Specific NDM-1 Inhibitor of Isoliquiritin Enhances the Activity of Meropenem against NDM-1-positive Enterobacteriaceae in vitro


Abstract
1. Introduction
2. Materials and Methods
2.1. Plasmid Construction and Protein Purification
2.2. Enzyme Inhibition Assays
2.3. Western Blot Assay
2.4. Minimal Inhibit Concentration (MIC) Assays
2.5. Growth Curves and Time-kill Assays
2.6. Statistical Analysis
3. Results
3.1. Isoliquiritin Significantly Inhibits the Activity of NDM-1
3.2. Isoliquiritin Has No Influence on the Growth of NDM-1-Positive Bacteria
3.3. Isoliquiritin Restores the Susceptibility of E. coli ZC-YN3 to Meropenem
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, T.Z.; Kodiyanplakkal, R.P.L.; Calfee, D.P. Antimicrobial resistance in nephrology. Nat. Rev. Nephrol. 2019, 15, 463–481. [Google Scholar] [CrossRef] [PubMed]
- WHO. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Microbiol. 2019, 17, 141–155. [Google Scholar] [CrossRef]
- King, A.M.; Reid-Yu, S.A.; Wang, W.L.; King, D.T.; De Pascale, G.; Strynadka, N.C.; Walsh, T.R.; Coombes, B.K.; Wright, G.D. Aspergillomarasmine a overcomes metallo-beta-lactamase antibiotic resistance. Nature 2014, 510, 503. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of ndm-1 positive bacteria in the New Delhi environment and its implications for human health: An environmental point prevalence study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Bush, K.; Courvalin, P.; Dantas, G.; Davies, J.; Eisenstein, B.; Huovinen, P.; Jacoby, G.A.; Kishony, R.; Kreiswirth, B.N.; Kutter, E.; et al. Tackling antibiotic resistance. Nat. Rev. Microbiol. 2011, 9, 894–896. [Google Scholar] [CrossRef]
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, Genetics and Worldwide Spread of New Delhi Metallo-β-lactamase (NDM): A threat to public health. BMC Microbiol. 2017, 17, 101. [Google Scholar] [CrossRef]
