Abstract
Chironomidae larvae play an important role in the food chain of river ecosystems in Korea, where it is dominant. However, detailed information on the diet of Chironomidae larvae are still lacking. The purpose of this study was to identify the gut contents of 4th instar larvae of a Chironomidae inhabiting four large-scale weirs (Sejong Weir, Juksan Weir, Gangjeong-Goryeong Weir, and Dalseong Weir) using a DNA meta-barcoding approach. We found that dominant Operational Taxonomic Unit (OUT) was assigned to Paractinolaimus sp. (Nematoda), and the sub-dominant OTU was assigned to Dicrotendipes fumidus (Chironomidae). The most common OTUs among the individuals included phytoplankton, such as Tetrahymena sp., D. armatus, Pseudopediastrum sp., Tetradesmus dimorphus, Biddulphia tridens, and Desmodesmus spp. We calculated the selectivity index (E’) and provided scientific evidence that Chironomidae larvae have a significant preference (E’ > 0.5) for Desmodesmus armatus, E. minima, and T. dimorphus, while it does not show preference for other species found in its gut. Differences in physico-chemical factors, such as water quality, nutrients, Chl-a, and carbon concentrations, resulting from anthropogenic impacts (i.e., construction of large-scale weirs) as well as the particle size of prey organisms (small-sized single cell) and effects of chemicals (chemokinesis) could affect the feeding behavior of Chironomidae larvae.
1. Introduction
Chironomidae is a large group of invertebrates, with a reported diversity of 8000–20,000 species, and its members are distributed worldwide [1]. Chironomidae adults inhabit areas near the riparian zone of rivers or lakes, but the larvae are aquatic organisms that are distributed in diverse aquatic habitat patches [2]. The 1st instar larvae starts its lifecycle by settling on the water surface of aquatic ecosystems after hatching from its egg [1]. The 3rd or 4th instar in the bottom substrates has a formed cage [1] and starts to filter phytoplankton and predate on organisms, such as zooplankton and other small organisms [3,4,5]. The life cycle of a Chironomidae larvae is sensitive to anthropogenic impacts, such as changes in habitat traits and water quality, and it is also important to fish and birds as a food source [6,7]. Therefore, identifying the food of the 3rd or 4th instar larvae of a Chironomidae species is critical for understanding the role of Chironomidae in aquatic ecosystems.
Recently, 16 large-scale weirs were built along the main channels of the four largest rivers (the Han, Nakdong, Geum, and Yeongsan) in South Korea to stimulate development of water resources for recreation and other purposes [8,9,10]. The construction of the weirs, which involved the dredging of the riverbed, channelization, and removal of riparian vegetation and large woody debris, resulted in dramatic alterations in the geography of the construction sites [11]. These changes have led to physico-chemical and habitat alterations, as well as a shift from lotic to more lentic conditions in the upper part of the weirs. These anthropogenic impacts may directly affect aquatic organism (i.e., fish, planktons, and macroinvertebrates), as well as the biodiversity and food-web structure [12,13,14,15]. Therefore, understanding the linkage among aquatic organisms is important to manage and control artificial environments, such as weirs.
Gut-content analysis is a fundamental step in the determination of food-web structures [16], and microscopic identification (MI) has conventionally been used for the analysis of gut contents. However, most gut content-analysis studies based on MI have the following disadvantages: (1) ambiguous prey specimen identification because of extensive digestion, (2) the presence of unidentified partial tissues, (3) identification failure due to a lack of expert knowledge, and 4) low-level identification resolution (identification of levels only higher than the family or order level). In addition, MI is unsuitable for tiny predators, such as a Chironomidae larvae and rotifers [17,18]. Applying DNA sequence-based techniques to gut-content identification has recently increased identification resolution, particularly in marine ecosystems [19,20,21,22,23,24]. However, relatively few studies have used DNA barcoding for gut-content analysis in complex freshwater ecosystems [25,26,27] although this technique has been recognized as a promising tool for studying food-web interactions [28].
