Population Genetic Structure Reveals Two Lineages of Amynthas triastriatus (Oligochaeta: Megascolecidae) in China, with Notes on a New Subspecies of Amynthas triastriatus


Abstract
1. Introduction
2. Materials and Methods
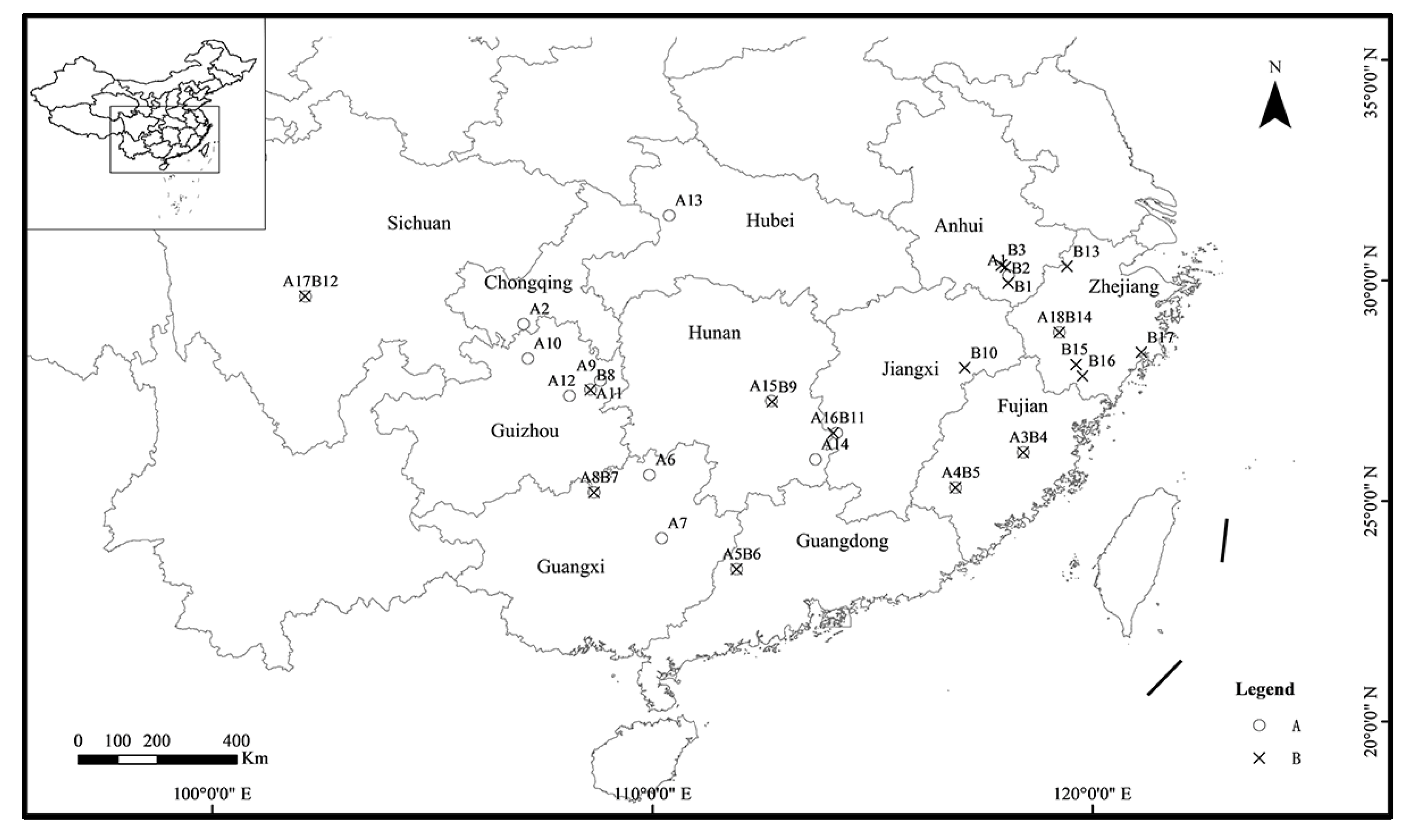
2.1. Earthworm Sampling
2.2. DNA Extraction, Amplification and Sequencing
2.3. Sequence Alignment, Population Genetic Structure and Genetic Diversity, Time of Divergence and Colonization History
3. Results
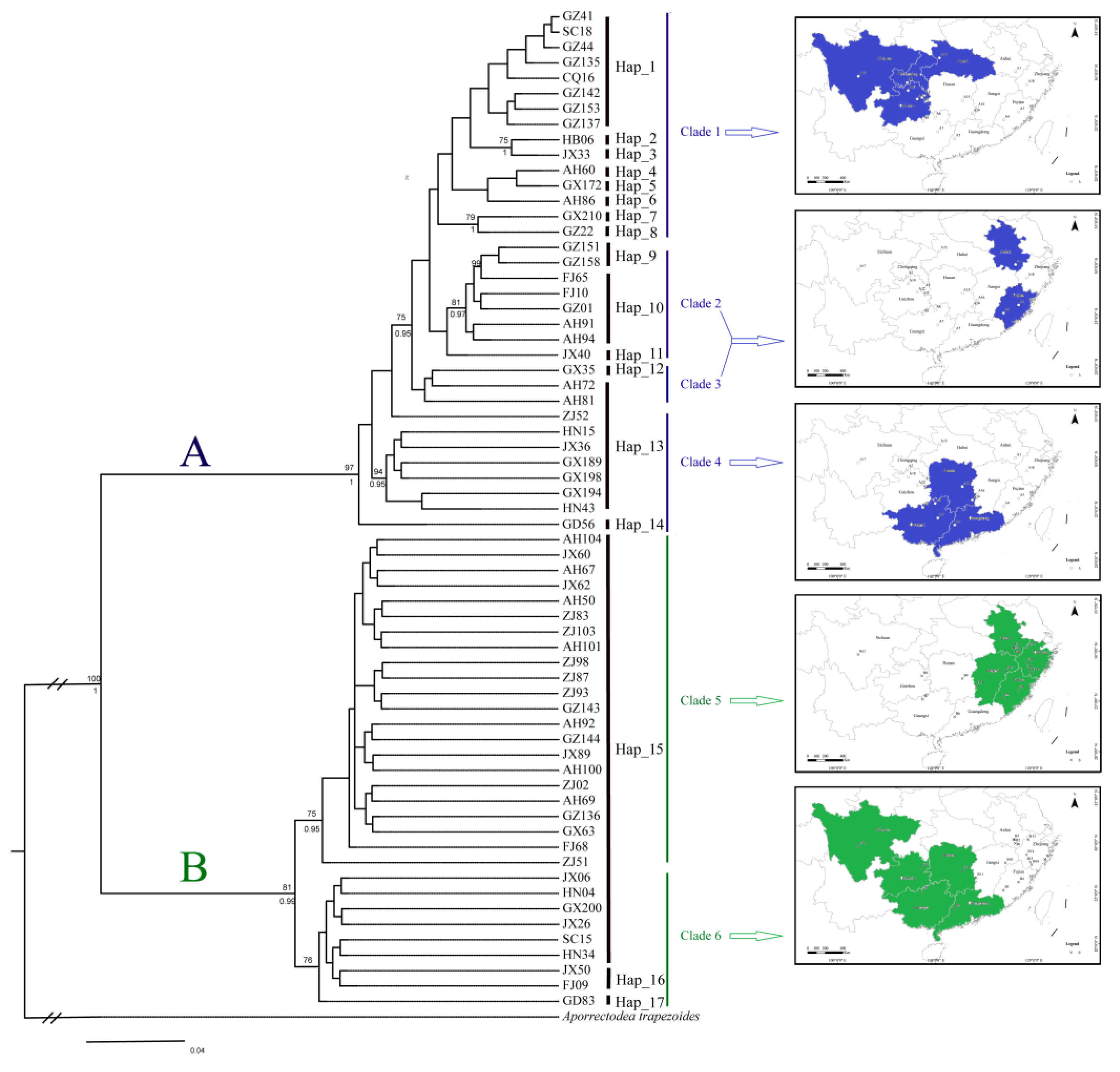
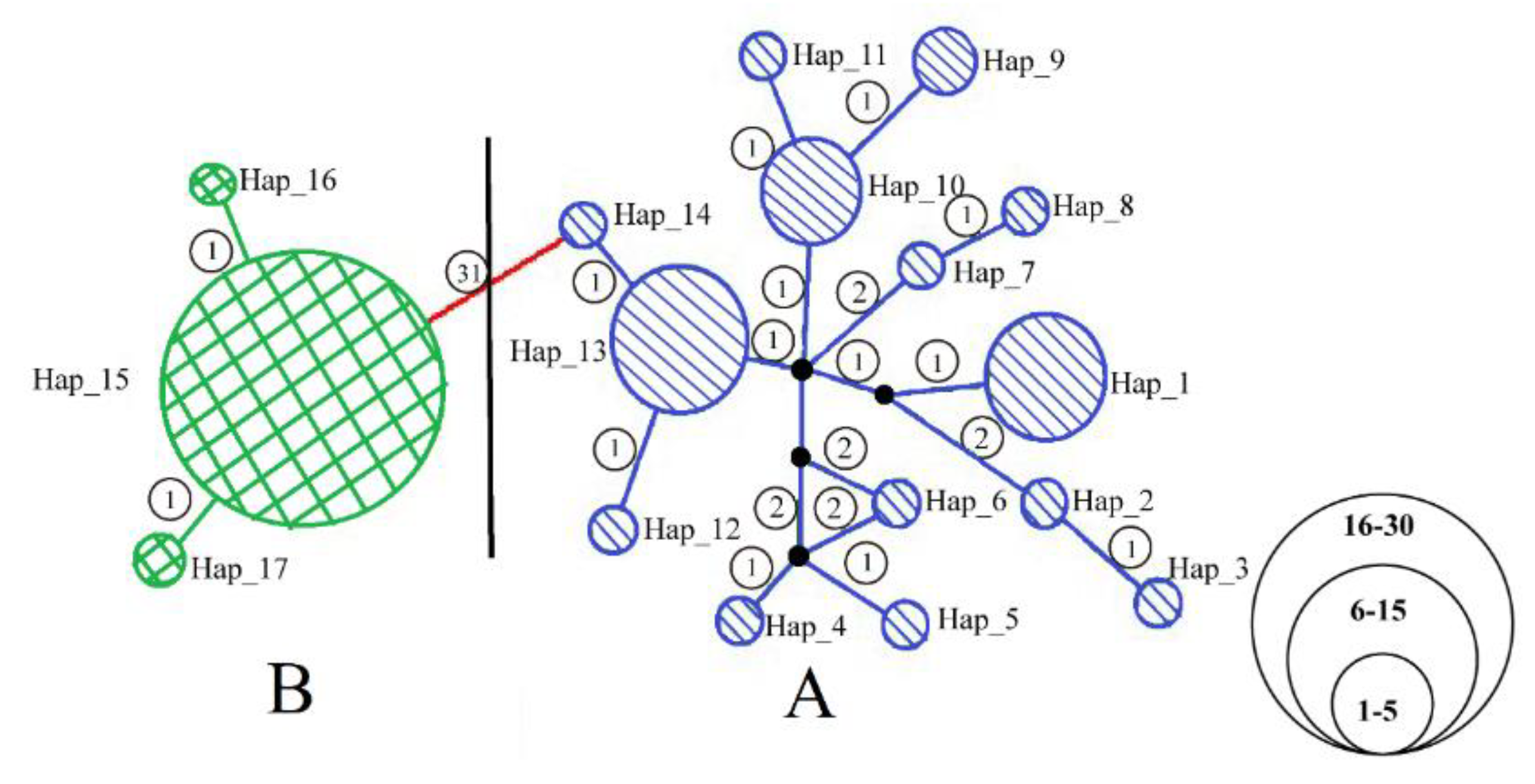
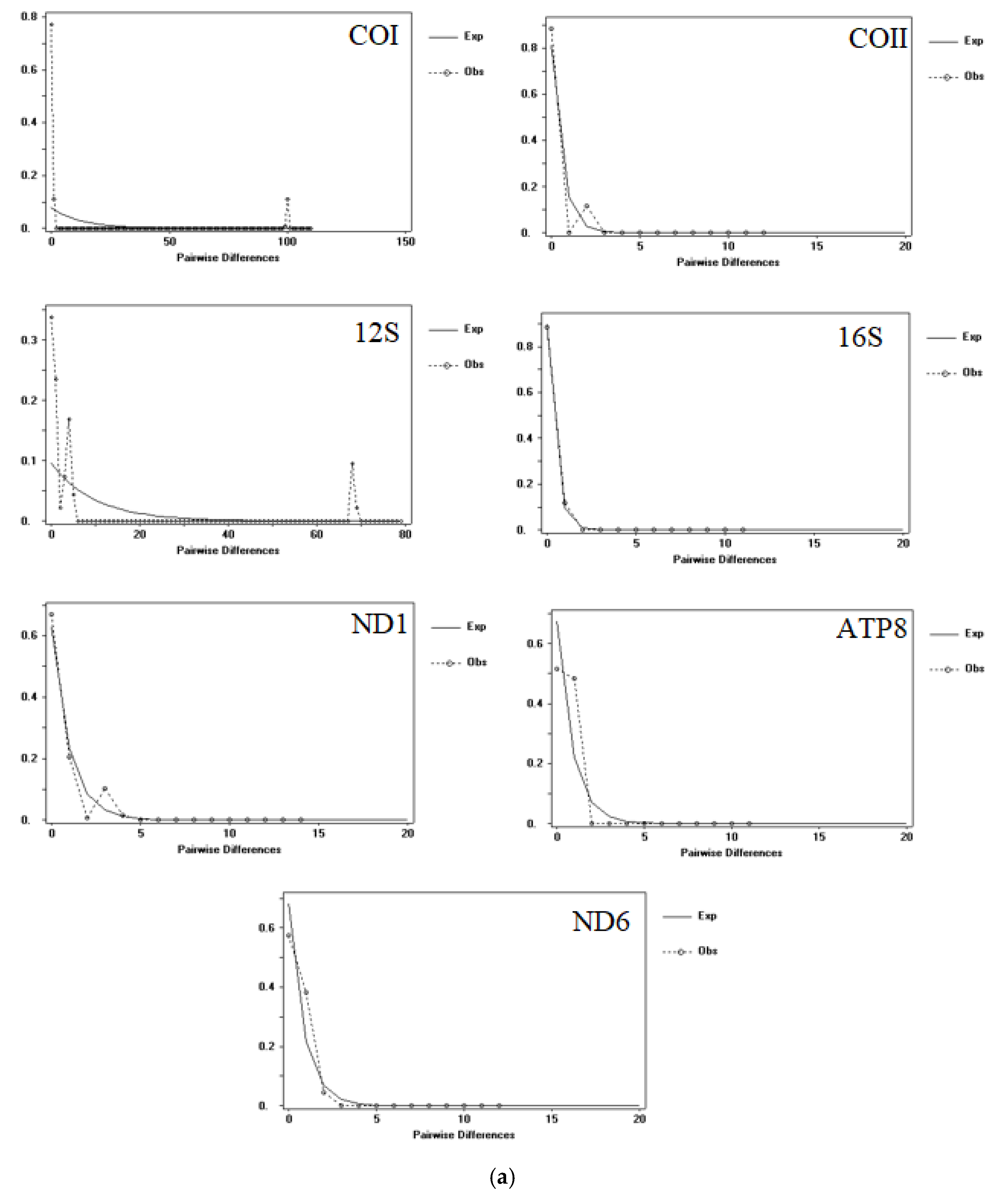
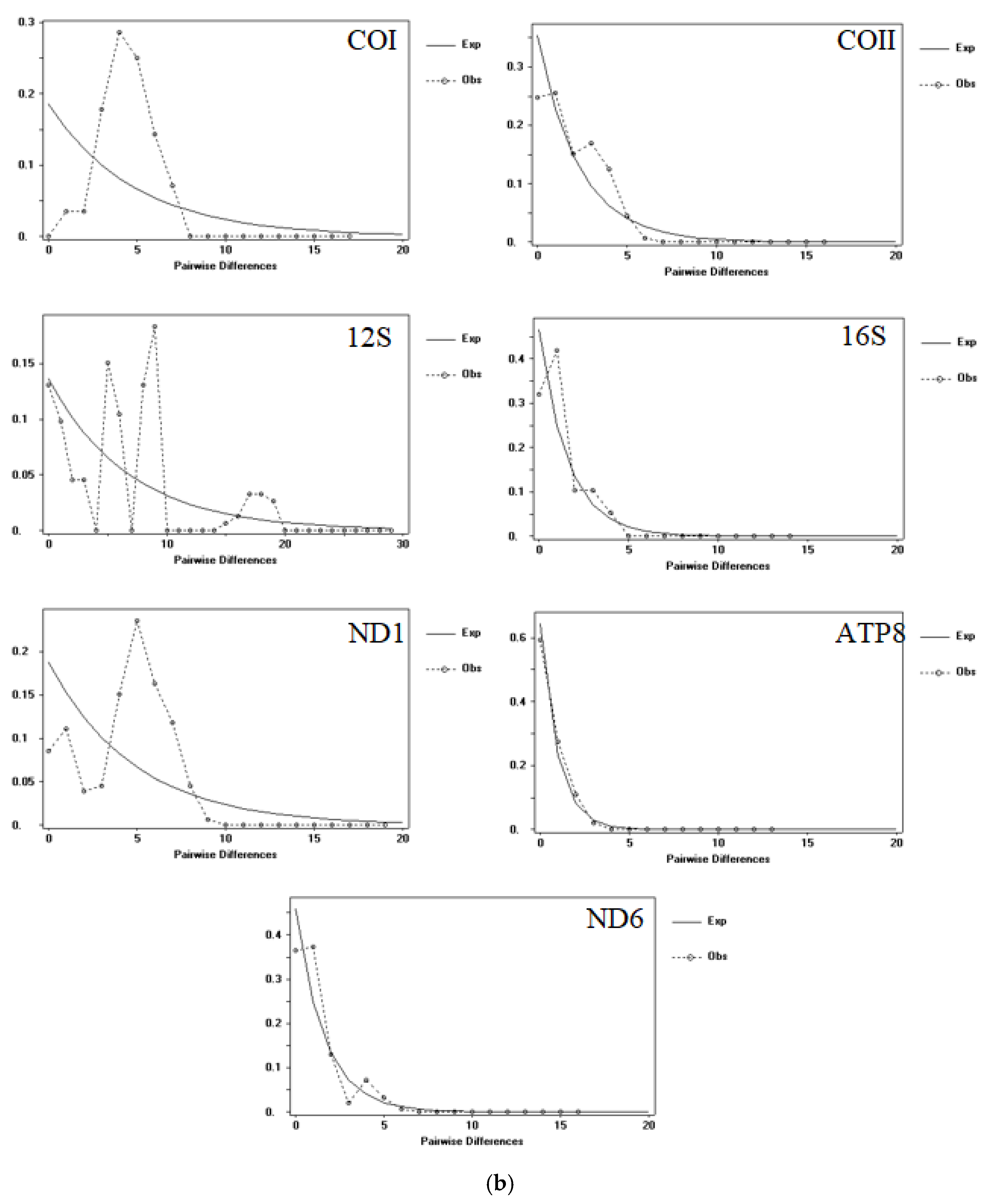
3.1. Population Genetic Structure and Genetic Diversity
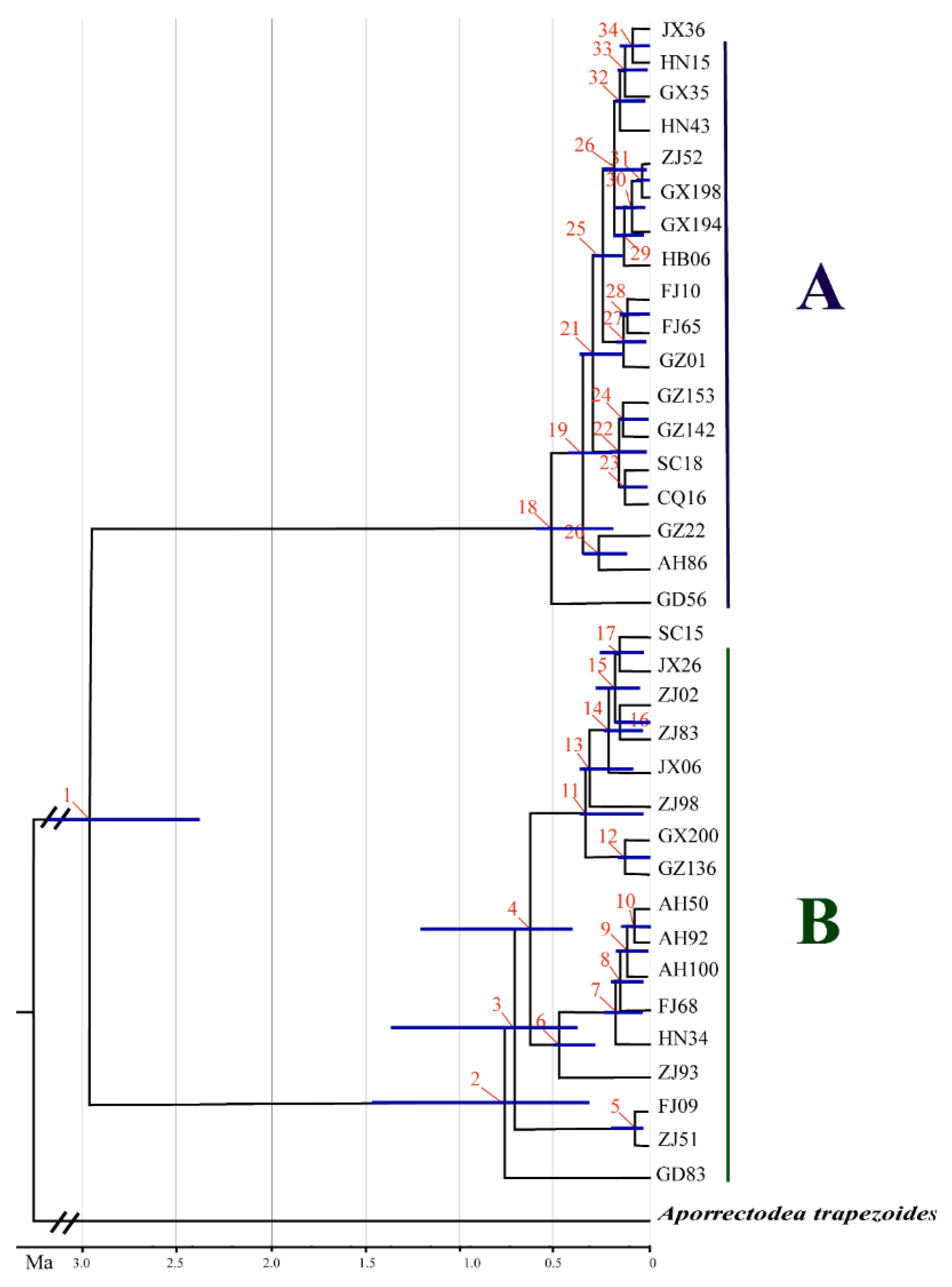
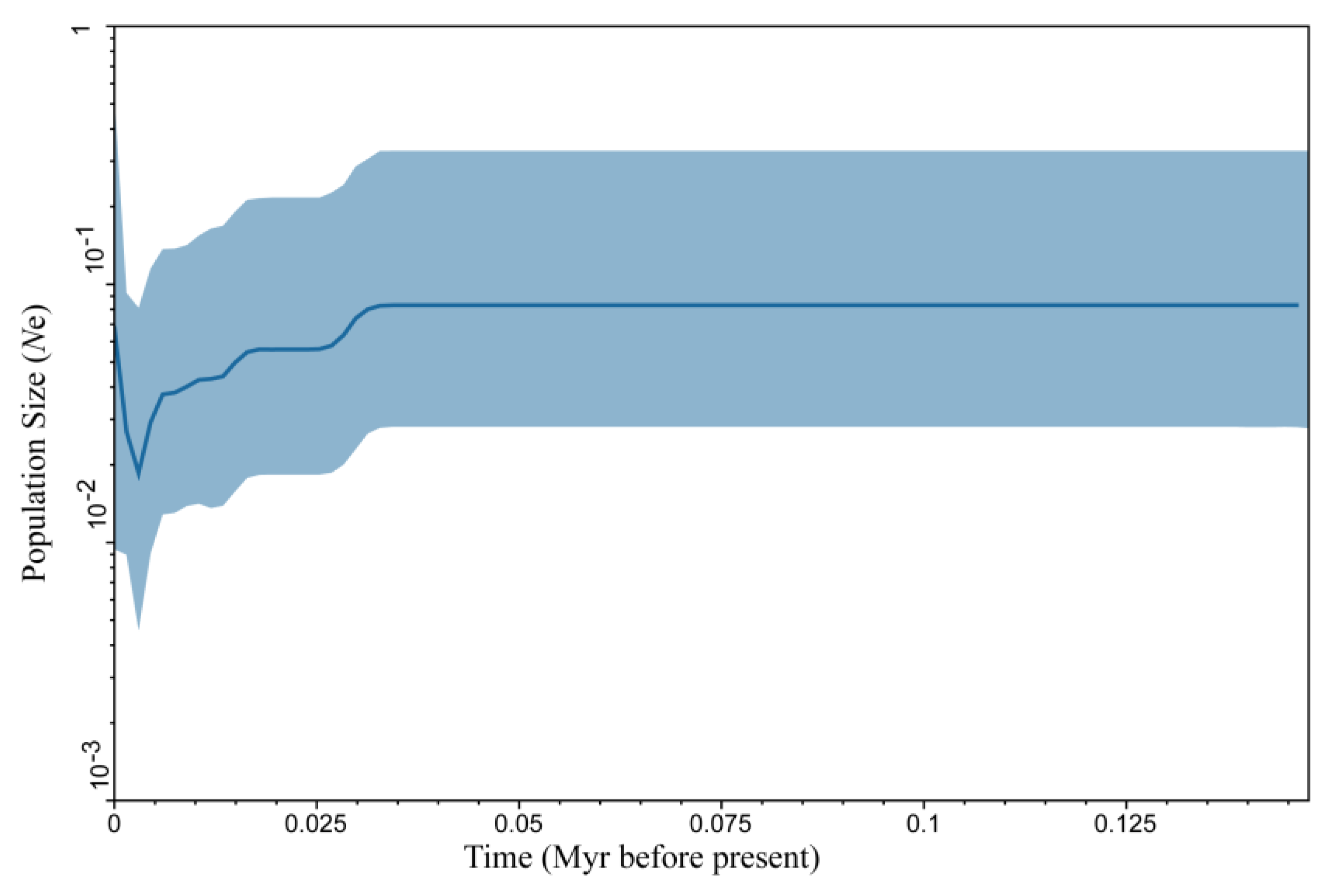
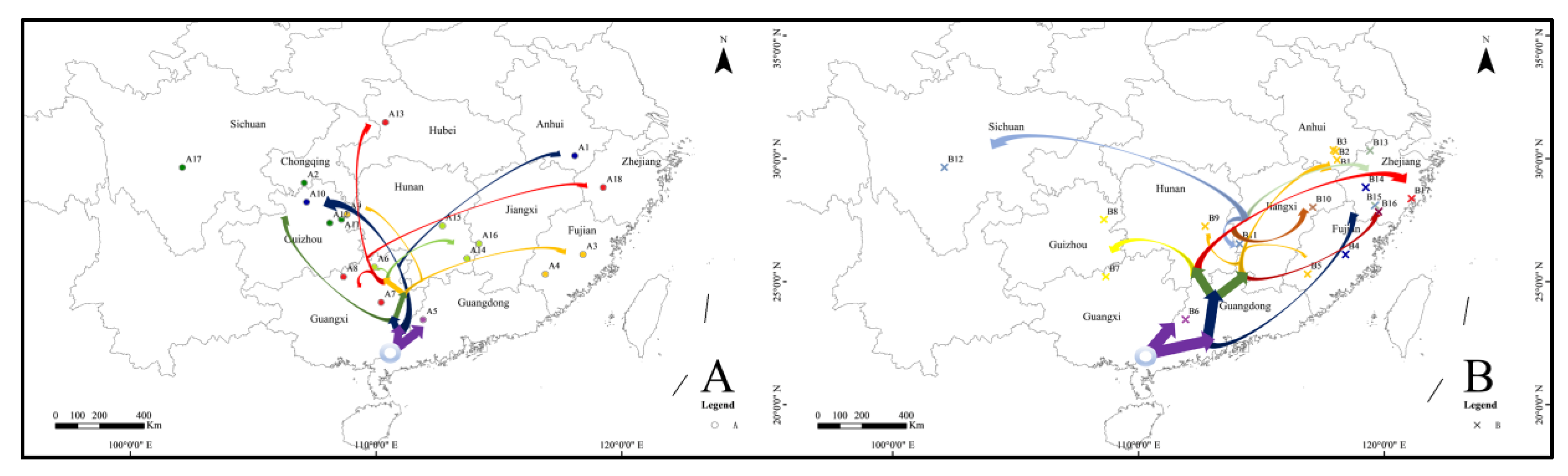
3.2. Time of Divergence and Colonization History
4. Discussion
4.1. Application of Molecular Techniques in Taxonomy and Differentiation of Earthworms
4.2. Parthenogenesis and the Demographic and Dispersal of A. triastriatus
4.3. Differentiation and Colonization of A. triastriatus
5. Conclusions
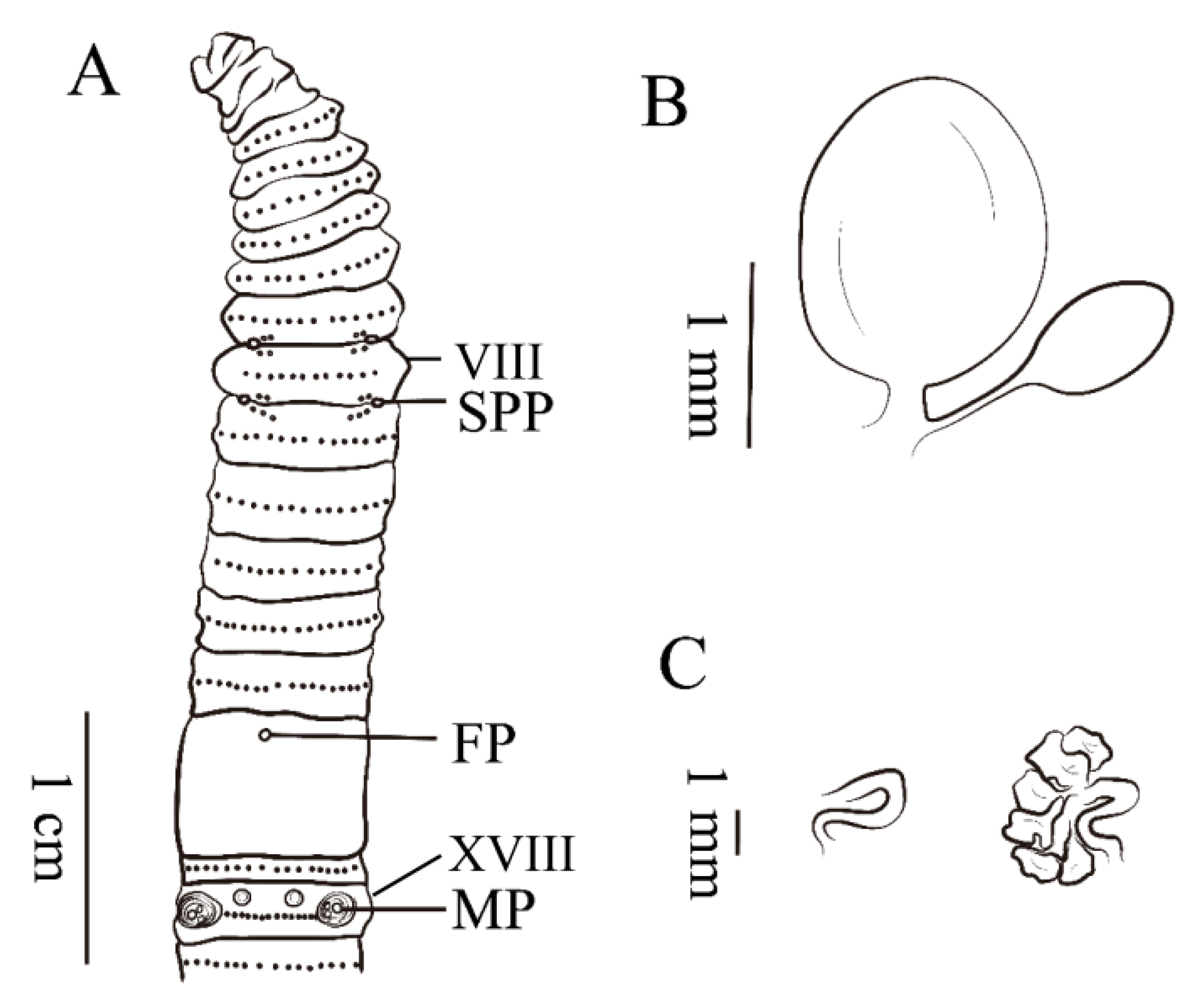
6. Description of a New Subspecies of Amynthas Triastriatus
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoffman, E.A.; Blouin, M.S. Evolutionary history of the northern leopard frog: Reconstruction of phylogeny, phylogeography, and historical changes in population demography from mitochondrial DNA. Evolution 2004, 58, 145–159. [Google Scholar] [CrossRef]
- Nyakaana, S.; Arctander, P. Population genetic structure of the African elephant in Uganda based on variation at mitochondrial and nuclear loci: Evidence for male-biased gene flow. Mol. Ecol. 1999, 8, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.L.; Chen, H.; Hu, L.L.; Wang, L.X.; Wang, S.H.; Wang, M.L.; Cheng, X.Z. Genetic diversity a population structure analysis of a accessions in the Chinese cowpea [Vigna unguiculata (L.) Walp.] germplasm collection. Crop J. 2017, 5, 363–372. [Google Scholar] [CrossRef]
- Winding, A.; Ronn, R.; Hendriksen, N.B. Bacteria and protozoa in soil microhabitats as affected by earthworms. Biol. Fert. Soils 1997, 24, 133–140. [Google Scholar] [CrossRef]
- Fernández, R.; Almodóvar, A.; Novo, M.; Gutiérrez, M.; Díaz Cosín, D.J. A vagrant clone in a peregrine species: Phylogeography, high clonal diversity and geographical distribution in the earthworm Aporrectodea trapezoides (Dugès, 1828). Soil Biol. Biochem. 2011, 43, 2085–2093. [Google Scholar] [CrossRef]
- Novo, M.; Almodovar, A.; Diaz-Cosin, D.J. High genetic divergence of hormogastrid earthworms (Annelida, Oligochaeta) in the central Iberian Peninsula: Evolutionary and demographic implications. Zool. Scr. 2009, 38, 537–552. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Berman, D.I.; Peltek, S.E. Phylogeography of the earthworm Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) in northeastern Eurasia. Dokl. Biol. Sci. 2015, 461, 85–88. [Google Scholar] [CrossRef]
- Fernández, R.; Almodóvar, A.; Novo, M.; Gutiérrez, M.; Díaz Cosín, D.J. Earthworms, good indicators for palaeogeographical studies? Testing the genetic structure and demographic history in the peregrine earthworm Aporrectodea trapezoides (Dugès, 1828) in southern Europe. Soil Biol. Biochem. 2013, 58, 127–135. [Google Scholar] [CrossRef]
- Thakuria, D.; Schmidt, O.; Finan, D.; Egan, D.; Doohan, F.M. Gut wall bacteria of earthworms: A natural selection process. ISME J. 2010, 4, 357–366. [Google Scholar] [CrossRef]
