Quercetin and Its Mixture Increase the Stress Resistance of Caenorhabditis elegans to UV-B



Abstract
1. Introduction
2. Materials and Methods
2.1. Strain, Cultivation and Grouping of C. elegans
2.2. UV-B Irradiation
2.3. Photographing of C. elegans
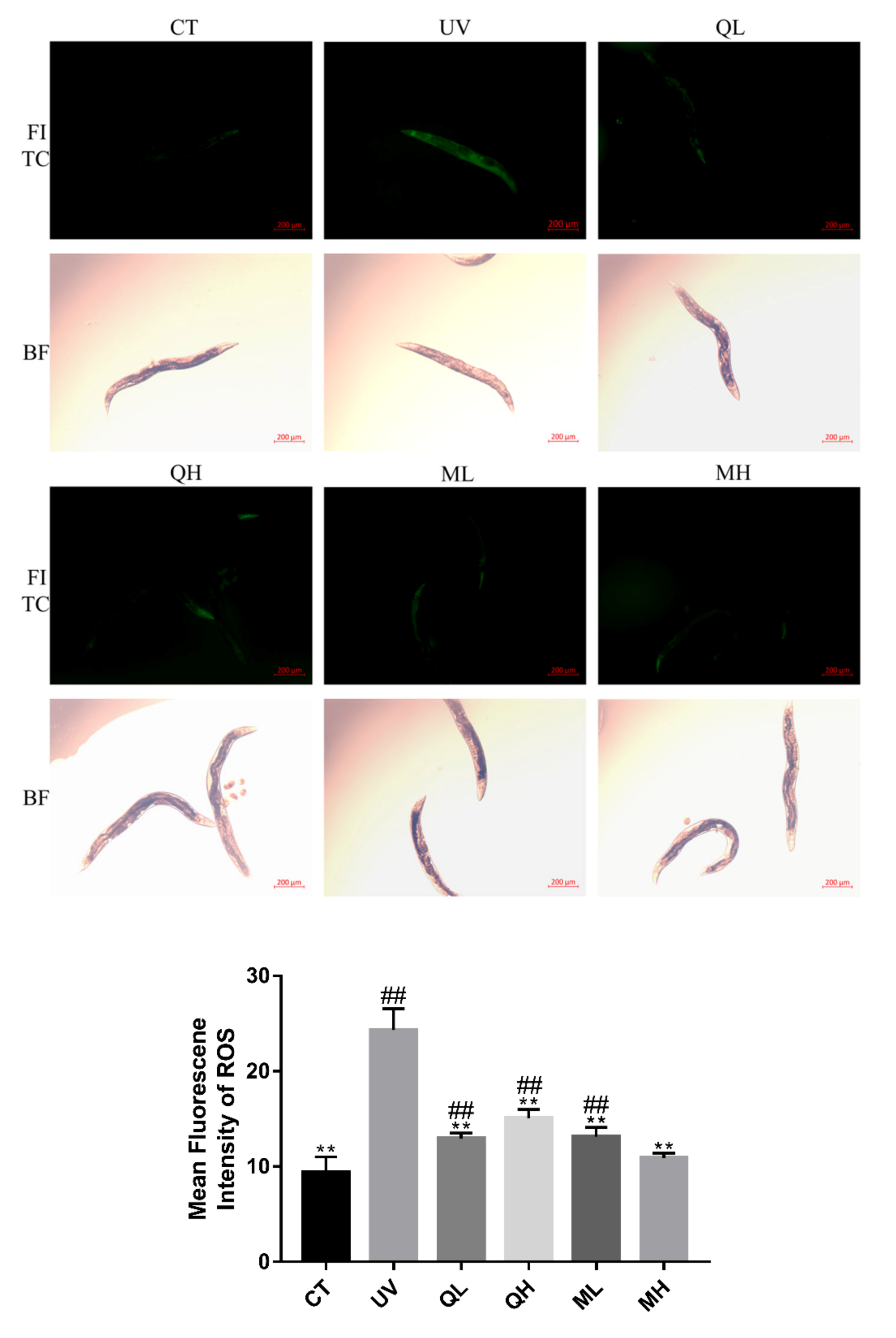
2.4. ROS Level Assay
2.5. Lifespan Test
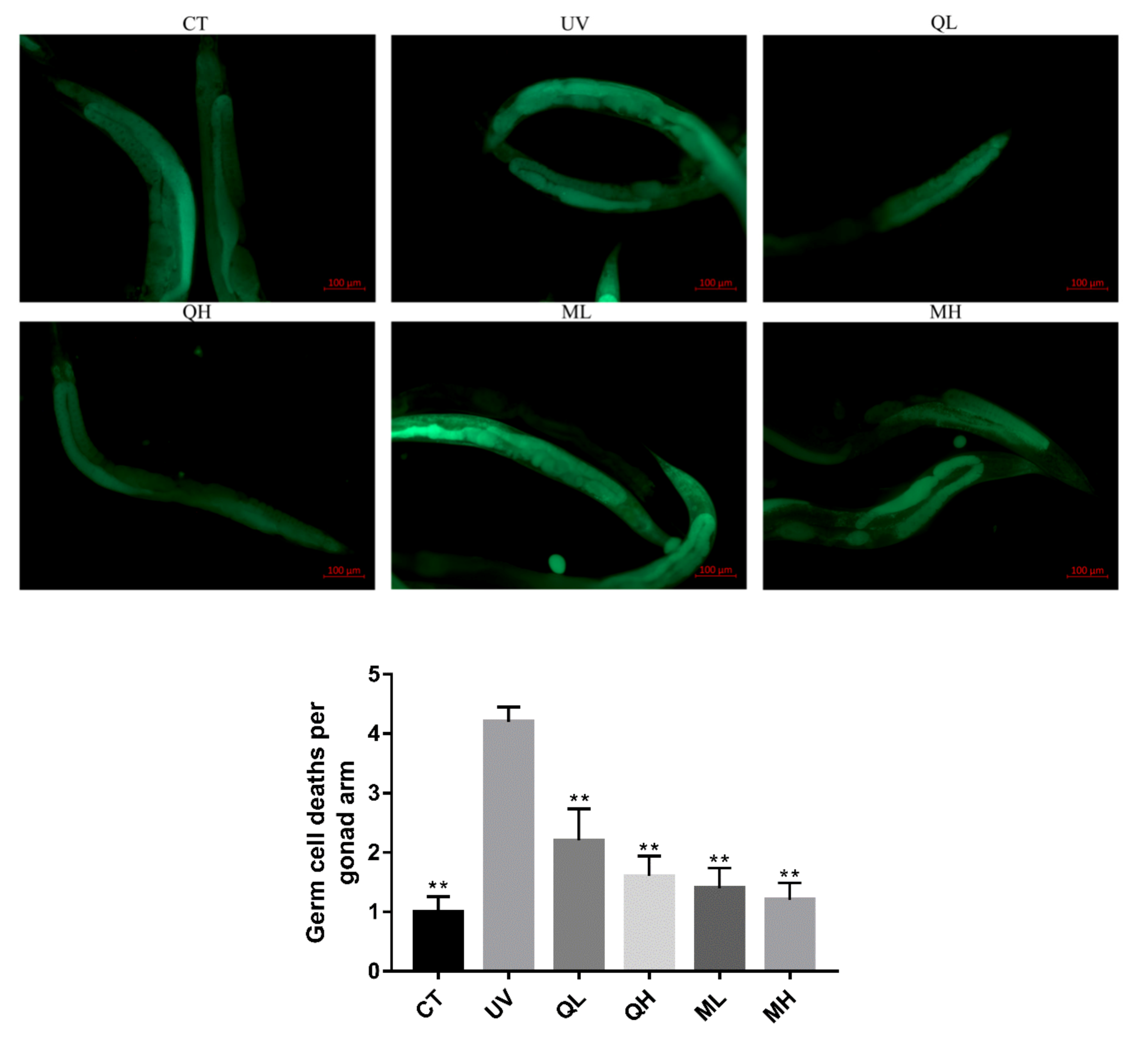
2.6. Gonadal Cell Apoptosis Test
2.7. Embryo Lethality Test
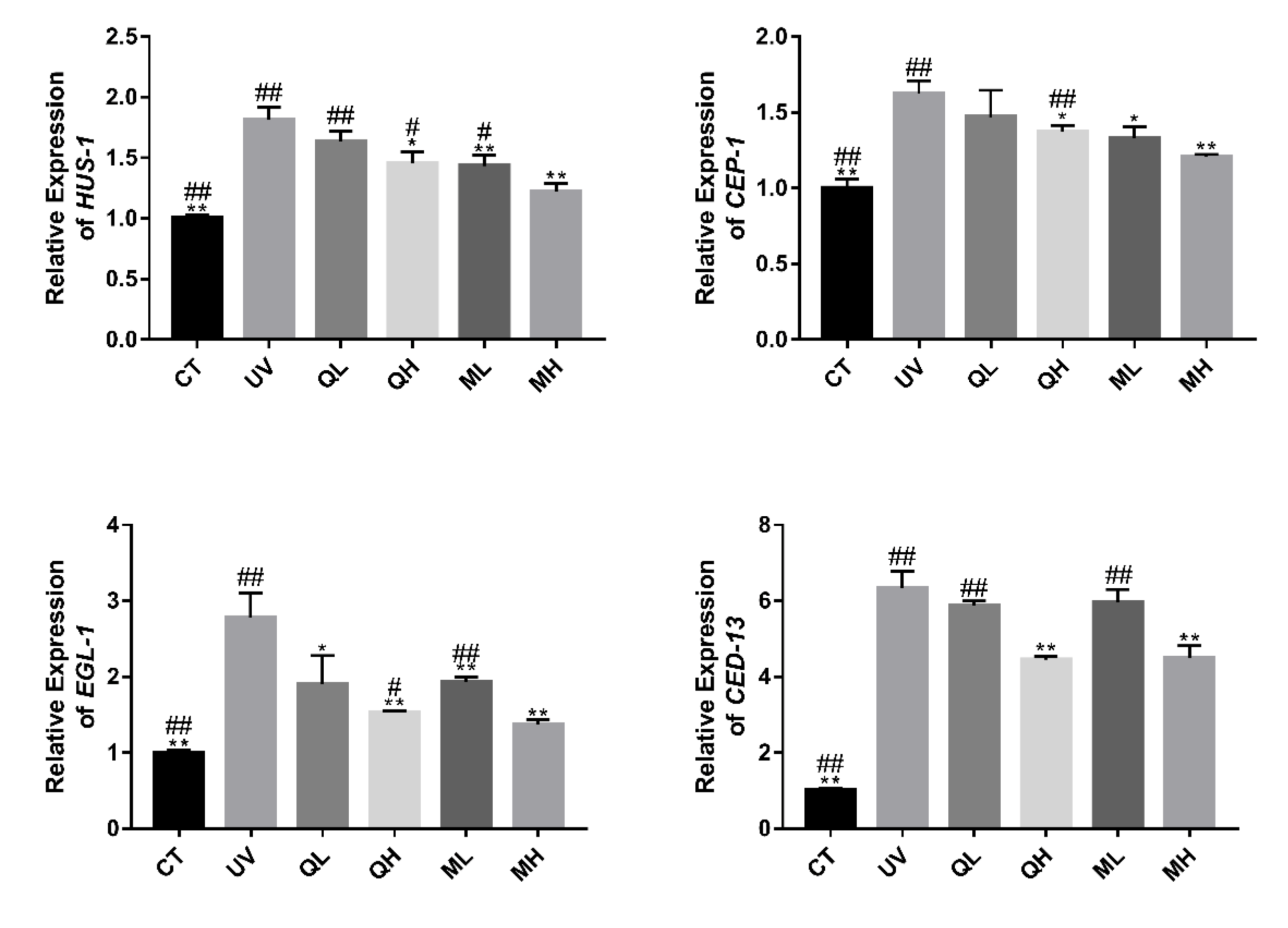
2.8. RT-qPCR Analysis of Transcription Levels of HUS-1, CEP-1, EGL-1, and CED-13
2.9. Statistical Analysis
3. Results
3.1. Initial Evaluation of Different Drugs on the Resistance to UV-B Stress by ROS Assay
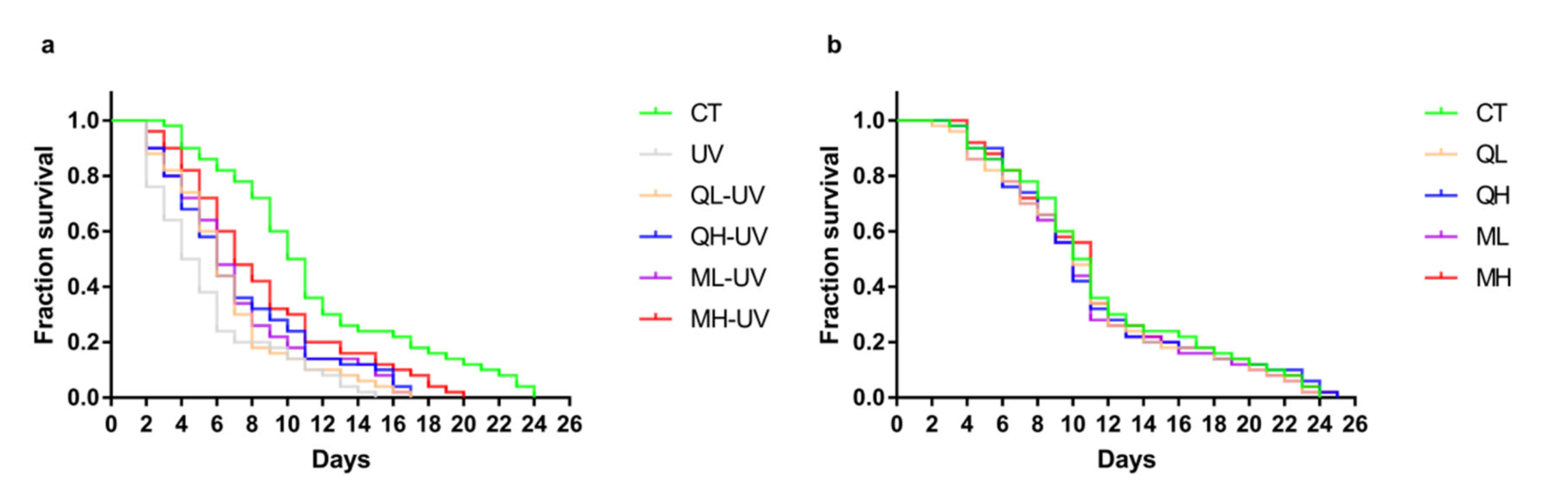
3.2. Quercetin and Its Mixture Helped C. elegans Resist UV-B Stress
3.3. Quercetin and Its Mixture Inhibit UV-B-Induced Increase in ROS Levels
3.4. Quercetin and Its Mixture Decreased DNA Damage of Germ Cells Caused by UV-B
3.5. Regulation of Quercetin and Its Mixture on Apoptosis Related Genes in C. elegans
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| UV | ultraviolet |
| C. elegans | Caenorhabditis elegans |
| CPD | cyclobutane pyrimidine dimer |
| NER | nucleotide excision repair |
| ROS | reactive oxygen species |
| RT-qPCR | quantitative real time polymerase chain reaction |
| NGM | nematode growth medium |
| CT | control |
| Q | quercetin |
| Lu | luteolin |
| Ly | lycopene |
| MIX | mixture |
| QL | quercetin low dose |
| QH | quercetin high dose |
| ML | mixture low dose |
| MH | mixture high dose |
| DCF | dichlorofluorescein |
| AO | acridine orange |
| FITC | fluorescein isothiocyanate isomer |