The objectives of this study were therefore to examine the pattern of prey selection (selectivity index [E’]) by the 4th instar larvae of a Chironomidae using an MiSeq™ NGS platform (Illumina, San Diego, CA, USA). Chironomidae is omnivorous and plays an important role in the food chain of river ecosystems in Korea, where it is dominant. We aimed to evaluate (1) the pattern of food sources selection by Chironomidae in the large-scale weirs and (2) the applicability and effectiveness of the DNA meta-barcoding approach for identification of food selection by Chironomidae.
2. Materials and Methods
2.1. Study Area and Field Sampling
A survey was carried out at four study sites in the Gum River (SJ: Sejong Weir), Yeongsan River (JS: Juksan Weir), and Nakdong River (GG: Gangjeong-Goryeong Weir, DS: Dalseong Weir) from June to July 2019 (Figure 1; Appendix A, Table A1). We sampled the surface water for water quality (approximately the top 50 cm). Water temperature (Temp., °C), conductivity (Cond., µS/cm), dissolved oxygen (DO, mg/l), pH, and turbidity (NTU) were measured on-site using portable equipment (Model: YSI Professional Plus, Ohio, USA). The nutrients, chlorophyll-a (Chl-a), and carbon concentrations were analyzed in the laboratory. For total phosphorus (TP), total nitrogen (TN), and Chl-a concentration measurements, water samples were first filtered through a 0.45 μm pore-size membrane (Model: Advantec MFS membrane filter, Dublin, USA) and measurements were then performed using a UV spectrophotometer. Dissolved organic carbon (DOC) and total organic carbon (TOC) concentrations were measured using a TOC analyzer (Model: vario TOC cub, Langenselbold, Germany) through an 850 °C combustion catalytic-oxidation method. To collect 4th instar larvae from Chronomidae individuals, we used a Surber net (25 cm × 20 cm), dredging (1 m × 1 m), Ekman grab, and Ponar grab. After capture, the 4th instar larvae Chronomidae individuals were preserved in 96% ethanol and stored at room temperature for laboratory DNA meta-barcoding analysis.
Figure 1.
Map of the study sites in the Gum River (SJ: Sejong Weir), Yeongsan River (JS: Juksan Weir), and Nakdong River (GG: Gangjeong-Goryeong Weir, DS: Dalseong Weir).
2.2. DNA Extraction of Gut Contents and Metagenomic Sequencing
The gut contents were removed from the guts of 4th instar larvae of a Chironomidae (n = 12, 3 individuals each study site), and the dissection process was carried out after the complete volatilization of ethanol following the steps listed in Appendix A, Table A2. Genomic DNA was extracted using a DNeasy Blood & Tissue Kit (Cat. No. 69504, Qiagen, Düsseldorf, Germany) according to the manufacturer’s protocol. gDNA extracted for sequencing was prepared according to Illumina 18S Metagenomic Sequencing Library protocols (San Diego, USA). DNA quantity, quality, and integrity were measured using PicoGreen (Thermo Fisher Scientific, Waltham, USA) and a VICTOR Nivo Multimode Microplate Reader (PerkinElmer, Ohio, USA).