- Groffman, P.M.; Fahey, T.J.; Fisk, M.C.; Yavitt, J.B.; Sherman, R.E.; Bohlen, P.J.; Maerz, J.C. Earthworms increase soil microbial biomass carrying capacity and nitrogen retention in northern hardwood forests. Soil Biol. Biochem. 2015, 87, 51–58. [Google Scholar] [CrossRef]
- Braga, L.P.P.; Yoshiura, C.A.; Borges, C.D.; Horn, M.A.; Brown, G.G.; Drake, H.L.; Tsai, S.M. Disentangling the influence of earthworms in sugarcane rhizosphere. Sci. Rep. 2016, 6, 38923. [Google Scholar] [CrossRef] [PubMed]
- King, R.A.; Tibble, A.L.; Symondson, W.O.C. Opening a can of worms: Unprecedented sympatric cryptic diversity within British lumbricid earthworms. Mol. Ecol. 2008, 17, 4684–4698. [Google Scholar] [CrossRef] [PubMed]
- Perez-Losada, M.; Eiroa, J.; Mato, S.; Dominguez, J. Phylogenetic species delimitation of the earthworms Eisenia fetida (Savigny, 1826) and Eisenia andrei Bouche, 1972 (Oligochaeta, Lumbricidae) based on mitochondrial and nuclear DNA sequences. Pedobiologia 2005, 49, 317–324. [Google Scholar] [CrossRef]
- Domínguez, J.; Bohlen, P.J.; Parmelee, R.W. Earthworms Increase Nitrogen Leaching to Greater Soil Depths in Row Crop Agroecosystems. Ecosystems 2004, 7, 672–685. [Google Scholar] [CrossRef]
- Novo, M.; Almodovar, A.; Fernandez, R.; Trigo, D.; Cosin, D.J.D. Cryptic speciation of hormogastrid earthworms revealed by mitochondrial and nuclear data. Mol. Phylogenet. Evol. 2010, 56, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Minamiya, Y.; Yokoyama, J.; Fukuda, T. A phylogeographic study of the Japanese earthworm, Metaphire sieboldi (Horst, 1883) (Oligochaeta: Megascolecidae): Inferences from mitochondrial DNA sequences. Eur. J. Soil Biol. 2009, 45, 423–430. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Zhang, D.H.; Xu, Y.L.; Zhang, G.S.; Sun, Z.J. Effects of fragmentation on genetic variation in populations of the terrestrial earthworm Drawida japonica Michaelsen, 1892 (Oligochaeta, Moniligastridae) in Shandong and Liaodong peninsulas, China. J. Nat. Hist. 2012, 46, 1387–1405. [Google Scholar] [CrossRef]
- Aspe, N.M.; James, S.W. Molecular phylogeny and biogeographic distribution of pheretimoid earthworms (clitellata: Megascolecidae) of the Philippine archipelago. Eur. J. Soil Biol. 2018, 85, 89–97. [Google Scholar] [CrossRef]
- Dupont, L.; Gresille, Y.; Richard, B.; Decaens, T.; Mathieu, J. Dispersal constraints and fine-scale spatial genetic structure in two earthworm species. Biol. J. Linn. Soc. 2015, 114, 335–347. [Google Scholar] [CrossRef]
- Novo, M.; Almodovar, A.; Fernandez, R.; Giribet, G.; Cosin, D.J.D. Understanding the biogeography of a group of earthworms in the Mediterranean basin-The phylogenetic puzzle of Hormogastridae (Clitellata: Oligochaeta). Mol. Phylogenet. Evol. 2011, 61, 125–135. [Google Scholar] [CrossRef]
- James, S.W.; Porco, D.; Decaens, T.; Richard, B.; Rougerie, R.; Erseus, C. DNA Barcoding Reveals Cryptic Diversity in Lumbricus terrestris L., 1758 (Clitellata): Resurrection of L. herculeus (Savigny, 1826). PLoS ONE 2010, 5, e15629. [Google Scholar] [CrossRef] [PubMed]
- Buckley, T.R.; James, S.; Allwood, J.; Bartlam, S.; Howitt, R.; Prada, D. Phylogenetic analysis of New Zealand earthworms (Oligochaeta: Megascolecidae) reveals ancient clades and cryptic taxonomic diversity. Mol. Phylogenet. Evol. 2011, 58, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Richard, B.; Decaens, T.; Rougerie, R.; James, S.W.; Porco, D.; Hebert, P.D.N. Re-integrating earthworm juveniles into soil biodiversity studies: Species identification through DNA barcoding. Mol. Ecol. Resour. 2010, 10, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; James, S. A critique of earthworm molecular phylogenetics. Pedobiologia 2011, 54, S3–S9. [Google Scholar] [CrossRef]