- Rehman, M.T.; AlAjmi, M.F.; Hussain, A.; Rather, G.M.; Khan, M.A. High-Throughput Virtual Screening, Molecular Dynamics Simulation, and Enzyme Kinetics Identified ZINC84525623 as a Potential Inhibitor of NDM-1. Int. J. Mol. Sci. 2019, 20, 819. [Google Scholar] [CrossRef]
- Linciano, P.; Cendron, L.; Gianquinto, E.; Spyrakis, F.; Tondi, D. Ten Years with New Delhi Metallo-β-lactamase-1 (NDM-1): From Structural Insights to Inhibitor Design. ACS Infect. Dis. 2019, 5, 9–34. [Google Scholar] [CrossRef]
- Jaramillo, M.C.; Mora, C.; Vélez, L.E.; Quijano, J. Kinetic and theoretical study of the chalcones as inhibitors of beta-lactamase enzyme. Med. Chem. 2009, 5, 434–439. [Google Scholar] [CrossRef]
- Liu, B.; Trout, R.E.L.; Chu, G.H.; McGarry, D.; Jackson, R.W.; Hamrick, J.C.; Daigle, D.M.; Cusick, S.M.; Pozzi, C.; De Luca, F.; et al. Discovery of Taniborbactam (VNRX-5133): A Broad-Spectrum Serine- and Metallo-β-lactamase Inhibitor for Carbapenem-Resistant Bacterial Infections. J. Med. Chem. 2019. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, S.; Wang, T.; Li, H.; Tang, S.; Wang, J.; Wang, Y.; Deng, X. Pterostilbene, a Potential MCR-1 Inhibitor That Enhances the Efficacy of Polymyxin B. Antimicrob. Agents Chemother. 2018, 62, e02146-17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhao, S.; Yang, S.; Lin, X.; He, X.; Wei, X.; Song, Q.; Li, R.; Fu, C.; Zhang, J.; et al. An “essential herbal medicine”-licorice: A review of phytochemicals and its effects in combination preparations. J. Ethnopharmacol. 2020, 249, 112439. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Kaur, S.; Kumar, N.; Singh, B.; Kumar, S. Evaluation of antigenotoxic activity of isoliquiritin apioside from glycyrrhiza glabra l. Toxicol. In Vitro 2009, 23, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Ma, J.Y. Isoliquiritin apioside suppresses in vitro invasiveness and angiogenesis of cancer cells and endothelial cells. Front. Pharmacol. 2018, 9, 1455. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.Z.; Li, X.; Gong, W.X.; Tian, J.S.; Gao, X.X.; Gao, L.; Zhang, X.; Du, G.H.; Qin, X.M. Protective effect of isoliquiritin against corticosterone-induced neurotoxicity in pc12 cells. Food Funct. 2017, 8, 1235–1244. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Ho, W.S. Combination of liquiritin, isoliquiritin and isoliquirigenin induce apoptotic cell death through upregulating p53 and p21 in the a549 non-small cell lung cancer cells. Oncol. Rep. 2014, 31, 298–304. [Google Scholar] [CrossRef]
- Yin, L.; Guan, E.S.; Zhang, Y.B.; Shu, Z.H.; Wang, B.; Wu, X.L.; Chen, J.; Liu, J.X.; Fu, X.Y.; Sun, W.H.; et al. Chemical profile and anti-inflammatory activity of total flavonoids from glycyrrhiza uralensis fisch. Iran. J. Pharm. Res. 2018, 17, 726–734. [Google Scholar]
- Liu, S.; Zhang, J.; Zhou, Y.; Hu, N.; Li, J.; Wang, Y.; Niu, X.; Deng, X.; Wang, J. Pterostilbene restores carbapenem susceptibility in ndm-producing isolates via inhibiting the activity of ndm enzymes. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Teng, Z.; Guo, Y.; Liu, X.; Zhang, J.; Niu, X.; Yu, Q.; Deng, X.; Wang, J. Theaflavin-3,3 -digallate increases the antibacterial activity of beta-lactam antibiotics by inhibiting metallo-beta-lactamase activity. J. Cell. Mol. Med. 2019. [Google Scholar] [CrossRef]
- Liu, S.; Zhou, Y.; Niu, X.; Wang, T.; Li, J.; Liu, Z.; Wang, J.; Tang, S.; Wang, Y.; Deng, X. Magnolol restores the activity of meropenem against ndm-1-producing Escherichia coli by inhibiting the activity of metallo-beta-lactamase. Cell. Death Discov. 2018, 4, 28. [Google Scholar] [CrossRef]