We selected two regions to amplify in the gDNA extracted from 4th instar larvae of a Chironomidae gut contents: the V9 region of the 18S rDNA gene (18S V9), primarily because of its broad range among eukaryotes [29,30]. The 18S rRNA gene was amplified using primers including an adaptor sequence: Forward Primer: 5’ TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGC- CCTGCCHTTTGTACACAC 3’ / Reverse Primer: 5’ GTCTCGTGGGCTCGGAGATGTGTATAAGA- GACAGCCTTCYGCAGGTTCACCTAC 3’. First, to amplify the target region corresponding to the adapters, one cycle of 3 min at 95 °C; 25 cycles of 30 s at 95 °C, 30 s at 55 °C, and 30 s at 72 °C; and a final step of 5 min at 72 °C were carried out using the 18S V9 primers. Second, to perform indexing PCR, the first PCR product was amplified using one cycle of 3 min at 95 °C; 8 cycles of 30 s at 95 °C, 30 s at 55 °C, and 30 s at 72 °C; and a final step of 5 min at 72°C. The final products were normalized and pooled using PicoGreen (Thermo Fisher Scientific, USA), and the sizes of the libraries were verified using the LabChip GX HT DNA High Sensitivity Kit (PerkinElmer, Massachusetts, USA).
The library was sequenced using the MiSeq™ NGS platform (Illumina, San Diego, CA, USA) provided as a commercial service (Macrogen Inc., Seoul, Korea). Raw reads were trimmed using CD-HIT-OTU [31], and chimeras were identified and removed using rDnaTools. For paired-end merging, FLASH (Fast Length Adjustment of Short reads) version 1.2.11 was used [32]. Merged reads were processed using Qiime version 1.9 [33] and were clustered into operational taxonomic units (OTUs) using UCLUST [34] with a greedy algorithm employing OTUs at a 97% OTU cutoff value. Taxonomic classifications were assigned to the obtained representative sequences using BLASTn [35] and UCLUST [34].
2.3. Data Collection and Statistical Analysis
We implemented a literature survey to determine potential prey phytoplankton of Chironomidae larvae at the study sites [34]. To determine the prey selectivity of Chironomidae larvae, we calculated the selectivity index (E’) [36] using the relative abundances in water (from where Chironomidae larvae was obtained) of phytoplankton species in the gut content of Chironomidae larvae. An E’ over 0.5 indicated a preference for a prey.
3. Results
3.1. Meta-Barcoding and Taxonomic Assignment of Operational Taxonomic Unit (OTU)
In total, 1,019,526 paired-end reads were generated from the eight samples using 18SV9 primer sets on the Illumina MiSeq™ platform (Illumina, San Diego, CA, USA); of these, 98.0% passed Q30 (Phred quality score > 30) for improving the accuracy of sequences in this study. Each sample yielded 60,767–108,924 paired-end reads (mean: 89,970 reads), similar to the number of reads reported in a previous study [37], and all samples exhibited saturation of the number of OTUs by rarefaction curve analysis. Gamma-diversity was 381 OTUs, which were produced with a similarity cutoff of 97%. The resulting 381 OTUs were classified into 21 species- or genus-level taxonomic groups (those presenting < 0.1% abundance were removed). Uncultured and non-assigned reads were discarded.
After the assignment was performed, OTUs belonging to 16 orders, including 17 families, were found based on a BLASTn search of the NCBI database (Table 1). The abundances of the assigned sequences showed different patterns among the individuals (Figure 2). The dominant OTU was assigned to Paractinolaimus sp. (Nematoda), and the sub-dominant OTU was assigned to Dicrotendipes fumidus (Chironomidae). However, the most common OTUs among the individuals included phytoplankton, such as Tetrahymena sp., D. armatus, Pseudopediastrum sp., Tetradesmus dimorphus, Biddulphia tridens, and Desmodesmus sp. (Figure 2). Interestingly, we found an OTU sequence that was common among all individuals from Hemibarbus labeo, which is a benthic fish (Table 1).

Table 1.
List of the Operational Taxonomic Unit (OTU) in the gut contents of Chironomidae based on the 18SV9 region (SJ.A-B: Sejong Weir, JS.D-E: Juksan Weir, GG.A-D: Gangjeong-Goryeong Weir, and DS.B-E: Dalseong Weir).
