Effects of Cadmium on Bioaccumulation, Bioabsorption, and Photosynthesis in Sarcodia suiae

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
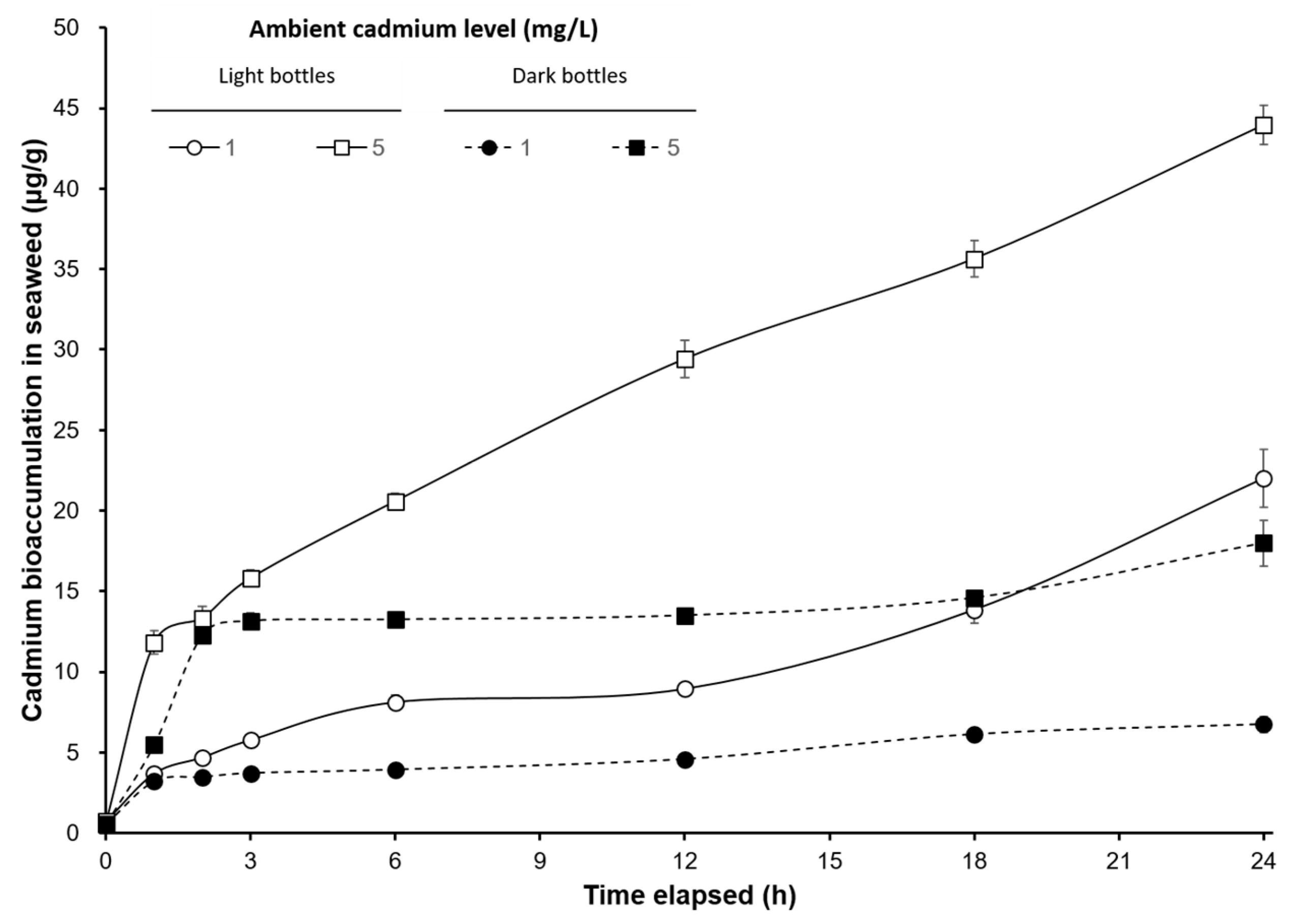
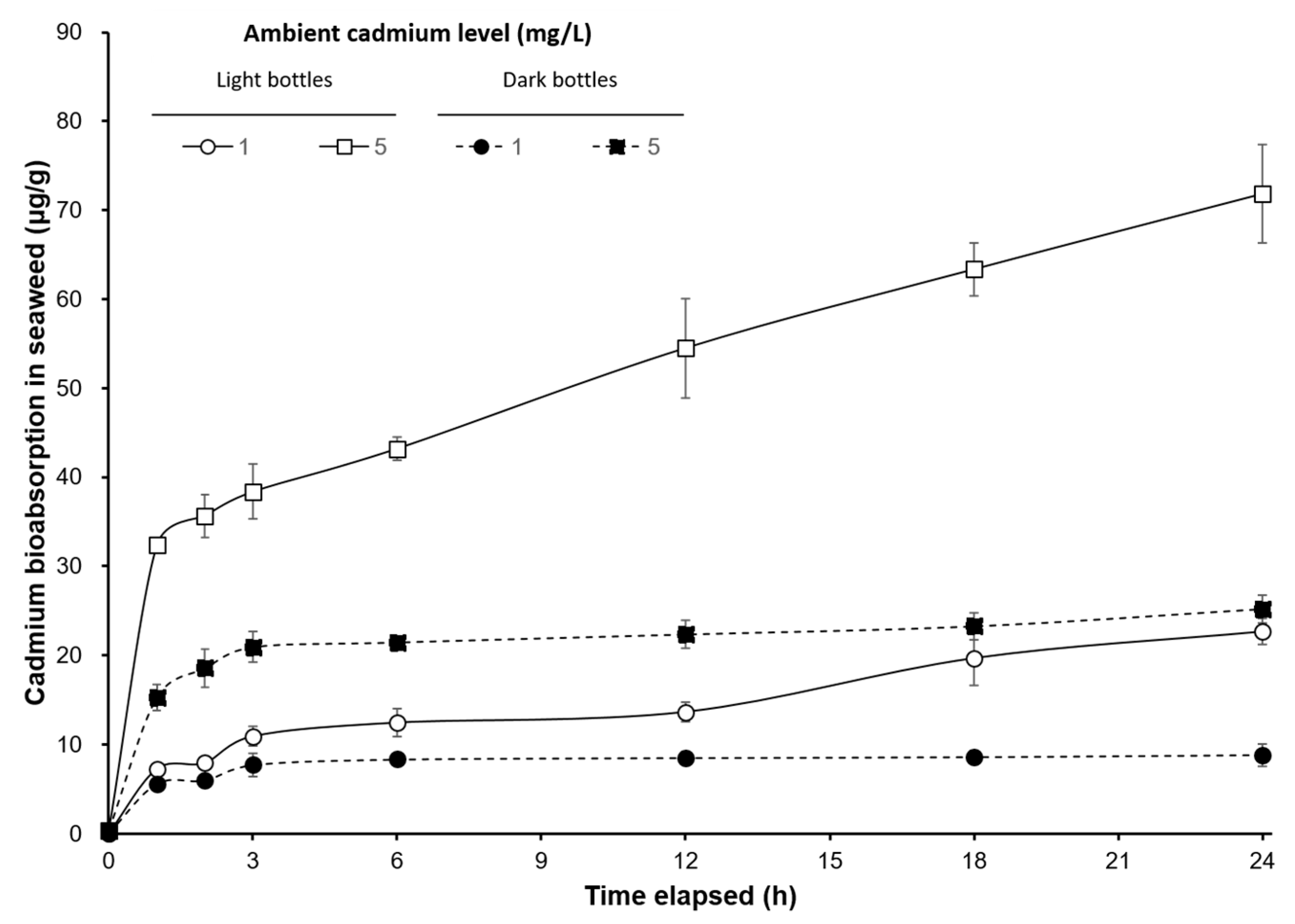
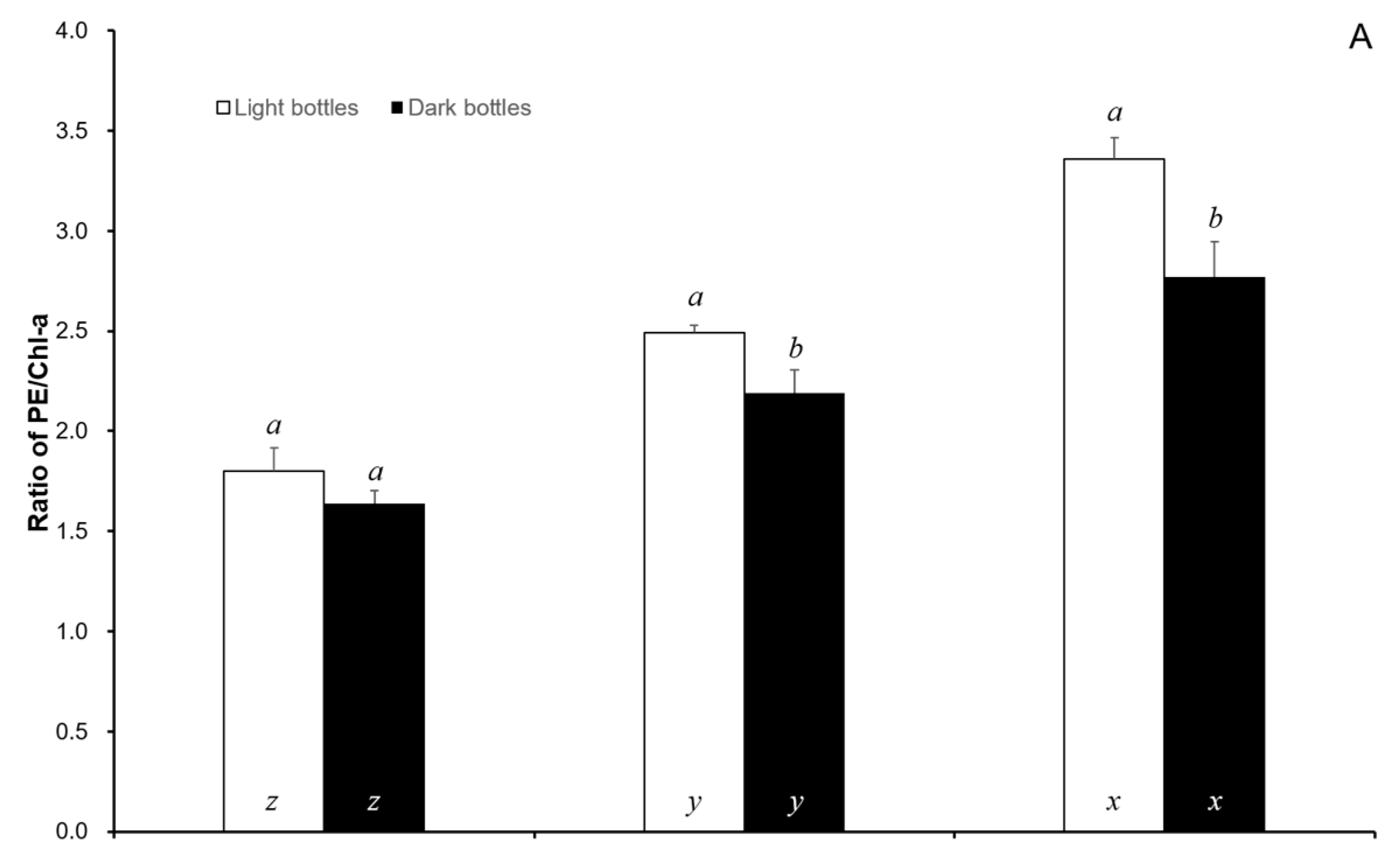
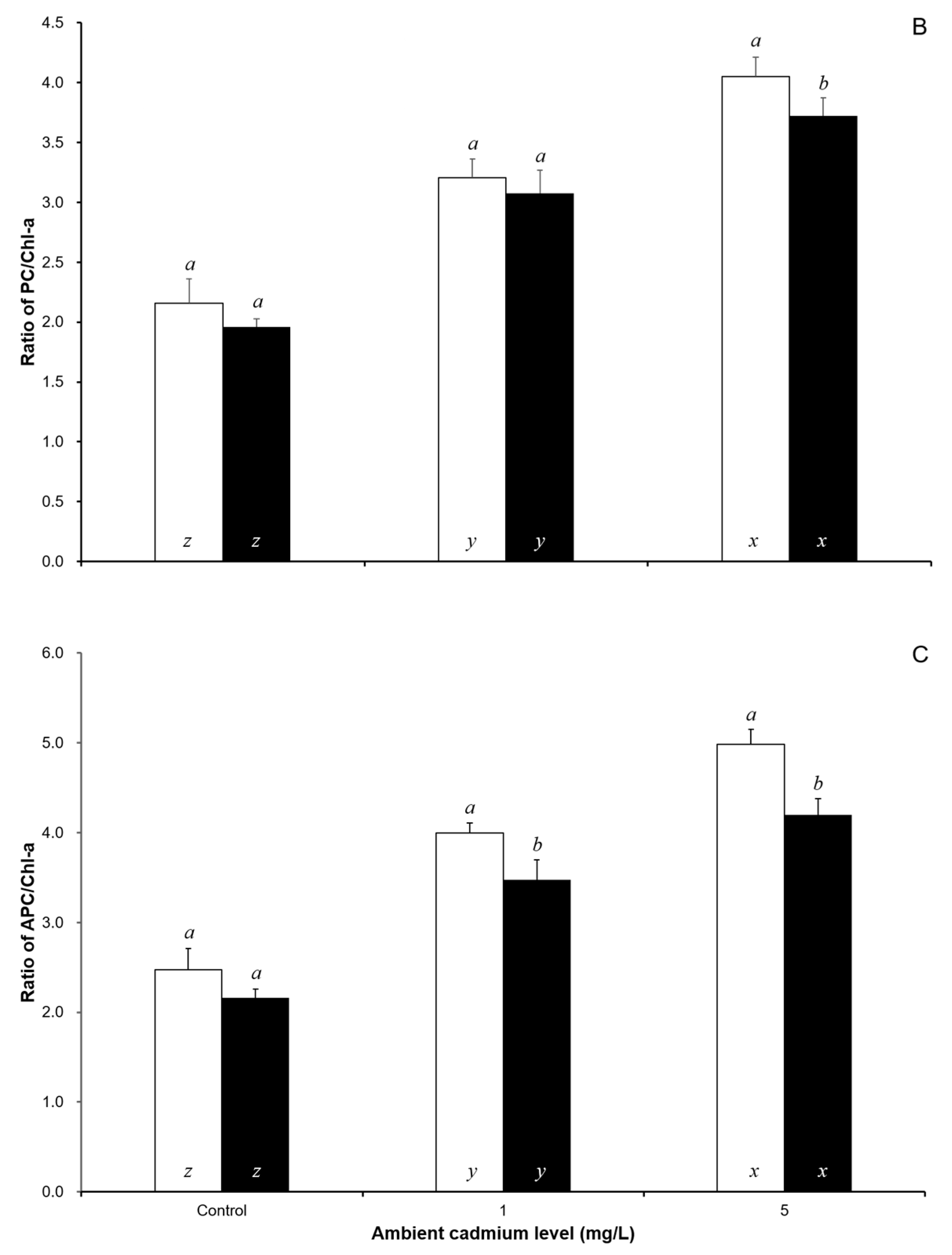
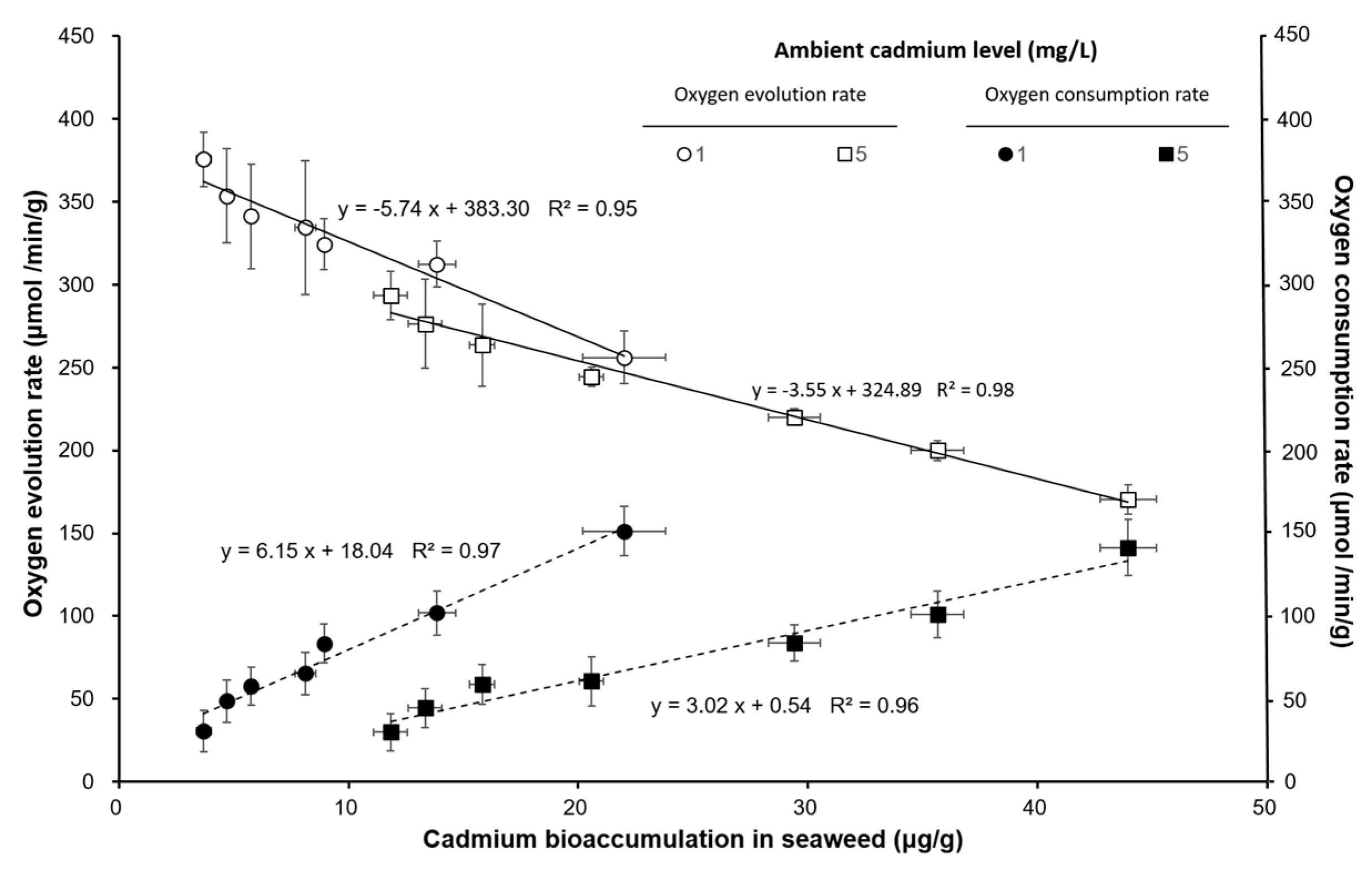
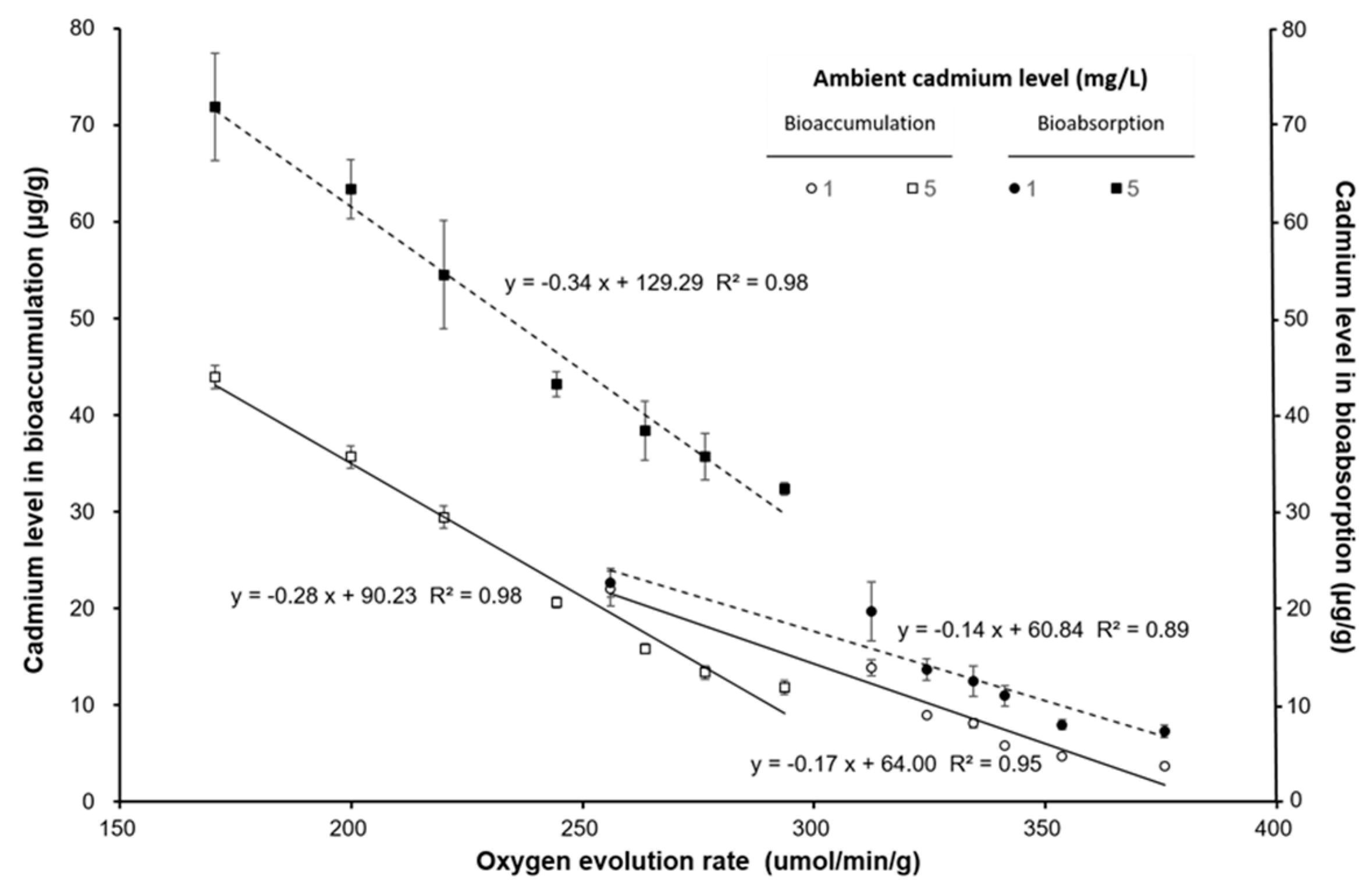
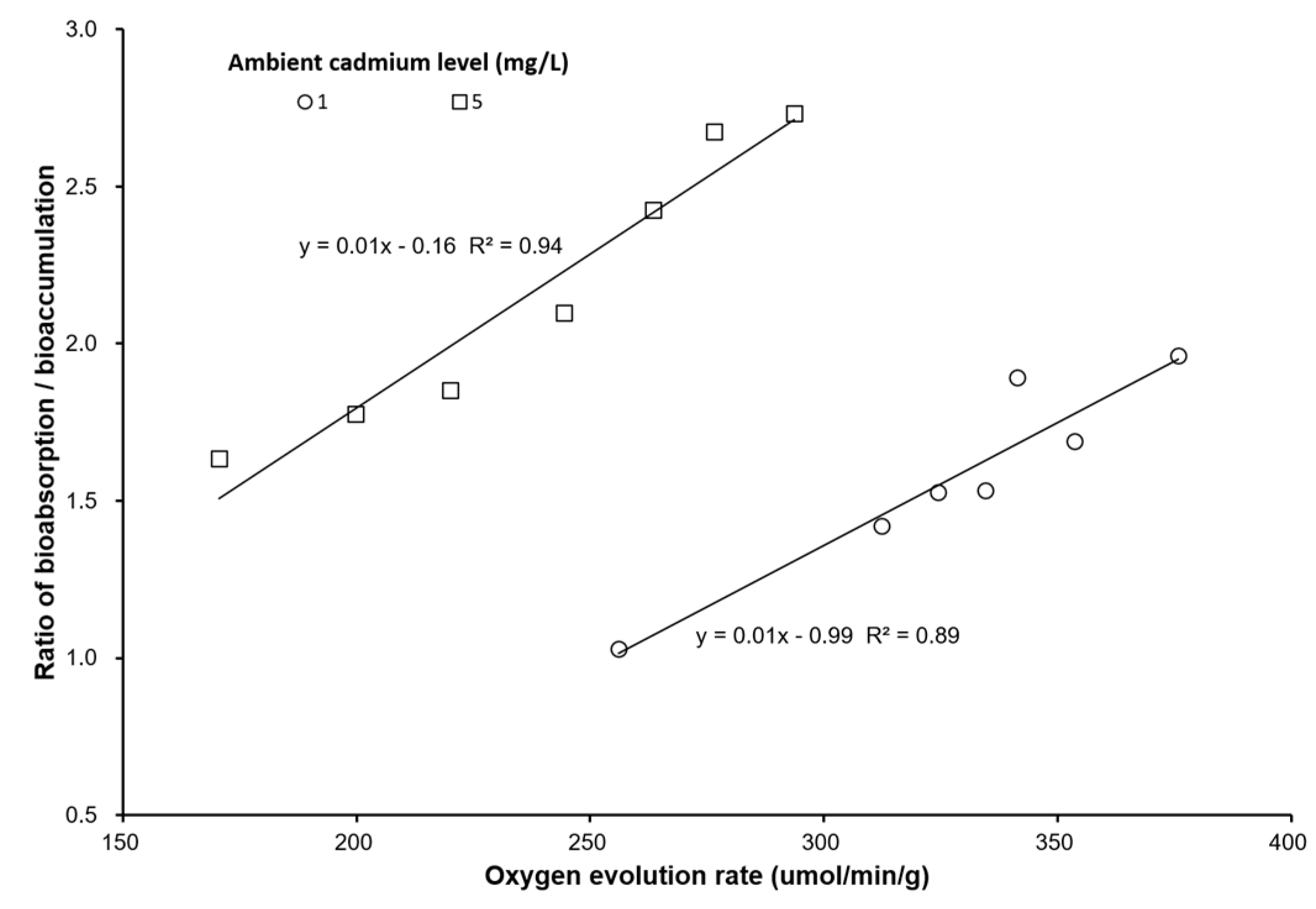
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yendo, K. Notes on algae new to Japan. VI. Bot. Mag. 1916, 30, 47–65. [Google Scholar] [CrossRef][Green Version]
- Rodríguez-Prieto, C.; Hommersand, M.H. Behaviour of the nuclei in pre- and post-fertilization stages in Kallymenia (Kallymeniaceae, Rhodophyta). Phycologia 2009, 48, 138–155. [Google Scholar] [CrossRef]
- Norris, R.E. Reproduction in Sarcodia dentata (Suhr) comb. nov. (Gigartinales, Rhodophyceae), with comments on the Sarcodiaceae. Br. Phycol. J. 1987, 22, 147–155. [Google Scholar] [CrossRef]
- Womersley, H.B.S. The Marine Benthic Flora of Southern Australia—Part IIIA; Australian Biological Resources Study: Canberra, Australia, 1994; p. 508. [Google Scholar]
- Renfro, W.C.; Fowler, S.W.; Heyraud, M.; La Rosa, J. Relative importance of food and water in long-term zinc-65 accumulation by marine biota. J. Fish. Res. Board Can. 1975, 32, 1339–1345. [Google Scholar] [CrossRef]
- Munshi, A.B.; Quan, S.Y.; Li, S.J. Acute toxicity of copper, cadmium and copper-cadmium mixture to the larvae of the shrimp Penaeus monodon. Pak. J. Sci. Ind. Res. 1996, 39, 68–71. [Google Scholar]