- Igawa, G.; Casey, M.; Sawabe, E.; Nukui, Y.; Okugawa, S.; Moriya, K.; Koike, R.; Tohda, S.; Saito, R. Comparison of agar dilution and broth microdilution methods for clostridium difficile antimicrobial susceptibility testing. J. Glob. Antimicrob. Resist. 2016, 7, 43–45. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.P.; Woodford, N. Global spread of antibiotic resistance: The example of New Delhi metallo-beta-lactamase (ndm)-mediated carbapenem resistance. J. Med. Microbiol. 2013, 62, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A. molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Delgado-Blas, J.F.; Ovejero, C.M.; Abadia-Patino, L.; Gonzalez-Zorn, B. Coexistence of mcr-1 and blandm-1 in Escherichia coli from Venezuela. Antimicrob. Agents Chemother. 2016, 60, 6356–6358. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Interplay between β-lactamases and new β-lactamase inhibitors. Nat. Rev. Microbiol. 2019, 17, 295–306. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, C.; Wang, Q.; Wang, Z.; Liang, X.; Zhang, F.; Zhang, Y.; Meng, H.; Chen, H.; Li, S.; et al. In vitro activity of the novel β-lactamase inhibitor taniborbactam (VNRX-5133), in combination with cefepime or meropenem, against MDR Gram-negative bacterial isolates from China. J. Antimicrob. Chemother. 2020. [Google Scholar] [CrossRef]
- Hamrick, J.C.; Docquier, J.D.; Uehara, T.; Myers, C.L.; Six, D.A.; Chatwin, C.L.; John, K.J.; Vernacchio, S.F.; Cusick, S.M.; Trout, R.E.L.; et al. VNRX-5133 (Taniborbactam), a Broad-Spectrum Inhibitor of Serine- and Metallo-β-Lactamases, Restores Activity of Cefepime in Enterobacterales and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2020, 64, e01963-19. [Google Scholar] [CrossRef]
- Tsivkovski, R.; Totrov, M.; Lomovskaya, O. Biochemical Characterization of QPX7728, a New Ultra-Broad-Spectrum Beta-lactamase Inhibitor of Serine and Metallo-Beta-Lactamases. Antimicrob. Agents Chemother. 2020. [Google Scholar] [CrossRef]
- Hecker, S.J.; Reddy, K.R.; Lomovskaya, O.; Griffith, D.C.; Rubio-Aparicio, D.; Nelson, K.; Tsivkovski, R.; Sun, D.; Sabet, M.; Tarazi, Z.; et al. Discovery of Cyclic Boronic Acid QPX7728, an Ultra-broad-spectrum Inhibitor of Serine and Metallo Beta-lactamases. J. Med. Chem. 2020. [Google Scholar] [CrossRef]