| MFI | mean fluorescence intensity |
| BF | bright field. |
References
- Stergiou, L.; Doukoumetzidis, K.; Sendoel, A.; Hengartner, M.O. The nucleotide excision repair pathway is required for UV-C-induced apoptosis in Caenorhabditis elegans. Cell Death Differ. 2007, 14, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Hader, D.P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Artal-Sanz, M.; Tavernarakis, N. Prohibitin couples diapause signalling to mitochondrial metabolism during ageing in C. elegans. Nature 2009, 461, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Bian, P.; Liang, J.; Wang, Y.; Li, L.; Wang, J.; Yuan, H.; Chen, S.; Xu, A.; Wu, L. Synergistic Effects Induced by a Low Dose of Diesel Particulate Extract and Ultraviolet-A in Caenorhabditis elegans: DNA Damage-Triggered Germ Cell Apoptosis. Chem. Res. Toxicol. 2014, 27, 990–1001. [Google Scholar] [CrossRef]
- De Gruijl, F.R. Photocarcinogenesis: UVA vs. UVB Radiation. Skin Pharmacol. Physiol. 2002, 15, 316–320. [Google Scholar] [CrossRef]
- Patt, H.M.; Tyeee, E.B.; Straube, R.L.; Smith, D.E. Cysteine protection against X irradiation. Science 1949, 110, 213–214. [Google Scholar] [CrossRef]
- Mathews-Roth, M.M.; Krinsky, N.I. Carotenoid Dose Level And Protection Against Uv-B Induced Skin Tumors. Photochem. Photobiol. 1985, 42, 35–38. [Google Scholar] [CrossRef]
- Beggs, C.J.; Schneider-Ziebert, U.; Wellmann, E. UV-B Radiation and Adaptive Mechanisms in Plants; Springer: Berlin/Heidelberg, Germany, 1986; pp. 235–250. [Google Scholar]
- Weiss, J.F.; Landauer, M.R. Protection against ionizing radiation by antioxidant nutrients and phytochemicals. Toxicology 2003, 189, 1–20. [Google Scholar] [CrossRef]