- Hsu, M.J.; Selvaraj, K.; Agoramoorthy, G. Taiwan’s industrial heavy metal pollution threatens terrestrial biota. Environ. Pollut. 2006, 143, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Rybak, A.; Messyasz, B.; Łeska, B. Freshwater Ulva (Chlorophyta) as a bioaccumulator of selected heavy metals (Cd, Ni and Pb) and alkaline earth metals (Ca and Mg). Chemosphere 2012, 89, 1066–1076. [Google Scholar] [CrossRef]
- Gresswell, T.; Simpson, S.L.; Smith, R.E.W.; Nugegoda, D.; Mazumder, D.; Twining, J. Bioaccumulation and retention kinetics of cadmium in the freshwater decapod Macrobrachium australiense. Aquat. Toxicol. 2014, 148, 174–183. [Google Scholar] [CrossRef]
- Monteiro, C.M.; Fonseca, S.C.; Castro, P.M.L.; Malcata, F.X. Toxicity of cadmium and zinc on two microalgae, Scenedesmus obliquus and Desmodesmus pleiomorphus, from Northern Portugal. J. Appl. Phycol. 2011, 23, 97–103. [Google Scholar] [CrossRef]
- Aguirre-von-Wobeser, E.; Figueroa, F.L. Photosynthesis and growth of red and green morphotypes of Kappaphycus alvarezii (Rhodophyta) from the Philippines. Mar. Biol. 2001, 138, 679–686. [Google Scholar] [CrossRef]
- Borlongan, I.A.; Gregory, N.N.; Shimada, S.; Terada, R. Photosynthetic performance of the red alga Solieria pacifica (Solieriaceae) from two different depths in the sublittoral waters of Kagoshima, Japan. J. Appl. Phycol. 2017, 29, 3077–3088. [Google Scholar] [CrossRef]
- Fukumoto, R.; Borlongan, I.A.; Nishihara, G.N.; Endo, H.; Terada, R. Effect of photosynthetically active radiation and temperature on the photosynthesis of two heteromorphic life history stages of a temperate edible brown alga, Cladosiphon umezakii (Chordariaceae, Ectocarpales), from Japan. J. Appl. Phycol. 2019, 31, 1259–1270. [Google Scholar] [CrossRef]