- Ko, H.C.; Wei, B.L.; Chiou, W.F. The effect of medicinal plants used in Chinese folk medicine on rantes secretion by virus-infected human epithelial cells. J. Ethnopharmacol. 2006, 107, 205–210. [Google Scholar] [CrossRef]
- Huang, X.L.; Wang, S.Y.; Wang, L.; Wang, H.; Li, X.; Cui, D.G. Administration of an herbal powder based on traditional Chinese veterinary medicine enhanced the fertility of holstein dairy cows affected with retained placenta. Theriogenology 2018, 121, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Lim, S.L.; Lu, K.H.; Sheen, L.Y. Antidepressant-like effects of gan-mai-dazao-tang via monoamine regulatory pathways on forced swimming test in rats. J. Tradit. Complement. Med. 2018, 8, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Kitano, M.; Kishida, H.; Hidaka, T.; Nabae, K.; Kawabe, M.; Hosoe, K. 90-day repeated-dose toxicity study of licorice flavonoid oil (lfo) in rats. Food Chem. Toxicol. 2008, 46, 2349–2357. [Google Scholar] [CrossRef] [PubMed]
- Nazari, S.; Rameshrad, M.; Hosseinzadeh, H. Toxicological effects of glycyrrhiza glabra (licorice): A. review. Phytother. Res. 2017, 31, 1635–1650. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, X.; Xu, R.; Zhang, S. Renoprotective Effects of Isoliquiritin Against Cationic Bovine Serum Albumin-Induced Membranous Glomerulonephritis in Experimental Rat Model Through Its Anti-Oxidative and Anti-Inflammatory Properties. Drug Des. Dev. Ther. 2019, 13, 3735–3751. [Google Scholar] [CrossRef]




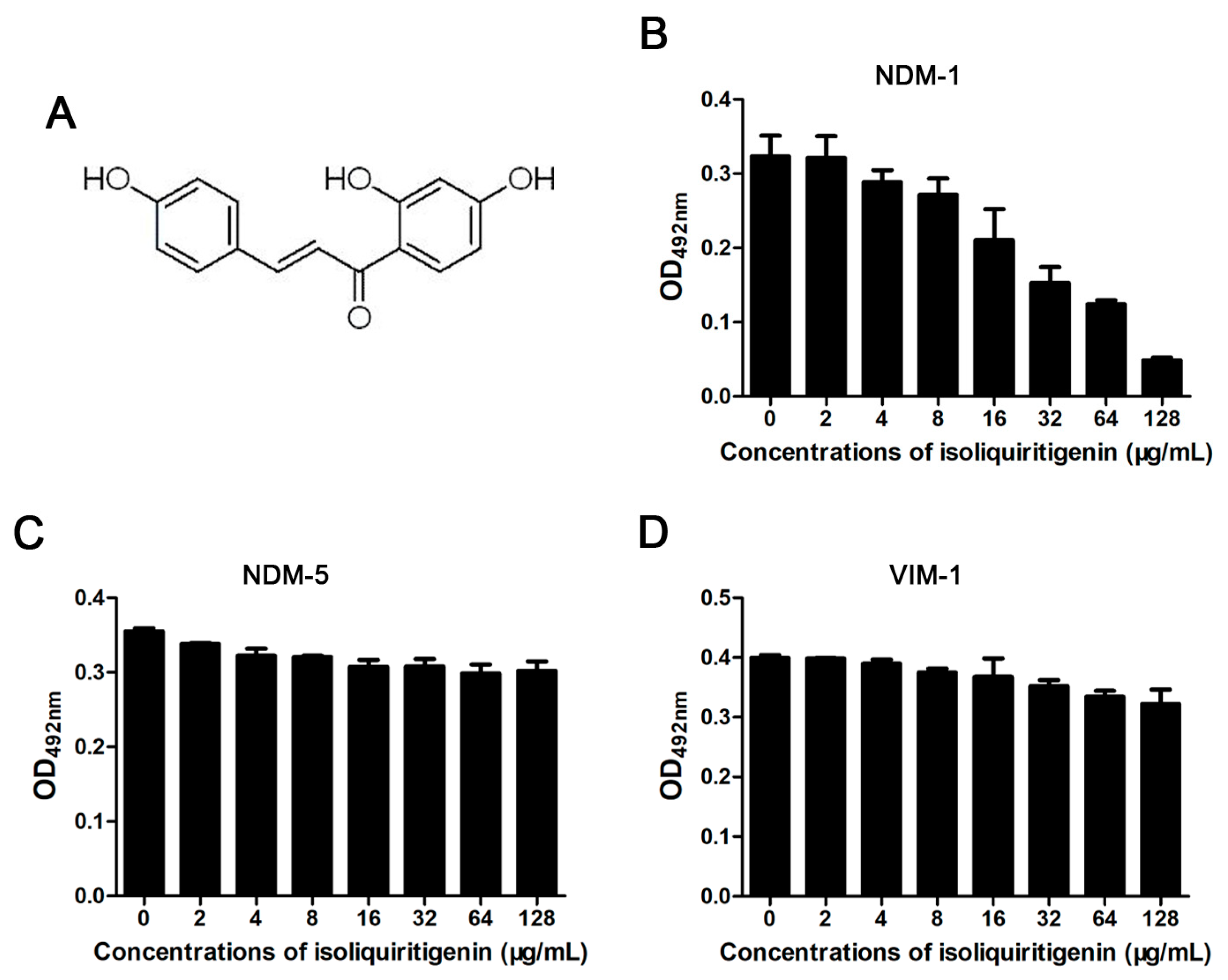{kind=link}
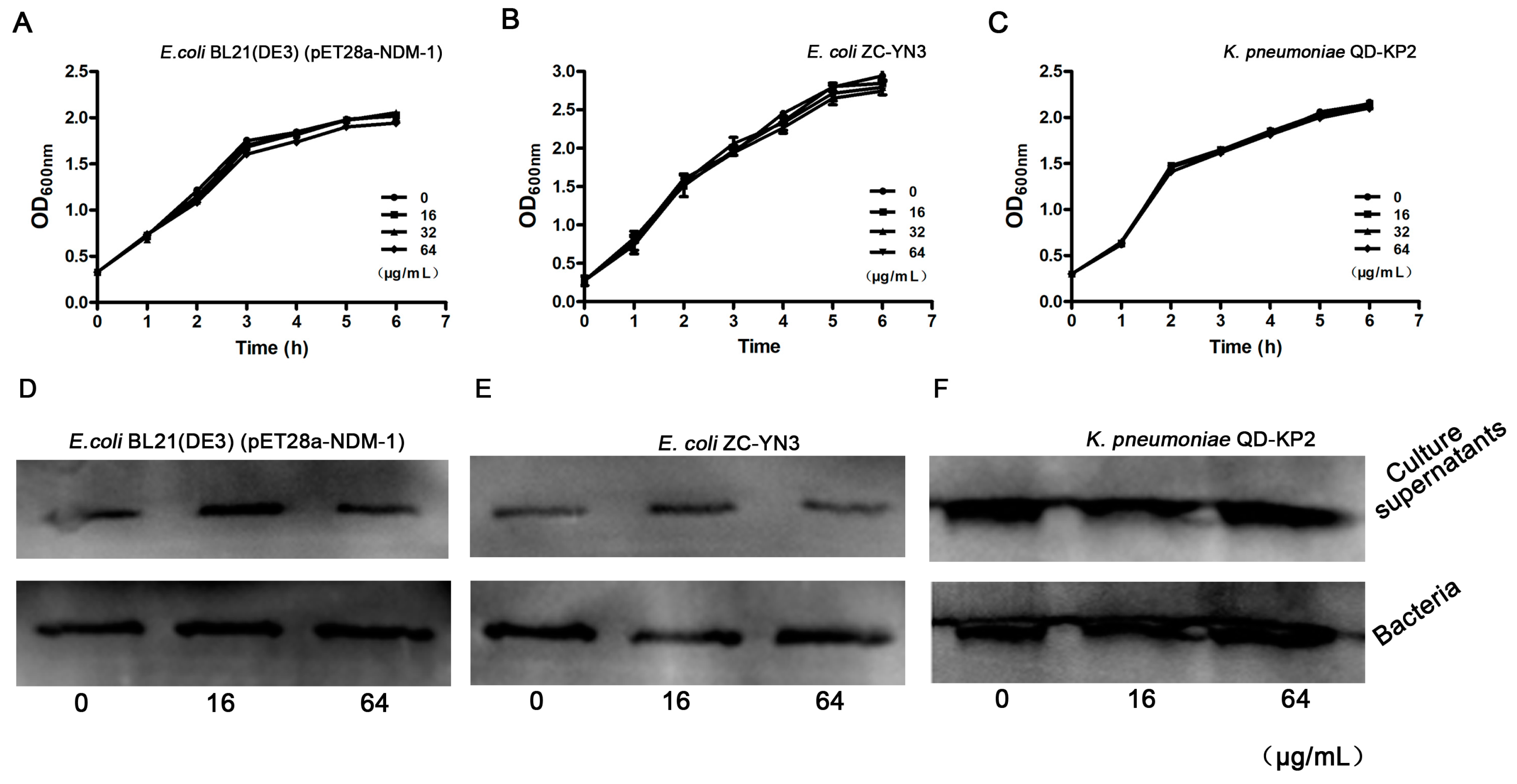{kind=link}
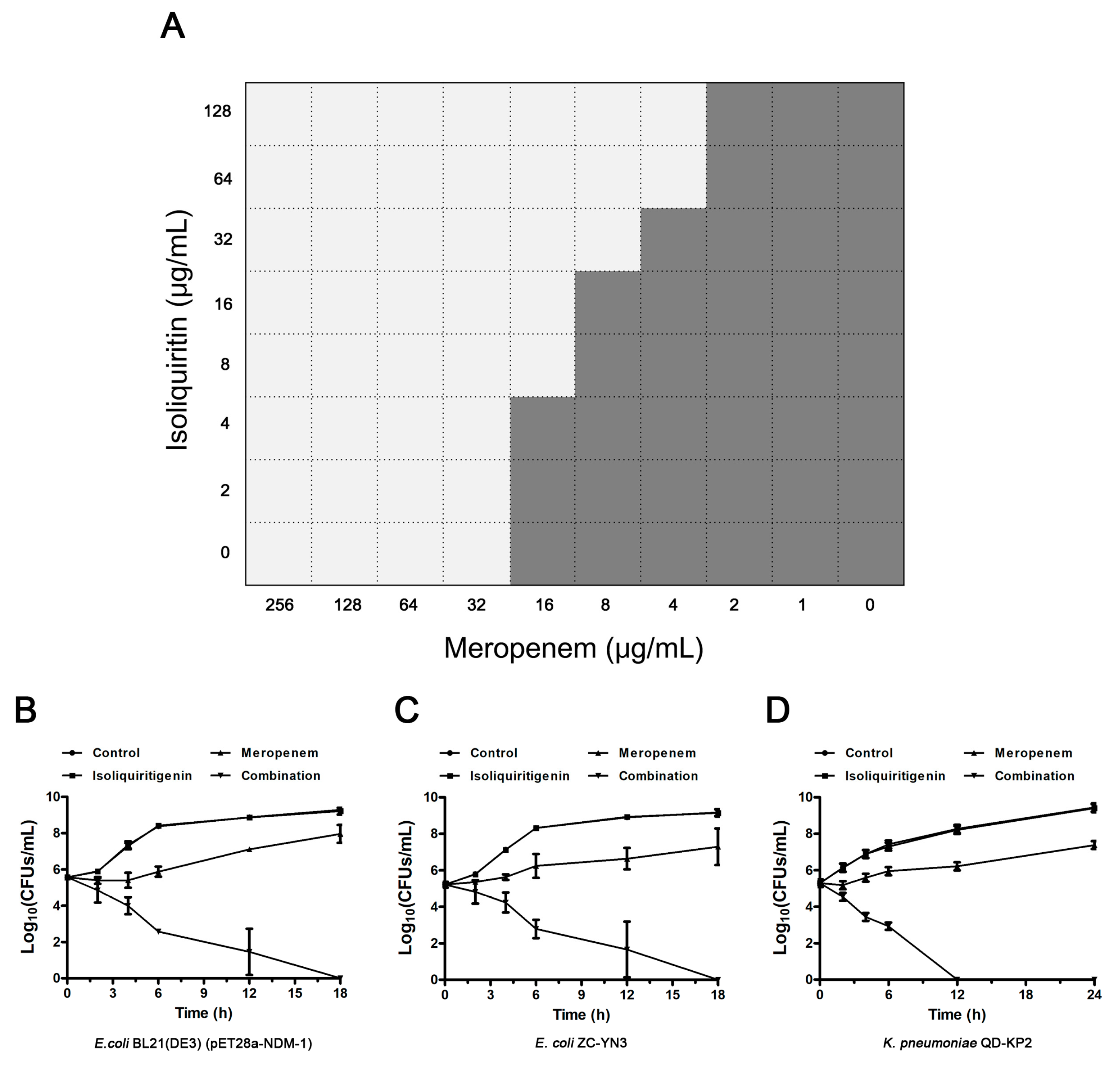{kind=link}
| Primers Name | Oligonucleotide Primer Sequence (5’–3’) |
|---|---|
| NDM-1-F | CTGGGATCCatggaattgcccaatattatg |
| NDM-1-R | CTGCTCGAGtcagcgcagcttgtcgg |