- Sitailo, L.A.; Tibudan, S.S.; Denning, M.F. Activation of Caspase-9 Is Required for UV-induced Apoptosis of Human Keratinocytes. J. Biol. Chem. 2002, 277, 19346–19352. [Google Scholar] [CrossRef]
- Derry, W.B.; Putzke, A.P.; Rothman, J.H. Caenorhabditis elegans p53: Role in Apoptosis, Meiosis, and Stress Resistance. Science 2001, 294, 591–595. [Google Scholar] [CrossRef]
- Hofmann, E.R.; Milstein, S.; Boulton, S.J.; Ye, M.; Hofmann, J.J.; Stergiou, L.; Gartner, A.; Vidal, M.; Hengartner, M.O. Caenorhabditis elegans HUS-1 Is a DNA Damage Checkpoint Protein Required for Genome Stability and EGL-1-Mediated Apoptosis. Curr. Biol. 2002, 12, 1908–1918. [Google Scholar] [CrossRef]
- Fairlie, W.D.; Perugini, M.A.; Kvansakul, M.; Chen, L.; Huang, D.C.S.; Colman, P.M. CED-4 forms a 2:2 heterotetrameric complex with CED-9 until specifically displaced by EGL-1 or CED-13. Cell Death Differ. 2006, 13, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, J.; Liu, L.; Sharma, S.; Dong, Q. Quercetin potentiates doxorubicin mediated antitumor effects against liver cancer through p53/Bcl-xl. PLoS ONE 2012, 7, e51764. [Google Scholar] [CrossRef] [PubMed]
- Erden Inal, M.; Kahraman, A. The protective effect of flavonol quercetin against ultraviolet an induced oxidative stress in rats1This research was supported by Osmangazi University Eskisehir-Turkey1. Toxicology 2000, 154, 21–29. [Google Scholar] [CrossRef]
- Kasala, E.R.; Bodduluru, L.N.; Barua, C.C.; Gogoi, R. Antioxidant and antitumor efficacy of Luteolin, a dietary flavone on benzo(a)pyrene-induced experimental lung carcinogenesis. Biomed. Pharmacother. 2016, 82, 568–577. [Google Scholar] [CrossRef]