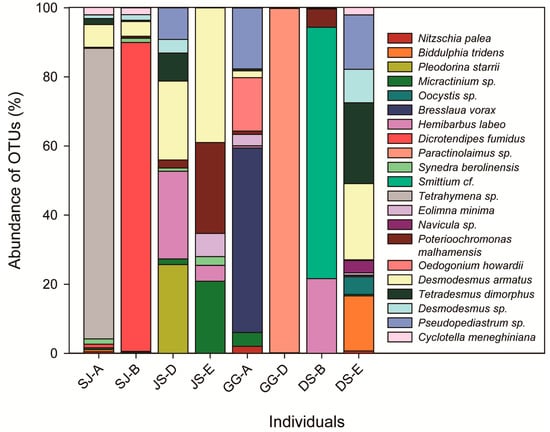
Figure 2.
Abundance of OTUs among the individuals in the gut contents of Chironomidae based on the species or genus identification level (18SV9 regions, SJ.A-B: Sejong Weir, JS.D-E: Juksan Weir, GG.A-D: Gangjeong-Goryeong Weir, and DS.B-E: Dalseong Weir).
3.2. Ecological Traits Based on Selectivity Index (E’) and Water Quality
Chironomidae larvae mainly consumed planktonic prey. Calculating the selectivity index (E’) using the relative abundance of gut-content phytoplankton species in the water from which Chironomidae larvae were obtained revealed differences in prey selectivity (Table 2) [36]. Chironomidae larvae showed a significant preference (E’ > 0.5) for D. armatus, E. minima, and T. dimorphus, while it showed negative selection for other species, even the ones that were found in its gut.
Table 2.
Relative abundances and selectivity indexes (E’) of phytoplankton species present in the gut content of Chironomidae in the water from where Chironomidae was obtained.
N. palea and E. minima appeared with higher DO, pH, and Chl-a values and Tetrahymena sp. appeared with higher NTU and TP values and lower Cond., TN, DOC, and TOC values, from the other prey sources OTUs (Table 1 and Table 3). Nutrient-related factors (Chl-a, TN, and TP) were related with N. palea and E. minima (small single cells), and carbon-related factors (DOC and TOC) were negatively related with Tetrahymena sp. (free swimming but chemokinetic cells).
Table 3.
Detailed information of water quality along sites in the Gum River (SJ: Sejong Weir), Yeongsan River (JS: Juksan Weir), and Nakdong River (GG: Gangjeong-Goryeong Weir, DS: Dalseong Weir) (Water temperature (Temp., °C), conductivity (Cond., µS/cm), dissolved oxygen (DO, mg/l), pH, turbidity (NTU), phosphorus (TP), total nitrogen (TN), Dissolved organic carbon (DOC), total organic carbon (TOC) and chlorophyll-a (Chl-a)).
4. Discussion
4.1. Prey Preference of Chironomidae Larvae
Our study provides scientific evidence that Chironomidae larvae have a significant preference (E’ > 0.5) for D. armatus, E. minima, and T. dimorphus, while showing negative selection for other species, even ones that are found in its gut (Table 2). These results coincide closely with those obtained in a tropic area by Henriques-Oliveira et al. [38] and Galizzi et al. [39], who reported that the Chironomidae family mainly consumes Bacillariophyta and Chlorophyta in the Tiradero River and Paraná River. We also found different patterns of OTUs’ relative abundance among the study sites (Table 1, Figure 2). Chironomidae showed a relatively high abundance for D. armatus in all study sites. We therefore suggest that Chironomidae prefers phytoplankton and that this preference is not affected by climate or natural environmental factors. However, differences in physio-chemical factors, such as water quality, nutrients, Chl-a, and carbon concentrations, caused by anthropogenic impacts (i.e., construction of large-scale weirs), particle size of prey species (small-sized single cell), and chemical factors (chemokinesis) can affect the feeding behavior of the 4th instar larvae of a Chironomidae. The diet preference of aquatic organisms hinders us in the application of environmental monitoring. Our results imply that application of NGS can be an alternative method to identify diet preferences of the macroinvertebrate organisms.