- Csuzdi, C.; Chang, C.H.; Pavlicek, T.; Szederjesi, T.; Esopi, D.; Szlavecz, K. Molecular phylogeny and systematics of native North American lumbricid earthworms (Clitellata: Megadrili). PloS ONE 2017, 12, 8–e0181504. [Google Scholar] [CrossRef]
- James, S.W.; Davidson, S.K. Molecular phylogeny of earthworms (Annelida:Crassiclitellata) based on 28S, 18S and 16S gene sequences. Invertebr. Syst. 2012, 26, 213–229. [Google Scholar] [CrossRef]
- Schubert, M.; Csuzdi, C.; Chang, C.-H.; Pavlícek, T.; Szederjesi, T.; Esopi, D.; Szlávecz, K. Molecular phylogeny and systematics of native North American lumbricid earthworms (Clitellata: Megadrili). PLoS ONE 2017, 12, e0181504. [Google Scholar]
- Sun, J.; James, S.W.; Jiang, J.B.; Yao, B.; Zhang, L.L.; Liu, M.Q.; Qiu, J.P.; Hu, F. Phylogenetic evaluation of Amynthas earthworms from South China reveals the initial ancestral state of spermathecae. Mol. Phylogenet. Evol. 2017, 115, 106–114. [Google Scholar] [CrossRef]
- Schult, N.; Pittenger, K.; Davalos, S.; McHugh, D. Phylogeographic analysis of invasive Asian earthworms (Amynthas) in the northeast United States. Invertebr. Biol. 2016, 135, 314–327. [Google Scholar] [CrossRef]
- Zhang, L.L.; Jiang, J.B.; Dong, Y.; Qiu, J.P. Complete mitochondrial genome of four pheretimoid earthworms (Clitellata: Oligochaeta) and their phylogenetic reconstruction. Gene 2015, 574, 308–316. [Google Scholar] [CrossRef]
- Halanych, K.M.; Janosik, A.M. A review of molecular markers used for annelid phylogenetics. Integr. Comp. Biol. 2006, 46, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Nygren, A.; Sundberg, P. Phylogeny and evolution of reproductive modes in Autolytinae (Syllidae, Annelida). Mol. Phylogenet. Evol. 2003, 29, 235–249. [Google Scholar] [CrossRef]
- Perez-Losada, M.; Ricoy, M.; Marshall, J.C.; Dominguez, J. Phylogenetic assessment of the earthworm Aporrectodea caliginosa species complex (Oligochaeta: Lumbricidae) based on mitochondrial and nuclear DNA sequences. Mol. Phylogenet. Evol. 2009, 52, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.B. Taxonomy and Molecular Phylogeny of the Family Megascolecidae Earthworms from China. Ph.D. Thesis, Shanghai Jiao Tong University Shanghai, Shanghai, China, 2016. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Bely, A.E.; Wray, G.A. Molecular phylogeny of naidid worms (Annelida: Clitellata) based on cytochrome oxidase I. Mol. Phylogenet. Evol. 2004, 30, 50–63. [Google Scholar] [CrossRef]
- Hillis, D.M.; Mable, B.K.; Larson, A.; Davis, S.K.; Zimmer, E.A. Nucleic acids, IV: Sequencing and cloning. In Molecular Systematics; Hillis, D.M., Moritz, C., Mable, B.K., Eds.; Sinauer: Sutherland, MA, USA, 1996; pp. 321–381. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphisms. Genetics 1989, 123, 585–595. [Google Scholar] [PubMed]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Dong, X.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Yuan, Z.; Jiang, J.B.; Dong, Y.; Zhao, Q.; Gao, X.; Qiu, J.P. The dispersal and diversification of earthworms (Annelida: Oligochaeta) related to paleogeographical events in the Hengduan Mountains. Eur. J. Soil Biol. 2019, 94, 103–118. [Google Scholar] [CrossRef]
- Bouche, M. L’eolution spatiotemporelle des lombrixiens. Doc. Pedozool. Integrol. 1998, 3, 1–28. [Google Scholar]