- Kelkel, M.; Schumacher, M.; Dicato, M.; Diederich, M. Antioxidant and anti-proliferative properties of lycopene. Free Radical Res. 2011, 45, 925–940. [Google Scholar] [CrossRef]
- Brenner, S. The Genetics of Caenorhabditis Elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Au-Porta-de-la-Riva, M.; Au-Fontrodona, L.; Au-Villanueva, A.; Au-Cerón, J. Basic Caenorhabditis elegans Methods: Synchronization and Observation. JoVE 2012. [Google Scholar] [CrossRef]
- Gumienny, T.L.; Lambie, E.; Hartwieg, E.; Horvitz, H.R.; Hengartner, M.O. Genetic control of programmed cell death in the Caenorhabditis elegans hermaphrodite germline. Development 1999, 126, 1011. [Google Scholar] [PubMed]
- Murakami, S.; Johnson, T.E. A Genetic Pathway Conferring Life Extension and Resistance to UV Stress in Caenorhabditis elegans. Genetics 1996, 143, 1207–1218. [Google Scholar]
- Lettre, G.; Kritikou, E.A.; Jaeggi, M.; Calixto, A.; Fraser, A.G.; Kamath, R.S.; Ahringer, J.; Hengartner, M.O. Genome-wide RNAi identifies p53-dependent and -independent regulators of germ cell apoptosis in C. elegans. Cell Death Differ. 2004, 11, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhang, M.; Zheng, L.; Liang, Q.; Li, H.; Chen, J.-T.; Guo, H.; Oshina, S.Y.; Chen, Y.-Z.; Zhao, X.; et al. Cysteine protease cathepsin B mediates radiation-induced bystander effects. Nature 2017, 547, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Ly, K.; Reid, S.J.; Snell, R.G. Rapid RNA analysis of individual Caenorhabditis elegans. MethodsX 2015, 2, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F.; Canonico, B.; Curci, R.; Battistelli, M.; Mannello, F.; Papa, S.; Tarzia, G.; Falcieri, E. Melatonin prevents apoptosis induced by UV-B treatment in U937 cell line. J. Pineal Res. 2006, 40, 158–167. [Google Scholar] [CrossRef]