4.2. Efficiency of Meta-Barcoding for Analysis of Chironomidae Larvae Gut Contents
The DNA meta-barcoding approach for analyzing the gut contents of Chironomidae larvae have two advantages over other methods: (1) the size range of Chironomidae stages open to study has increased from adult to instar larvae and (2) prey identification to the species or genus level has become possible. Previously, small Chironomidae species stages, including instar larvae, could not be studied because their guts were too small to be examined by MI. Samples obtained from this larval stage of Chironomidae had to be analyzed using methods requiring great expertise. Therefore, despite the importance of assessing prey in terms of predator size [40], MI often ignored the larval stage of Chironomidae. However, if surgical evisceration of the gut is possible for both larvae and adults, their gut content analysis can be successfully carried out (Table 1 and Table 2); thus, a detailed understanding of the effects of Chironomidae populations on the biodiversity in an ecosystem can be based on analysis accounting for the size of species at all stages of growth [41]. The second advantage of DNA meta-barcoding is the high resolution of prey identification. MI is often impeded by the incomplete nature of the prey specimen, and digestion degrades prey specimens, resulting in identification failure. These problems can be overcome by DNA meta-barcoding. The high-resolution characterization of the food web structure is possible, and detailed remedial strategies for species management (of both predator and prey) can be achieved. Of course, the impact of predators on prey species should also be investigated quantitatively in conjunction with the qualitative identification of prey species using DNA meta-barcoding.
4.3. Potential Indicator of the Surrounding Environment
Interestingly, we found OTUs from H. labeo, which is a benthic fish, in Chironomidae individuals among all weirs (Table 1), even though Chironomidae cannot consume H. labeo directly. Chironomid species are known to eat castoff cells from benthic fish as a source of food. Some studies have shown that MI of the gut contents of aquatic organisms can be used to supplement the biodiversity inventories of benthic macroinvertebrates [42,43,44,45]. Few studies have examined the potential of using aquatic organisms in gut contents to monitor ecosystem function using the DNA meta-barcoding approach [46]. However, none of these studies have examined the effectiveness of the procedure for monitoring or evaluating biodiversity. DNA meta-barcoding not only provides dietary insights for estimating the impact of the food-web structure in large-scale weirs but also a tool to assess biological indicators. Additional experimental studies, based on the approach used in the present study, are necessary to develop a reliable biological indicator.
Author Contributions
Conceptualization, H.J. and I.-S.K.; methodology, H.J. and K.P.; formal analysis, H.J. and W.-S.K.; investigation, H.J. and W-S.K.; writing—original draft preparation, H.J. and I.-S.K.; writing—review and editing, B.C. and I.-S.K.; project administration, I.-S.K.; funding acquisition, I.-S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research Foundation of Korea, grant number NRF-2018R1A6A1A03024314.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
Detailed information of samples (the Gum River (SJ: Sejong Weir), Yeongsan River (JS: Juksan Weir), and Nakdong River (GG: Gangjeong-Goryeong Weir, DS: Dalseong Weir)).
Table A1.
Detailed information of samples (the Gum River (SJ: Sejong Weir), Yeongsan River (JS: Juksan Weir), and Nakdong River (GG: Gangjeong-Goryeong Weir, DS: Dalseong Weir)).
| Sample ID | Sampling Depth (m) | Sampling Date (Month) | Sampling Sites |
|---|---|---|---|
| SJ-A | 0.5 | July | Sejong Weir |
| SJ-B | 1 | June | Sejong Weir |
| JS-D | 0.5 | July | Juksan Weir |
| JS-E | 0.5 | July | Juksan Weir |
| GG-A | 0.5 | June | Gangjeong-Goryeong Weir |
| GG-D | 0.5 | June | Gangjeong-Goryeong Weir |
| DS-B | 0.5 | June | Dalseong Weir |
| DS-E | 0.5 | June | Dalseong Weir |
Table A2.