- Xia, J.R.; Li, Y.J.; Lu, J.; Chen, B. Effects of Copper and Cadmium on Growth, Photosynthesis, and Pigment Content in Gracilaria lemaneiformis. Bull. Environ. Contam. Toxicol. 2004, 73, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Dewez, D.; Geoffroy, L.; Vernet, G.; Popovic, R. Determination of photosynthetic and enzymatic biomarkers sensitivity used to evaluate toxic effects of copper and fludioxonil in alga Scenedesmus obliquus. Aquat. Toxicol. 2005, 74, 150–159. [Google Scholar] [CrossRef]
- Ji, Y.; Xie, X.J.; Wang, G.G. Effects of the heavy metal cadmium on photosynthetic activity and the xanthophyll cycle in Phaeodactylum tricornutum. J. Oceanol. Limnol. 2018, 36, 2194–2201. [Google Scholar] [CrossRef]
- Sunda, W.G.; Huntsman, S.A. Control of Cd concentrations in a coastal diatom by interactions among free ionic Cd, Zn, and Mn in seawater. Environ. Sci. Technol. 1998, 32, 2961–2968. [Google Scholar] [CrossRef]
- Reis, P.A.; Salgado, M.A.; Vasconcelos, V. Barnacles as biomonitors of metal contamination in coastal waters. Estuar. Coast. Shelf Sci. 2011, 93, 269–278. [Google Scholar] [CrossRef]
- Dos Santos, R.W.; Schmidt, E.C.; Martins, R.P.; Latini, A.; Maraschin, M.; Horta, P.A.; Bouzon, Z.L. Effects of Cadmium on Growth, Photosynthetic Pigments, Photosynthetic Performance, Biochemical Parameters and Structure of Chloroplasts in the Agarophyte Gracilaria domingensis (Rhodophyta, Gracilariales). Am. J. Plant. Sci. 2012, 3, 1077–1084. [Google Scholar] [CrossRef]
- Airs, R.L.; Temperton, B.; Sambles, C.; Farnham, G.; Skill, S.C.; Llewellyn, C.A. Chlorophyll f and chlorophyll d are produced in the cyanobacterium Chlorogloeopsis fritschii when cultured under natural light and near-infrared radiation. FEBS Lett. 2014, 588, 3770–3777. [Google Scholar] [CrossRef]
