64 | 84.21 | 161 | 85.64 | NS |
| A | 12 | 15.79 | 27 | 14.36 | |
| rs2431413 | |||||
| TT | 38 | 100.00 | 69 | 73.4 | NS |
| TC | 0 | 0.00 | 22 | 23.4 | |
| CC | 0 | 0.00 | 3 | 3.19 | |
| T | 76 | 100.00 | 160 | 85.11 | NS |
| C | 0 | 0.00 | 28 | 14.89 | |
| rs5031017 | |||||
| GG | 19 | 50.00 | 43 | 45.74 | NS |
| GT | 13 | 34.21 | 35 | 37.23 | |
| TT | 6 | 15.79 | 16 | 17.02 | |
| G | 51 | 67.11 | 121 | 64.36 | NS |
| T | 25 | 32.89 | 67 | 35.64 | |
| Gene/SNP Genotype/Allele | e-Dec | On-Site | p * | ||
|---|---|---|---|---|---|
| n = 38 | GF/AF % | n = 94 | GF/AF % | ||
| NRXN1 | |||||
| rs10189159 | |||||
| TT | 32 | 84.21 | 44 | 46.8 | 0.0004 |
| TC | 5 | 13.16 | 45 | 47.87 | |
| CC | 1 | 2.63 | 5 | 5.32 | |
| T | 69 | 90.79 | 133 | 70.74 | 0.0009 |
| C | 7 | 9.21 | 55 | 29.26 | |
| rs985919 | |||||
| TT | 21 | 55.26 | 29 | 30.85 | 0.0333 |
| TG | 14 | 36.84 | 54 | 57.44 | |
| GG | 3 | 7.89 | 11 | 11.7 | |
| T | 56 | 73.68 | 112 | 59.57 | 0.0437 |
| G | 20 | 26.32 | 76 | 40.43 | |
| rs10865246 | |||||
| AA | 21 | 55.26 | 38 | 40.42 | 0.0621 |
| AC | 15 | 39.47 | 42 | 44.68 | |
| CC | 2 | 5.26 | 14 | 14.89 | |
| A | 57 | 75.00 | 118 | 62.77 | 0.0783 |
| C | 19 | 25.00 | 70 | 37.23 | |
| rs1882296 | |||||
| AA | 26 | 68.42 | 45 | 47.87 | 0.0348 |
| AG | 11 | 28.95 | 43 | 45.74 | |
| GG | 1 | 2.63 | 6 | 6.38 | |
| A | 63 | 82.89 | 133 | 70.74 | 0.0589 |
| G | 13 | 17.11 | 55 | 29.26 | |
| DRD4 | |||||
| rs1800955 | |||||
| TT | 19 | 50 | 43 | 45.74 | 0.2936 |
| TC | 17 | 44.74 | 37 | 39.36 | |
| CC | 2 | 5.26 | 14 | 14.89 | |
| T | 55 | 72.37 | 123 | 65.43 | <0.0001 |
| C | 21 | 27.63 | 65 | 34.57 | |
| HTR2A | |||||
| rs6311 | |||||
| GG | 19 | 50 | 35 | 37.23 | 0.3103 |
| GA | 14 | 36.84 | 46 | 48.93 | |
| AA | 5 | 13.16 | 13 | 13.82 | |
| G | 52 | 68.42 | 116 | 61.70 | 0.3750 |
| A | 24 | 31.58 | 72 | 38.30 | |
| rs6313 | |||||
| CC | 22 | 57.89 | 77 | 81.91 | 0.0772 |
| CT | 14 | 36.84 | 9 | 9.57 | |
| TT | 2 | 5.26 | 8 | 8.51 | |
| C | 58 | 76.32 | 163 | 86.70 | 0.0594 |
| T | 18 | 23.68 | 25 | 13.30 | |
| CHRNA3 | |||||
| rs12914385 | |||||
| CC | 23 | 60.53 | 49 | 52.12 | 0.6203 |
| CT | 12 | 31.58 | 40 | 42.55 | |
| TT | 3 | 7.89 | 5 | 5.32 | |
| C | 58 | 76.32 | 138 | 73.40 | 0.7380 |
| T | 18 | 23.68 | 50 | 26.60 | |
| rs1317286 | |||||
| AA | 28 | 73.68 | 49 | 52.12 | 0.0311 |
| AG | 9 | 23.68 | 40 | 42.55 | |
| GG | 1 | 2.63 | 5 | 5.32 | |
| A | 63 | 82.89 | 138 | 73.40 | 0.1390 |
| G | 13 | 17.11 | 50 | 26.60 | |
| rs6495307 | |||||
| CC | 29 | 76.32 | 53 | 56.38 | 0.0903 |
| CT | 7 | 18.42 | 37 | 39.36 | |
| TT | 2 | 5.26 | 4 | 4.25 | |
| C | 65 | 85.53 | 143 | 76.06 | 0.1240 |
| T | 11 | 14.47 | 45 | 23.94 | |
| rs615470 | |||||
| TT | 31 | 81.58 | 57 | 60.63 | 0.0355 |
| TC | 6 | 15.79 | 33 | 35.1 | |
| CC | 1 | 2.63 | 4 | 4.25 | |
| T | 68 | 89.47 | 147 | 78.19 | 0.0499 |
| C | 8 | 10.53 | 41 | 21.81 | |
| CHRNA5 | |||||
| rs16969968 | |||||