Sequence of the steps in the experiment.
Table A2.
Sequence of the steps in the experiment.
| Step | Details |
|---|---|
| Washing | Remove impurities from the larva surface |
| Weight measurement | Weigh with an electronic balance (SHIMADZU, Kyoto, Japan) |
| Total length measurement | Observe the larva using a dissecting microscope (LEICA LED2000, Wetzlar, Germany), and measure its length |
| Identification of larva stage | 1st, 2nd, 3rd, 4th of instar larva |
| Dissection | Grab the head of the larva and cut the tissue of the torso with a scalpel |
| DNA extraction | Samples of micro-tubes were extracted DNA by DNeasy Blood & Tissue Kit (Cat. No. 69504, Qiagen, Düsseldorf, Germany) |
References
- Armitage, P.D.; Pinder, L.C.; Cranston, P.S. (Eds.) The Chironomidae: Biology and Ecology of Non-Biting Midges; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Pinder, L.C.V. The habitats of chironomid larvae. In The Chironomidae; Springer: Dordrecht, The Netherlands, 1995; pp. 107–135. [Google Scholar]
- Walker, I.R. Midges: Chironomidae and related diptera. In Tracking Environmental Change Using Lake Sediments; Springer: Dordrecht, The Netherlands, 2001; pp. 43–66. [Google Scholar]
- Oliver, D.R.; Roussel, M.E. Redescription of Brillia Kieffer (Diptera: Chironomidae) with Descriptions of Nearctic Species. Can. Èntomol. 1983, 115, 257–279. [Google Scholar] [CrossRef]
- Wallace, J.B.; Merritt, R.W. Filter-Feeding Ecology of Aquatic Insects. Annu. Rev. Èntomol. 1980, 25, 103–132. [Google Scholar] [CrossRef]
- James, A.B.; Dewson, Z.S.; Death, R.G. The effect of experimental flow reductions on macroinvertebrate drift in natural and streamside channels. River Res. Appl. 2007, 24, 22–35. [Google Scholar] [CrossRef]
- Dézerald, O.; Leroy, C.; Corbara, B.; Carrias, J.-F.; Pélozuelo, L.; Dejean, A.; Céréghino, R. Food-Web Structure in Relation to Environmental Gradients and Predator-Prey Ratios in Tank-Bromeliad Ecosystems. PLoS ONE 2013, 8, e71735. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L. Local Distribution, Food Choice and Food Consumption of Diving Ducks on a South Swedish Lake. Oikos 1972, 23, 82. [Google Scholar] [CrossRef]
- Byeon, H.K. Population ecology of Squalidus japonicus coreanus (Cyprinidae) in the Namhan River, Korea. Korean, J. Environ. Ecol. 2012, 26, 367–373. [Google Scholar]
- Chae, Y.G. (2018). A study concerning legal aspects on the use of water acquired by South Korea’s Four Rivers Restoration Project. Korean Public Law Assoc. 2018, 46, 199–223. [Google Scholar]
- Jo, H.; Jeppesen, E.; Ventura, M.; Buchaca, T.; Gim, J.-S.; Yoon, J.-D.; Kim, N.-H.; Joo, G.-J. Responses of fish assemblage structure to large-scale weir construction in riverine ecosystems. Sci. Total. Environ. 2019, 657, 1334–1342. [Google Scholar] [CrossRef]
- Normile, D. Restoration or Devastation? Sci. 2010, 327, 1568–1570. [Google Scholar] [CrossRef]
- Canobbio, S.; Mezzanotte, V.; Sanfilippo, U.; Benvenuto, F. Effect of Multiple Stressors on Water Quality and Macroinvertebrate Assemblages in an Effluent-Dominated Stream. Water Air Soil Pollut. 2008, 198, 359–371. [Google Scholar] [CrossRef]