- Wolf, B.M.; Blankenship, R.E. Far-red light acclimation in diverse oxygenic photosynthetic organisms. Photosynth. Res. 2019, 142, 349–359. [Google Scholar] [CrossRef]
- Austin, P.A.; Ross, I.S.; Mills, J.D. Regulation of pigment content and enzyme activity in the cyanobacterium Nostoc sp. Mac grown in continuous light, a light-dark photoperiod, or darkness. Biochim. Biophys. Acta Bioenerg. 1996, 1277, 141–149. [Google Scholar] [CrossRef]
- Jiang, T.; Zhang, J.P.; Chang, W.R.; Liang, D.C. Crystal structure of R-phycocyanin and possible energy transfer pathways in the phycobilisome. Biophys. J. 2001, 81, 1171–1179. [Google Scholar] [CrossRef][Green Version]
- Kwak, M.J.; Je, S.M.; Cheng, H.C.; Seo, S.M.; Park, J.H.; Baek, S.G.; Khaine, I.; Lee, T.Y.; Jang, J.; Li, Y.; et al. Night Light-Adaptation Strategies for Photosynthetic Apparatus in Yellow-Poplar (Liriodendron tulipifera L.) Exposed to Artificial Night Lighting. Forests 2018, 9, 74. [Google Scholar] [CrossRef]
- Küpper, H.; Küpper, F.; Spiller, M. In situ detection of heavy metal substituted chlorophylls in water plants. Photosynth. Res. 1998, 58, 123–133. [Google Scholar] [CrossRef]
- Ma, R.; Lu, F.; Bi, Y.; Hu, Z. Effects of light intensity and quality on phycobiliprotein accumulation in the cyanobacterium Nostoc sphaeroides Kützing. Biotechnol. Lett. 2015, 37, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, T.; Plusquin, M.; Hogervorst, J.; Roels, H.A.; Celis, H.; Thijs, L.; angronsveld, J.; Van Hecke, E.; Staessen, J.A. Environmental exposure to cadmium and risk of cancer: A prospective population-based study. Lancet Oncol. 2006, 7, 119–126. [Google Scholar] [CrossRef]