- Simon, H.-U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-J.; Jeng, J.-Y.; Lin, C.-W.; Wu, C.-Y.; Chen, Y.-C. Quercetin inhibition of ROS-dependent and -independent apoptosis in rat glioma C6 cells. Toxicology 2006, 223, 113–126. [Google Scholar] [CrossRef]
- Kampkötter, A.; Nkwonkam, C.G.; Zurawski, R.F.; Timpel, C.; Chovolou, Y.; Wätjen, W.; Kahl, R. Investigations of protective effects of the flavonoids quercetin and rutin on stress resistance in the model organism Caenorhabditis elegans. Toxicology 2007, 234, 113–123. [Google Scholar] [CrossRef]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Kulms, D.; Schwarz, T. Molecular mechanisms of UV-induced apoptosis. Photodermatol. Photoimmunol. Photomed. 2000, 16, 195–201. [Google Scholar] [CrossRef]
- Nehme, R.; Conradt, B. egl-1: A key activator of apoptotic cell death in C. elegans. Oncogene 2008, 27, S30–S40. [Google Scholar] [CrossRef]
- Schumacher, B.; Schertel, C.; Wittenburg, N.; Tuck, S.; Mitani, S.; Gartner, A.; Conradt, B.; Shaham, S. C. elegans ced-13 can promote apoptosis and is induced in response to DNA damage. Cell Death Differ. 2005, 12, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, B.; Hanazawa, M.; Lee, M.-H.; Nayak, S.; Volkmann, K.; Hofmann, R.; Hengartner, M.; Schedl, T.; Gartner, A. Translational Repression of C. elegans p53 by GLD-1 Regulates DNA Damage-Induced Apoptosis. Cell 2005, 120, 357–368. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| ACT-1 | GCTCTTGCCCCATCAACCAT | AGAAAGCTGGTGGTGACGAT |