- Jiang, J.B.; Qiu, J.P. Origin and evolution of earthworms belonging to the family Megascolecidae in China. Biodivers. Sci. 2018, 26, 1074–1082. [Google Scholar] [CrossRef]
- Dominguez, J.; Aira, M.; Breinholt, J.W.; Stojanovic, M.; James, S.W.; Perez-Losada, M. Underground evolution: New roots for the old tree of lumbricid earthworms. Mol. Phylogenet. Evol. 2015, 83, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Boore, J.L.; Brown, W.M. Complete DNA sequence of the mitochondrial genome of the annelid worm Lumbricus terrestris. Genet. Mol. Res. 1995, 141, 305–319. [Google Scholar]
- Sket, B. The nature of biodiversity in hypogean waters and how it is endangered. Biodivers. Conserv. 1999, 8, 1319–1338. [Google Scholar] [CrossRef]
- Proudlove, G.; Wood, P.J. The blind leading the blind: Cryptic subterranean species and DNA taxonomy. Trends Ecol. Evol. 2003, 18, 272–273. [Google Scholar] [CrossRef]
- Klarica, J.; Kloss-Brandstatter, A.; Traugott, M.; Juen, A. Comparing four mitochondrial genes in earthworms—Implications for identification, phylogenetics, and discovery of cryptic species. Soil Biol. Biochem. 2012, 45, 23–30. [Google Scholar] [CrossRef]
- Decaens, T.; Porco, D.; Rougerie, R.; Brown, G.G.; James, S.W. Potential of DNA barcoding for earthworm research in taxonomy and ecology. Appl. Soil Ecol. 2013, 65, 35–42. [Google Scholar] [CrossRef]
- Martinsson, S.; Rhodén, C.; Erséus, C. Barcoding gap, but no support for cryptic speciation in the earthworm Aporrectodea longa (Clitellata: Lumbricidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2017, 28, 147–155. [Google Scholar]
- Welch, D.M.; Meselson, M. Evidence for the evolution of bdelloid rotifers without sexual reproduction or genetic exchange. Science 2000, 288, 1211–1215. [Google Scholar] [CrossRef]
- Graham, B. The Masterpiece of Nature: The Evolution and Genetics of Sexuality; University of Califormia Press: Berkeley, CA, USA, 1982; p. 635. [Google Scholar]
- Muldal, S. The chromosomes of the earthworms: I. The evolution of polyploidy. Helv. Chim. Acta 1952, 6, 56–76. [Google Scholar] [CrossRef]
- Casellato, S. On polyploidy in oligochaetes with particular reference to Lumbricids. In On Earthworms; Omodeo, P., Ed.; Bonvicini Pagliai AM: Modena, Italy, 1987; pp. 77–87. [Google Scholar]
- Viktorov, A.G. Diversity of polyploid races in the family Lumbricidae. Soil Biol. Biochem. 1997, 29, 217–221. [Google Scholar] [CrossRef]
- Edwards, C.; Bohlen, P. Biology and Ecology of Earthworms, 3rd ed.; Chapman and Hall: London, UK, 1996; p. 426. [Google Scholar]
- Omodeo, P. Specie e razze poliploidi nei lombrichi. Conv. Genet. 1953, 23, 43–49. [Google Scholar]
- Gates, G. Contributions to North American earthworms. No. 3. IV. The trapezoides species group. Bull. Tall Timbers Res. Stn. 1972, 12, 146. [Google Scholar]
- Qiu, J.; Bouché, M. Contribution to the taxonomy of Hormogastridae (Annelida: Oligochaeta) with description of new species from Spain. Doc. Pedozool. Integrol. 1998, 4, 164–177. [Google Scholar]
- Cosín, D.J.D.; Novo, M.; Fernández, R. Reproduction of Earthworms: Sexual Selection and Parthenogenesis. In Biology of Earthworms; Springer: Berlin/Heidelberg, Germany, 2011; pp. 69–86. [Google Scholar]
- Terhivuo, J.; Saura, A. Dispersal and clonal diversity of North-European parthenogenetic earthworms. Biol. Invasions 2006, 8, 1205–1218. [Google Scholar] [CrossRef]
- Ghiselin, M.T. The Economy of Nature and the Evolution of Sex; University of California Press: Berkeley, CA, USA, 1974; p. 346. [Google Scholar]