- De Oliveira, E.F.; Minte-Vera, C.V.; Goulart, E. Structure of fish assemblages along spatial gradients in a deep subtropical reservoir (Itaipu Reservoir, Brazil-Paraguay border). Environ. Boil. Fishes 2005, 72, 283–304. [Google Scholar] [CrossRef]
- Rosa, A.G.; Menezes, G.; Melo, O.; Pinho, M. Weight–length relationships of 33 demersal fish species from Azores archipelago. Fish. Res. 2006, 80, 329–332. [Google Scholar] [CrossRef]
- Kuch, M.; Steppan, S.; Rohland, N.; Betancourt, J.L.; Latorre, C.; Poinar, H.N. Molecular analysis of a 11 700-year-old rodent midden from the Atacama Desert, Chile. Mol. Ecol. 2002, 11, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.K.; Jo, H.; Park, J.W.; Chang, K.H.; Kwak, I.S. (2020) The gut content analysis of Polyperdium scalaenum in the large-scale weirs of 4 major river ecosystem. Korean J. Ecol. Environ. 2020, 53, 55–62. [Google Scholar]
- Oh, H.-J.; Krogh, P.H.; Jeong, H.-G.; Joo, G.-J.; Kwak, I.-S.; Hwang, S.-J.; Gim, J.-S.; Chang, K.-H.; Jo, H. Pretreatment Method for DNA Barcoding to Analyze Gut Contents of Rotifers. Appl. Sci. 2020, 10, 1064. [Google Scholar] [CrossRef]
- Blankenship, L.E.; Yayanos, A.A. Universal primers and PCR of gut contents to study marine invertebrate diets. Mol. Ecol. 2005, 14, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Durbin, E.; Casas, M.C.; Rynearson, T.A.; Smith, D.C. Measurement of copepod predation on nauplii using qPCR of the cytochrome oxidase I gene. Mar. Boil. 2007, 153, 699–707. [Google Scholar] [CrossRef]
- Nejstgaard, J.C.; Frischer, M.; Simonelli, P.; Troedsson, C.; Brakel, M.; Adiyaman, F.; Sazhin, A.F.; Artigas, L.F. Quantitative PCR to estimate copepod feeding. Mar. Boil. 2007, 153, 565–577. [Google Scholar] [CrossRef]
- Riemann, L.; Alfredsson, H.; Hansen, M.M.; Als, T.D.; Nielsen, T.G.; Munk, P.; Aarestrup, K.; Maes, G.; Sparholt, H.; Petersen, M.I.; et al. Qualitative assessment of the diet of European eel larvae in the Sargasso Sea resolved by DNA barcoding. Boil. Lett. 2010, 6, 819–822. [Google Scholar] [CrossRef]
- Cleary, A.; Durbin, E.; Rynearson, T. Krill feeding on sediment in the Gulf of Maine (North Atlantic). Mar. Ecol. Prog. Ser. 2012, 455, 157–172. [Google Scholar] [CrossRef]
- Durbin, E.; Casas, M.C.; Rynearson, T.A. Copepod feeding and digestion rates using prey DNA and qPCR. J. Plankton Res. 2011, 34, 72–82. [Google Scholar] [CrossRef]
- Carreon-Martinez, L.; Johnson, T.B.; Ludsin, S.A.; Heath, D.D. Utilization of stomach content DNA to determine diet diversity in piscivorous fishes. J. Fish Boil. 2011, 78, 1170–1182. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Gim, J.-A.; Jeong, K.-S.; Kim, H.-S.; Joo, G.-J. Application of DNA barcoding for identification of freshwater carnivorous fish diets: Is number of prey items dependent on size class for Micropterus salmoides? Ecol. Evol. 2013, 4, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Ventura, M.; Vidal, N.; Gim, J.; Buchaca, T.; Barmuta, L.; Jeppesen, E.; Joo, G.-J. Discovering hidden biodiversity: The use of complementary monitoring of fish diet based on DNA barcoding in freshwater ecosystems. Ecol. Evol. 2015, 6, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Andrew, R.L.; Bernatchez, L.; Bonin, A.; Buerkle, C.A.; Carstens, B.C.; Emerson, B.C.; Garant, D.; Giraud, T.; Kane, N.C.; Rogers, S.M.; et al. A road map for molecular ecology. Mol. Ecol. 2013, 22, 2605–2626. [Google Scholar] [CrossRef]