| HUS-1 | GGCAATCGACGTGTTTATCAAAAT | TCGTTTCGTGGATTCATGCC |
| CEP-1 | TGTCCAGAAAATGATAGACGGAGT | GCATCGGAAATCTTTGGCGT |
| EGL-1 | ACACCCAAAACATTCACACCG | GGCAAAGGTGAGCATCAGCA |
| CED-13 | TCGAGGGCAGAAAAACGTGA | ACAACAGCGGGAGAAAGTGT |
| Groups | Median Survival (d) | p Value (Log-Rank) |
|---|---|---|
| CT | 10.5 **** | <0.0001 |
| UV | 4.5 | - |
| QL-UV | 6 * | 0.0770 |
| QH-UV | 6 * | 0.0169 |
| ML-UV | 6 * | 0.0149 |
| MH-UV | 7 *** | 0.0003 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.-m.; Liu, D.; Liu, Y.-l.; Liu, B.; Chen, X.-h. Quercetin and Its Mixture Increase the Stress Resistance of Caenorhabditis elegans to UV-B. Int. J. Environ. Res. Public Health 2020, 17, 1572. https://doi.org/10.3390/ijerph17051572
Li S-m, Liu D, Liu Y-l, Liu B, Chen X-h. Quercetin and Its Mixture Increase the Stress Resistance of Caenorhabditis elegans to UV-B. International Journal of Environmental Research and Public Health. 2020; 17(5):1572. https://doi.org/10.3390/ijerph17051572
Chicago/Turabian StyleLi, Shi-ming, Dan Liu, Yi-lin Liu, Bin Liu, and Xing-huang Chen. 2020. "Quercetin and Its Mixture Increase the Stress Resistance of Caenorhabditis elegans to UV-B" International Journal of Environmental Research and Public Health 17, no. 5: 1572. https://doi.org/10.3390/ijerph17051572
APA StyleLi, S.-m., Liu, D., Liu, Y.-l., Liu, B., & Chen, X.-h. (2020). Quercetin and Its Mixture Increase the Stress Resistance of Caenorhabditis elegans to UV-B. International Journal of Environmental Research and Public Health, 17(5), 1572. https://doi.org/10.3390/ijerph17051572