- Glesener, R.R.; Tilman, D. Sexuality and the Components of Environmental Uncertainty: Clues from Geographic Parthenogenesis in Terrestrial Animals. Am. Nat. 1978, 112, 659–673. [Google Scholar] [CrossRef]
- Hamilton, W.D.; Henderson, P.A.; Moran, N.; Alexander, R.; Tinkle, D.; Press, C. Fluctuation of environment and coevolved antagonist polymorphism as factors in the maintenance of sex. In Natural Selection and Social Behaviour; Alexander, R.D., Tinkle, D.W., Eds.; Chiron: New York, NY, USA, 1981; pp. 363–381. [Google Scholar]
- Lynch, M. Destabilizing Hybridization, General-Purpose Genotypes and Geographic Parthenogenesis. Q. Rev. Biol. 1984, 59, 527–590. [Google Scholar] [CrossRef]
- Kearney, M.; Blacket, M.J.; Strasburg, J.L.; Moritz, C. Waves of parthenogenesis in the desert: Evidence for the parallel loss of sex in a grasshopper and a gecko from Australia. Mol. Ecol. 2006, 15, 1743–1748. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.C.; Delmotte, F.; Rispe, C.; Crease, T. Phylogenetic relationships between parthenogens and their sexual relatives: The possible routes to parthenogenesis in animals. Biol. J. Linn. Soc. 2003, 79, 151–163. [Google Scholar] [CrossRef]
- Minamiya, Y.; Hayakawa, H.; Ohga, K.; Shimano, S.; Ito, M.T.; Fukuda, T. Variability of sexual organ possession rates and phylogenetic analyses of a parthenogenetic Japanese earthworm, Amynthas vittatus (Oligochaeta: Megascolecidae). Genes Genet. Syst. 2011, 86, 27–35. [Google Scholar] [CrossRef][Green Version]
- Zhou, S.Z.; Li, J.J.; Zhang, S.Q. Quaternary glaciation of the Bailang River Valley, Qilian Shan. Quat. Int. 2002, 97–98, 103–110. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Wu, Y.Q.; Cui, Z.J. Kunlun-Yellow River Tectonic Motion and formation of modern physical geography pattern of China. J. Beijing Normal Univ. 2005, 41, 85–88. [Google Scholar]
- Wu, W. Natural Hybridization, PhyIogeography and Speciation Patterns of Altingiaceae. Ph.D. Thesis, Sun Yat-sen University Guangdong, Guangdong, China, 2009. [Google Scholar]
- Jörg, M.S.; Silvio, T.; Zhizhong, Z.; Xihao, W.; Susan, I.-O.; Rainer, W.; Heinrich, B.; Peter, W.K.; Christian, S. The limited influence of glaciations in Tibet on global climate over the past 170,000 yr. Earth Planet. Sci. Lett. 2002, 194, 287–297. [Google Scholar]
- Zhao, J.; Zhou, S.Z.; He, Y.Q.; Ye, Y.G.; Liu, S.Y. ESR dating of glacial tills and glaciations in the Urumqi River headwaters, Tianshan Mountains, China. Quat. Int. 2006, 144, 61–67. [Google Scholar] [CrossRef]
- Owen, L.A.; Finkel, R.C.; Barnard, P.L.; Haizhou, M.; Asahi, K.; Caffee, M.W.; Derbyshire, E. Climatic and topographic controls on the style and timing of Late Quaternary glaciation throughout Tibet and the Himalaya defined by 10Be cosmogenic radionuclide surface exposure dating. Quat. Sci. Rev. 2005, 24, 1391–1411. [Google Scholar]
- Zhao, J.; Liu, S.; He, Y.; Song, Y. Quaternary glacial chronology of the Ateaoyinake River Valley, Tianshan Mountains, China. Geomorphology 2009, 103, 276–284. [Google Scholar] [CrossRef]
- Shen, L.; Chen, X.Y.; Zhang, X.; Li, Y.Y.; Fu, C.X.; Qiu, Y.X. Genetic variation of ginkgo biloba l. (ginkgoaceae) based on cpdna pcr-rflps: Inference of glacial refugia. Heredity 2004, 94, 396–401. [Google Scholar] [CrossRef]
- Gong, W.; Chen, C.; Dobeš, C.; Fu, C.-X.; Koch, M. Phylogeography of a living fossil: Pleistocene glaciations forced Ginkgo biloba L. (Ginkgoaceae) into two refuge areas in China with limited subsequent postglacial expansion. Mol. Phylogenet. Evol. 2008, 48, 1094–1105. [Google Scholar] [CrossRef]