- Zhou, W.; Juneau, P.; Qiu, B. Growth and photosynthetic responses of the bloom-forming cyanobacterium Microcystis aeruginosa to elevated levels of cadmium. Chemosphere 2006, 65, 1738–1746. [Google Scholar] [CrossRef]
- Boyle, E.A.; Sclater, F.; Edmond, J.M. On the marine geochemistry of cadmium. Nature 1976, 263, 42–44. [Google Scholar]
- Hsu, S.C.; Lin, F.J.; Jeng, W.L.; Tang, T.Y. Spatial distribution of cadmium over a cyclonic eddy in the southern East China Sea. J. Mar. Syst. 2003, 39, 153–166. [Google Scholar] [CrossRef]
- Bruland, K.W.; Knauer, G.A.; Martin, J.H. Cadmium in northeast Pacific waters. Limnol. Oceanogr. 1978, 23, 618–625. [Google Scholar] [CrossRef]
- Licata, P.; Trombetta, D.; Cristani, M.; Martino, D.; Naccari, F. Organochlorine compounds and heavy metals in the soft tissue of the mussel Mytilus galloprovincialis collected from Lake Faro (Sicily, Italy). Environ. Int. 2004, 30, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Fargašová, A.; Bumbálová, A.; Havránek, E. Ecotoxicological effects and uptake of metals (Cu+, Cu2+, Mn2+, Mo6+, Ni2+, V5+) in freshwater alga Scenedesmus quadricauda. Chemosphere 1999, 38, 1165–1173. [Google Scholar] [CrossRef]
- Payne, C.D.; Price, N.M. Effects of cadmium toxicity on growth and elemental composition of marine phytoplankton. J. Phycol. 1999, 35, 293–302. [Google Scholar] [CrossRef]
- Kola, H.; Wilkinson, K.J. Cadmium uptake by a green alga can be predicted by equilibrium modelling. Environ. Sci. Technol. 2005, 39, 3040–3047. [Google Scholar] [CrossRef]