| GG | 25 | 65.79 | 53 | 56.38 | 0.3700 |
| GA | 11 | 28.95 | 35 | 37.23 | |
| AA | 2 | 5.26 | 6 | 6.38 | |
| G | 61 | 80.26 | 141 | 75.00 | 0.4510 |
| A | 15 | 19.74 | 47 | 25.00 | |
| rs17408276 | |||||
| TT | 31 | 81.58 | 53 | 56.38 | 0.0161 |
| TC | 6 | 15.79 | 39 | 41.48 | |
| CC | 1 | 2.63 | 2 | 2.12 | |
| T | 62 | 81.58 | 145 | 77.13 | 0.5280 |
| C | 14 | 18.42 | 43 | 22.87 | |
| rs951266 | |||||
| CC | 36 | 94.74 | 51 | 54.25 | <0.0001 |
| CT | 2 | 5.26 | 38 | 40.42 | |
| TT | 0 | 0 | 5 | 5.31 | |
| C | 74 | 97.37 | 140 | 74.47 | <0.0001 |
| T | 2 | 2.63 | 48 | 25.53 | |
| rs680244 | |||||
| GG | 27 | 71.05 | 52 | 55.32 | 0.1006 |
| GA | 10 | 26.32 | 37 | 39.36 | |
| AA | 1 | 2.63 | 5 | 5.31 | |
| G | 64 | 84.21 | 141 | 75.00 | 0.1430 |
| A | 12 | 15.79 | 47 | 25.00 | |
| rs17486278 | |||||
| AA | 24 | 63.16 | 46 | 48.93 | 0.3213 |
| AC | 11 | 28.95 | 43 | 45.74 | |
| CC | 3 | 7.89 | 5 | 5.31 | |
| A | 59 | 77.63 | 153 | 81.38 | 0.6000 |
| C | 17 | 22.37 | 35 | 18.62 | |
| near to CYP2A6 | |||||
| rs4105144 | |||||
| TT | 26 | 68.42 | 62 | 65.95 | 0.9709 |
| TC | 10 | 26.32 | 29 | 30.85 | |
| CC | 2 | 5.26 | 3 | 3.19 | |
| T | 62 | 81.58 | 153 | 81.38 | 0.9701 |
| C | 14 | 18.42 | 35 | 18.62 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Rubio, G.; López-Flores, L.A.; Cupertino, A.P.; Cartujano-Barrera, F.; Reynales-Shigematsu, L.M.; Ramírez, M.; Ellerbeck, E.F.; Rodríguez-Bolaños, R.; Falfan-Valencia, R. Genetic Variants in Smoking-Related Genes in Two Smoking Cessation Programs: A Cross-Sectional Study. Int. J. Environ. Res. Public Health 2021, 18, 6597. https://doi.org/10.3390/ijerph18126597
Pérez-Rubio G, López-Flores LA, Cupertino AP, Cartujano-Barrera F, Reynales-Shigematsu LM, Ramírez M, Ellerbeck EF, Rodríguez-Bolaños R, Falfan-Valencia R. Genetic Variants in Smoking-Related Genes in Two Smoking Cessation Programs: A Cross-Sectional Study. International Journal of Environmental Research and Public Health. 2021; 18(12):6597. https://doi.org/10.3390/ijerph18126597
Chicago/Turabian StylePérez-Rubio, Gloria, Luis Alberto López-Flores, Ana Paula Cupertino, Francisco Cartujano-Barrera, Luz Myriam Reynales-Shigematsu, Mariana Ramírez, Edward F. Ellerbeck, Rosibel Rodríguez-Bolaños, and Ramcés Falfan-Valencia. 2021. "Genetic Variants in Smoking-Related Genes in Two Smoking Cessation Programs: A Cross-Sectional Study" International Journal of Environmental Research and Public Health 18, no. 12: 6597. https://doi.org/10.3390/ijerph18126597
APA StylePérez-Rubio, G., López-Flores, L. A., Cupertino, A. P., Cartujano-Barrera, F., Reynales-Shigematsu, L. M., Ramírez, M., Ellerbeck, E. F., Rodríguez-Bolaños, R., & Falfan-Valencia, R. (2021). Genetic Variants in Smoking-Related Genes in Two Smoking Cessation Programs: A Cross-Sectional Study. International Journal of Environmental Research and Public Health, 18(12), 6597. https://doi.org/10.3390/ijerph18126597