- Li, J.J.; Fang, X.M.; Pan, B.T.; Zhao, Z.J.; Song, Y.G. Late cenozoic intensive uplift of qinghai-xizang plateau and its impacts on environments in surrounding area. Quat. Sci. 2001, 21, 381–391. [Google Scholar]
- Thompson, L.G.; Yao, T.; Davis, M.E.; Henderson, K.A.; MosleyThompson, E.; Lin, P.N.; Beer, J.; Synal, H.A.; ColeDai, J.; Bolzan, J.F. Tropical climate instability: The last glacial cycle from a Qinghai-Tibetan ice core. Science 1997, 276, 1821–1825. [Google Scholar] [CrossRef]
- Yi, C.L.; Zhu, Z.Y.; Wei, L.; Cui, Z.J.; Zheng, B.X.; Shi, Y.F. Advances in numerical dating of Quaternary glaciations in China. Z. Geomorphol. 2007, 51, 153–175. [Google Scholar] [CrossRef]
- Yang, J.Q.; Zhang, W.; Cui, Z.J.; Yi, C.L.; Chen, Y.X.; Xu, X.G. Climate change since 11.5 ka on the Diancang Massif on the southeastern margin of the Tibetan Plateau. Quat. Res. 2010, 73, 304–312. [Google Scholar] [CrossRef]
- Cui, Z.J.; Wu, Y.Q.; Liu, G.N.; Ge, A.K.; Pang, Q.Q.; Xu, Q.H. On Kunlun-Yellow River tectonic movement. Sci. China 1998, 41, 592–600. [Google Scholar] [CrossRef]
- Shi, Y.F. Evolution of the cryosphere in the Tibetan Plateau, China, and its relationship with the global change in the mid-Quaternary. J. Glaciol. Geocryol. 1998, 20, 197–208. [Google Scholar]
- Chen, C.D. Biodiversity in the Wuyi Mountains and its importance in China. Chin. Biodivers. 1999, 7, 320–326. [Google Scholar]
- Novo, M.; Cunha, L.; Maceda-Veiga, A.; Talavera, J.A.; Hodson, M.E.; Spurgeon, D.; Bruford, M.W.; Morgan, A.J.; Kille, P. Multiple introductions and environmental factors affecting the establishment of invasive species on a volcanic island. Soil Biol. Biochem. 2015, 85, 89–100. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Tans. R. Soc. Lond. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenet. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef]
- Ren, G.P.; Mateo, R.G.; Liu, J.Q.; Suchan, T.; Alvarez, N.; Guisan, A.; Conti, E.; Salamin, N. Genetic consequences of Quaternary climatic oscillations in the Himalayas: Primula tibetica as a case study based on restriction site-associated DNA sequencing. New Phytol. 2017, 213, 1500–1512. [Google Scholar] [CrossRef]
- Wright, H.E. Late Pleistocene climate of Europe: A review. GSA Bull. 1961, 72, 933–984. [Google Scholar] [CrossRef]
- Dawson, A.G. Ice Age Earth; Routledge: London, UK, 1992; p. 293. [Google Scholar]
- Flenley, J.R. Quaternary Vegetation. (Book Reviews: The Equatorial Rain Forest. A Geological History). Science 1979, 206, 550. [Google Scholar]
- Verstappen, H. Quaternary Climatic Changes and Natural Environment in SE Asia. GeoJournal 1980, 4, 45–54. [Google Scholar] [CrossRef]
- Sun, J. Taxonomy and Molecular Phylogeny of Amynthas Earthworms from China. Ph.D. Thesis, Shanghai Jiao Tong University Shanghai, Shanghai, China, 2013. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Label | Location Code | Collection Location | GPS Coordinates | Collection Date | Elevation (m) | Gene Code | Number of Individuals |
|---|---|---|---|---|---|---|---|---|
| A | C-AH201608-01 | 1 | Anhui | 30.12324° N, 118.09639° E | 05/2016 | 93 | AH86 AH60 AH72 AH81 AH91 AH94 | 19 |
| C-CQ201304-02 | 2 | Chongqing | 29.01467° N, 107.08238° E | 06/2013 | 1162 | CQ16 | 2 | |
| C-FJ201106-01 | 3 | Fujian | 26.10627° N, 118.42997° E | 07/2011 | 342 | FJ10 | 1 | |
| C-FJ201111-01 | 4 | Fujian | 25.31278° N, 116.89306° E | 07/2011 | 1207 | FJ65 | 18 | |
| C-GD201109-03 | 5 | Guangdong | 23.46417° N, 111.91444° E | 10/2011 | 184 | GD56 | 4 | |
| C-GX201210-05 | 6 | Guangxi | 25.59723° N, 109.93519° E | 06/2012 | 1536 | GX35 | 13 | |