- De Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahé, F.; Logares, R.; Lara, E.; Berney, C.; Le Bescot, N.; Probert, I.; et al. Eukaryotic plankton diversity in the sunlit ocean. Science 2015, 348, 1261605. [Google Scholar] [CrossRef]
- Abad, D.; Albaina, A.; Rodrigo, M.A.; Laza-Martínez, A.; Uriarte, I.; Iriarte, A.; Villate, F.; Estonba, A. Is metabarcoding suitable for estuarine plankton monitoring? A comparative study with microscopy. Mar. Boil. 2016, 163, 149. [Google Scholar] [CrossRef]
- Wu, S.; Zhu, Z.; Fu, L.; Niu, B.; Li, W. WebMGA: A customizable web server for fast metagenomic sequence analysis. BMC Genomics 2011, 12, 444. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ivlev, V.S. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CT, USA, 1961. [Google Scholar]
- Jo, H.; Kim, D.-K.; Park, K.; Kwak, I.-S. Discrimination of Spatial Distribution of Aquatic Organisms in a Coastal Ecosystem Using eDNA. Appl. Sci. 2019, 9, 3450. [Google Scholar] [CrossRef]
- Henriques-Oliveira, A.L.; Nessimian, J.L.; Dorvillé, L.F.M. Feeding habits of chironomid larvae (Insecta: Diptera) from a stream in the Floresta da Tijuca, Rio de Janeiro, Brazil. Braz. J. Boil. 2003, 63, 269–281. [Google Scholar] [CrossRef]
- Galizzi, M.C.; Zilli, F.; Marchese, M.R. Diet and functional feeding groups of Chironomidae (Diptera) in the Middle Paraná River floodplain (Argentina). Iheringia. Série Zoologia 2012, 102, 117–121. [Google Scholar] [CrossRef]
- Huss, M.; Byström, P.; Persson, L. Resource heterogeneity, diet shifts and intra-cohort competition: Effects on size divergence in YOY fish. Oecologia 2008, 158, 249–257. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who is eating what: Diet assessment using next generation sequencing. Mol. Ecol. 2011, 21, 1931–1950. [Google Scholar] [CrossRef]
- Callisto, M.; Vono, V.; Barbosa, F.A.; Santeiro, S.M. Chironomidae as a food resource for Leporinus amblyrhynchus (Teleostei: Characiformes) and Pimelodus maculatus (Teleostei: Siluriformes) in a Brazilian reservoir. Lundiana 2002, 3, 67–73. [Google Scholar]
- Tupinambás, T.H.; Callisto, M.; Santos, G.B. Benthic macroinvertebrate assemblages structure in two headwater streams, south-eastern Brazil. Rev. Bras. Zoöl. 2007, 24, 887–897. [Google Scholar] [CrossRef]
- Maroneze, D.M.; Tupinambás, T.H.; Alves, C.B.M.; Vieira, F.; Pompeu, P.S.; Callisto, M. Fish as ecological tools to complement biodiversity inventories of benthic macroinvertebrates. Hydrobiologia 2011, 673, 29–40. [Google Scholar] [CrossRef]
- Cook, A.; Bundy, A. Use of fishes as sampling tools for understanding biodiversity and ecosystem functioning in the ocean. Mar. Ecol. Prog. Ser. 2012, 454, 1–18. [Google Scholar] [CrossRef]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zoöl. 2013, 10, 34. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).