- Miao, X.; Wu, Q. Biodiesel production from heterotrophic microalgal oil. Bioresour. Technol. 2006, 97, 841–846. [Google Scholar] [CrossRef]
- Kapkov, V.I.; Belenikina, O.A.; Fedorov, V.D. Effect of heavy metals on marine phytoplankton. Mosc. Univ. Biol. Sci. Bull. 2011, 66, 32–36. [Google Scholar] [CrossRef]
- Echeveste, P.; Silva, J.C.; Lombardi, A.T. Cu and Cd affect distinctly the physiology of a cosmopolitan tropical freshwater phytoplankton. Ecotoxicol. Environ. Saf. 2017, 143, 228–235. [Google Scholar] [CrossRef]
- Zeraatkar, A.K.; Ahmadzadeh, H.; Talebi, A.F.; Moheimani, N.R.; McHenry, M.P. Potential use of algae for heavy metal bioremediation, a critical review. J. Environ. Manag. 2016, 181, 817–831. [Google Scholar] [CrossRef]
- Davis, T.A.; Volesky, B.; Vieira, R. Sargassum seaweed as biosorbent for heavy metals. Water Res. 2000, 34, 4270–4278. [Google Scholar] [CrossRef]
- Romera, E.; González, F.; Ballester, A.; Blázquez, M.; Munoz, J. Comparative study of biosorption of heavy metals using different types of algae. Bioresour. Technol. 2007, 98, 3344–3353. [Google Scholar] [CrossRef] [PubMed]
- Perales-Vela, H.V.; Pena-Casro, J.M.; Canizares-Villanueva, R.O. Heavy metal detoxification in eukaryotic microalgae. Chemosphere 2006, 64, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wilde, K.L.; Stauber, J.L.; Markich, S.J.; Franklin, N.M.; Brown, P.L. The effect of pH on the uptake and toxicity of copper and zinc in a tropical freshwater alga (Chlorella sp.). Arch. Environ. Contam. Toxicol. 2006, 51, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Macfie, S.M.; Tarmohamed, Y.; Welbourn, P.M. Effects of cadmium, cobalt, copper, and nickel on growth of the green alga Chlamydomonas reinhardtii: The influences of the cell wall and pH. Arch. Environ. Contam. Toxicol. 1994, 27, 454–458. [Google Scholar] [CrossRef]