| NDM-5-F | CTGGGATCCatggaattgcccaatattatg |
| NDM-5-R | CTGCTCGAGtcagcgcagcttgtcgg |
| VIM-1-F | CTGGGATCCatgttaaaagttattagtag |
| VIM-1-R | CTGCTCGAGctactcggcgactgag |
| Strains | Meropenem | Combination (reduction fold) | FIC Index |
|---|---|---|---|
| E. coli ZJ487 | 32 | 8 (4) | 0.375 1 |
| E. coli ZC-YN3 | 64 | 4 (16) | 0.312 1 |
| K. pneumoniae QD-KP2 | 64 | 8 (8) | 0.375 1 |
| E. coli ZC-YN5 | 32 | 16 (2) | 0.75 2 |
| E. coli BL21(DE3) (pET28a-NDM-1) | 32 | 4 (8) | 0.375 1 |
| E. coli BL21(DE3) (pET28a) | 0.008 | 0.008 (0) | 1 2 |
| E. coli ATCC25922 | 0.031 | 0.031 (0) | 1.25 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Sun, X.; Kong, F.; Xia, L.; Deng, X.; Wang, D.; Wang, J. Specific NDM-1 Inhibitor of Isoliquiritin Enhances the Activity of Meropenem against NDM-1-positive Enterobacteriaceae in vitro. Int. J. Environ. Res. Public Health 2020, 17, 2162. https://doi.org/10.3390/ijerph17062162
Wang Y, Sun X, Kong F, Xia L, Deng X, Wang D, Wang J. Specific NDM-1 Inhibitor of Isoliquiritin Enhances the Activity of Meropenem against NDM-1-positive Enterobacteriaceae in vitro. International Journal of Environmental Research and Public Health. 2020; 17(6):2162. https://doi.org/10.3390/ijerph17062162
Chicago/Turabian StyleWang, Yanling, Xiaodi Sun, Fanrong Kong, Lining Xia, Xuming Deng, Dacheng Wang, and Jianfeng Wang. 2020. "Specific NDM-1 Inhibitor of Isoliquiritin Enhances the Activity of Meropenem against NDM-1-positive Enterobacteriaceae in vitro" International Journal of Environmental Research and Public Health 17, no. 6: 2162. https://doi.org/10.3390/ijerph17062162
APA StyleWang, Y., Sun, X., Kong, F., Xia, L., Deng, X., Wang, D., & Wang, J. (2020). Specific NDM-1 Inhibitor of Isoliquiritin Enhances the Activity of Meropenem against NDM-1-positive Enterobacteriaceae in vitro. International Journal of Environmental Research and Public Health, 17(6), 2162. https://doi.org/10.3390/ijerph17062162