| C-GX201318-05 | 7 | Guangxi | 24.15622° N, 110.21279° E | 05/2013 | 1198 | GX194 GX172 GX189 | 20 | |
| C-GX201319-04 | 8 | Guangxi | 25.20595° N, 108.67555° E | 05/2013 | 1226 | GX198 GX210 | 2 | |
| C-GZ201101-01 | 9 | Guizhou | 27.73722° N, 108.81611° E | 10/2011 | 370 | GZ01 GZ41 GZ44 | 27 | |
| C-GZ201103-02 | 10 | Guizhou | 28.23472° N, 107.17444° E | 10/2011 | 1534 | GZ22 | 4 | |
| C-GZ201610-01 | 11 | Guizhou | 27.53520° N, 108.59300° E | 10/2016 | 604 | GZ142 GZ135 GZ137 | 14 | |
| C-GZ201611-05 | 12 | Guizhou | 27.39100° N, 108.11610° E | 10/2016 | 890 | GZ153 GZ151 GZ158 | 3 | |
| C-HB201101-06 | 13 | Hubei | 31.47611° N, 110.38055° E | 09/2011 | 1399 | HB06 | 1 | |
| C-HU201304-01 | 14 | Hunan | 25.94700° N, 113.70489° E | 06/2013 | 982 | HN15 | 4 | |
| C-HU201311-04 | 15 | Hunan | 27.27608° N, 112.70904° E | 06/2013 | 500 | HN43 | 1 | |
| C-JX201313-02 | 16 | Jiangxi | 26.54490° N, 114.18954° E | 06/2013 | 571 | JX36 JX33 JX40 | 10 | |
| C-SC201006-02 | 17 | Sichuan | 29.64400° N, 102.12433° E | 08/2010 | 1625 | SC18 | 2 | |
| C-ZJ201511-02 | 18 | Zhejiang | 28.83361° N, 119.24639° E | 07/2015 | 160 | ZJ52 | 1 | |
| B | C-AH201601-01 | 1 | Anhui | 29.95601° N, 118.09112° E | 05/2016 | 325 | AH50 AH67 AH69 | 10 |
| C-AH201609-02 | 2 | Anhui | 30.31267° N, 118.02561° E | 05/2016 | 120 | AH92 | 2 | |
| C-AH201610-01 | 3 | Anhui | 30.35924° N, 117.93609° E | 05/2016 | 146 | AH100 AH101 AH104 | 10 | |
| C-FJ201004-01 | 4 | Fujian | 26.11244° N, 118.42387° E | 08/2010 | 952 | FJ09 | 4 | |
| C-FJ201111-04 | 5 | Fujian | 25.31278° N, 116.89306° E | 07/2011 | 1207 | FJ68 | 3 | |
| C-GD201115-04 | 6 | Guangdong | 23.46417° N, 111.91444° E | 08/2011 | 410 | GD83 | 1 | |
| C-GX201319-06 | 7 | Guangxi | 25.20595° N, 108.67555° E | 05/2013 | 1226 | GX200 GX63 | 12 | |
| C-GZ201609-02 | 8 | Guizhou | 27.52480° N, 108.59300° E | 10/2016 | 731 | GZ136 GZ143 GZ144 | 4 | |
| C-HU201309-02 | 9 | Hunan | 27.25680° N, 112.72433° E | 06/2013 | 172 | HN34 HN04 | 2 | |
| C-JX201302-04 | 10 | Jiangxi | 28.03037° N, 117.09680° E | 06/2013 | 185 | JX06 | 2 | |
| C-JX201311-01 | 11 | Jiangxi | 26.55213° N, 114.10773° E | 06/2013 | 1300 | JX26 JX50 JX60 JX62 JX89 | 19 | |
| C-SC201004-06 | 12 | Sichuan | 29.65556° N, 102.11667° E | 08/2010 | 1558 | SC15 | 4 | |
| C-ZJ201101-02 | 13 | Zhejiang | 30.32886° N, 119.42599° E | 11/2011 | 1052 | ZJ02 | 1 | |
| C-ZJ201511-01 | 14 | Zhejiang | 28.83361° N, 119.24639° E | 07/2015 | 160 | ZJ51 | 1 | |
| C-ZJ201515-03 | 15 | Zhejiang | 28.10444° N, 119.63472° E | 07/2015 | 290 | ZJ83 ZJ87 | 4 | |
| C-ZJ201517-02 | 16 | Zhejiang | 27.84556° N, 119.77306° E | 07/2015 | 710 | ZJ93 | 1 | |
| C-ZJ201518-01 | 17 | Zhejiang | 28.37639° N, 121.10972° E | 07/2015 | 120 | ZJ98 ZJ103 | 6 |
| Gene | Primer | Sequence | Source |
|---|---|---|---|
| CO1 | LCO1490 | 5′-GGTCAACAAATCATAAAGATATTGG-3′ | [36] |
| HCO2198 | 5′-TAAACTTCAGGGTGACCAAAAAATCA-3 | [36] | |
| CO1-E | 5′-TATACTTCTGGGTGTCCGAAGAATCA-3′ | [37] | |
| CO2 | rRNA-Asn-CO2-tRNA-Asp: LumbF1 | 5′-GGCACCTATTTGTTAATTAGG-3′ | [33] |
| rRNA-Asn-CO2-tRNA-Asp: LumbR2 | 5′-GTGAGGCATAGAAATACACC-3′ | [33] | |
| ATP8 | ATP8-F | 5′-GYTTAGTTRCCAMCYGGTGTATTTC-3′ | This study |
| ATP8-R | 5′-CTTYYTACTTGGAAGGTARRTGTAC-3′ | This study | |
| ND6 | ND6-F | 5′-TGTATGGYGCACACRGGYHTTTGAA-3′ | This study |
| ND6-R | 5′-RATDGCTGGRGTNGGYTTAAACATA-3′ | This study | |
| 12S rRNA | 12S-tRNA-Val-16S-LumbF1 | 5′-CTTAAAGATTTTGGCGGTGTC-3′ | [33] |
| 12S-tRNA-Val-16S-LumbR1 | 5′-CCTTTGCACGGTTAGGATAC-3′ | [33] | |