- Bastosa, E.; Schneider, M.; de Quadros, D.P.C.; Welz, B.; Batista, M.B.; Horta, P.A.; Rörig, L.R.; Barufi, J.B. Phytoremediation potential of Ulva ohnoi (Chlorophyta): Influence of temperature and salinity on the uptake efficiency and toxicity of cadmium. Ecotoxicol. Environ. Saf. 2019, 174, 334–343. [Google Scholar] [CrossRef]
- Perreault, F.; Dionne, J.; Didur, O.; Juneau, P.; Popovic, R. Effect of cadmium on photosystem II activity in Chlamydomonas reinhardtii: Alteration of OJIP fluorescence transients indicating the change of apparent activation energies within photosystem II. Photosynth. Res. 2011, 107, 151–157. [Google Scholar] [CrossRef]
- Parmar, P.; Kumari, N.; Sharma, V. Structural and functional alterations in photosynthetic apparatus of plants under cadmium stress. Bot. Stud. 2013, 54, 45. [Google Scholar] [CrossRef]
- Sigfridsson, K.G.V.; Bernát, G.; Mamedov, F.; Styring, S. Molecular interference of Cd2+ with Photosystem II. Biochim. Biophys. Acta Bioenerg. 2004, 1659, 19–31. [Google Scholar] [CrossRef]
- Lu, F.; Dind, G.; Liu, W.; Zhan, D.M.; Wu, H.Y.; Guo, W. Comparative study of responses in the brown algae Sargassum thunbergii to zinc and cadmium stress. J. Oceanol. Limn. 2018, 36, 933–941. [Google Scholar] [CrossRef]
- Blot, N.; Wu, X.J.; Thomas, J.C.; Zhang, J.; Garczarek, L.; Bohm, S. Phycourobilin in trichromatic phycocyanin from oceanic cyanobacteria is formed post-translationally by a phycoerythrobilin lyase-isomerase. J. Biol. Chem. 2009, 284, 9290–9298. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, R.M.; de Morais, A.M. Health applications of bio-active compounds from marine microalgae. Life Sci. 2013, 10, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Ikeuchi, M. Phycobilisome: Architecture of alight-harvesting supercomplex. Photosynth. Res. 2013, 116, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Mulders, K.J.M.; Lamers, P.P.; Martens, D.E.; Wijffels, R.H. Phototrophic pigment production with microalgae: Biological constraints and opportunities. J. Phycol. 2014, 50, 229–242. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, T.-W.; Tseng, C.-C.; Cai, M.; Chen, K.; Cheng, S.-Y.; Wang, J. Effects of Cadmium on Bioaccumulation, Bioabsorption, and Photosynthesis in Sarcodia suiae. Int. J. Environ. Res. Public Health 2020, 17, 1294. https://doi.org/10.3390/ijerph17041294
Han T-W, Tseng C-C, Cai M, Chen K, Cheng S-Y, Wang J. Effects of Cadmium on Bioaccumulation, Bioabsorption, and Photosynthesis in Sarcodia suiae. International Journal of Environmental Research and Public Health. 2020; 17(4):1294. https://doi.org/10.3390/ijerph17041294
Chicago/Turabian StyleHan, Tai-Wei, Chung-Chih Tseng, Minggang Cai, Kai Chen, Sha-Yen Cheng, and Jun Wang. 2020. "Effects of Cadmium on Bioaccumulation, Bioabsorption, and Photosynthesis in Sarcodia suiae" International Journal of Environmental Research and Public Health 17, no. 4: 1294. https://doi.org/10.3390/ijerph17041294
APA StyleHan, T.-W., Tseng, C.-C., Cai, M., Chen, K., Cheng, S.-Y., & Wang, J. (2020). Effects of Cadmium on Bioaccumulation, Bioabsorption, and Photosynthesis in Sarcodia suiae. International Journal of Environmental Research and Public Health, 17(4), 1294. https://doi.org/10.3390/ijerph17041294