| 16S rRNA | 16Sar | 5′-CGCCTGTTTATCAAAAACAT-3′ | [38] |
| 16Sbr | 5′-CCGGTCTGAACTCAGATCACGT-3′ | [38] | |
| ND1 | rRNA-Leu-ND1-LumbF2 | 5′-GAATAGTGCCACAGGTTTAAAC-3′ | [33] |
| rRNA-Leu-ND1-LumbR1b | 5′-TTAACGTCATCAGAGTTATC-3′ | [33] |
| Taxon | Number of Individuals | Number Haplotypes | Polymorphic Sites | Haplotype Diversity | Nucleotide Diversity | Tajima’s D | Fu’s FS | Mean Number of Pairwise Distance | |
|---|---|---|---|---|---|---|---|---|---|
| COI 7 | Gene | ||||||||
| A (within lineage) | 34 | 14 | 16 | 0.866 | 0.0052 | −1.028 | −4.637 | 0.5% | 0.6% |
| B (within lineage) | 31 | 3 | 2 | 0.127 | 0.0002 | −1.506 | −2.397 | 0.1% | 0.3% |
| Total (with lineage) | 65 | 17 | 49 | 0.769 | 0.0309 | 2.303 * | 7.525 | 6.3% | 6.7% |
| Node | Estimated Age in Millions of Years Ago (Ma) | ||
|---|---|---|---|
| Age | Lower 95% Confidence Limit | Upper 95% Confidence Limit | |
| 1 | 2.97 | 2.17 | 3.15 |
| 2 | 0.72 | 0.29 | 1.47 |
| 3 | 0.69 | 0.27 | 1.42 |
| 4 | 0.60 | 0.26 | 1.35 |
| 5 | 0.08 | 0.01 | 0.13 |
| 6 | 0.42 | 0.26 | 0.41 |
| 7 | 0.09 | 0.04 | 0.14 |
| 8 | 0.07 | 0.03 | 0.11 |
| 9 | 0.03 | 0.01 | 0.06 |
| 10 | 0.02 | 0.00 | 0.04 |
| 11 | 0.26 | 0.03 | 0.31 |
| 12 | 0.02 | 0.00 | 0.04 |
| 13 | 0.15 | 0.02 | 0.19 |
| 14 | 0.05 | 0.01 | 0.05 |
| 15 | 0.03 | 0.01 | 0.04 |
| 16 | 0.02 | 0.00 | 0.03 |
| 17 | 0.02 | 0.01 | 0.04 |
| 18 | 0.48 | 0.22 | 0.53 |
| 19 | 0.33 | 0.19 | 0.39 |
| 20 | 0.24 | 0.17 | 0.28 |
| 21 | 0.27 | 0.15 | 0.35 |
| 22 | 0.04 | 0.01 | 0.06 |
| 23 | 0.02 | 0.00 | 0.04 |
| 24 | 0.01 | 0.00 | 0.03 |
| 25 | 0.23 | 0.14 | 0.25 |
| 26 | 0.09 | 0.04 | 0.11 |
| 27 | 0.04 | 0.01 | 0.06 |
| 28 | 0.02 | 0.00 | 0.04 |
| 29 | 0.06 | 0.03 | 0.09 |
| 30 | 0.04 | 0.01 | 0.07 |
| 31 | 0.02 | 0.00 | 0.03 |
| 32 | 0.05 | 0.02 | 0.07 |
| 33 | 0.03 | 0.01 | 0.05 |
| 34 | 0.01 | 0.00 | 0.03 |
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation | ΦST |
|---|---|---|---|---|---|
| Between lineages | 1 | 629.617 | 19.37529 Va | 93.89 | 0.9396 |
| Within lineages | 63 | 79.445 | 1.26103 Vb | 6.11 | (p < 0.0001) |
| Total | 64 | 709.062 | 20.63632 | ||
| FST | 0.9389 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Jiang, J.; Yuan, Z.; Zhao, Q.; Qiu, J. Population Genetic Structure Reveals Two Lineages of Amynthas triastriatus (Oligochaeta: Megascolecidae) in China, with Notes on a New Subspecies of Amynthas triastriatus. Int. J. Environ. Res. Public Health 2020, 17, 1538. https://doi.org/10.3390/ijerph17051538
Dong Y, Jiang J, Yuan Z, Zhao Q, Qiu J. Population Genetic Structure Reveals Two Lineages of Amynthas triastriatus (Oligochaeta: Megascolecidae) in China, with Notes on a New Subspecies of Amynthas triastriatus. International Journal of Environmental Research and Public Health. 2020; 17(5):1538. https://doi.org/10.3390/ijerph17051538
Chicago/Turabian StyleDong, Yan, Jibao Jiang, Zhu Yuan, Qi Zhao, and Jiangping Qiu. 2020. "Population Genetic Structure Reveals Two Lineages of Amynthas triastriatus (Oligochaeta: Megascolecidae) in China, with Notes on a New Subspecies of Amynthas triastriatus" International Journal of Environmental Research and Public Health 17, no. 5: 1538. https://doi.org/10.3390/ijerph17051538
APA StyleDong, Y., Jiang, J., Yuan, Z., Zhao, Q., & Qiu, J. (2020). Population Genetic Structure Reveals Two Lineages of Amynthas triastriatus (Oligochaeta: Megascolecidae) in China, with Notes on a New Subspecies of Amynthas triastriatus. International Journal of Environmental Research and Public Health, 17(5), 1538. https://doi.org/10.3390/ijerph17051538