Comparison of Habitat Suitability Models for Haemaphysalis longicornis Neumann in North America to Determine Its Potential Geographic Range

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
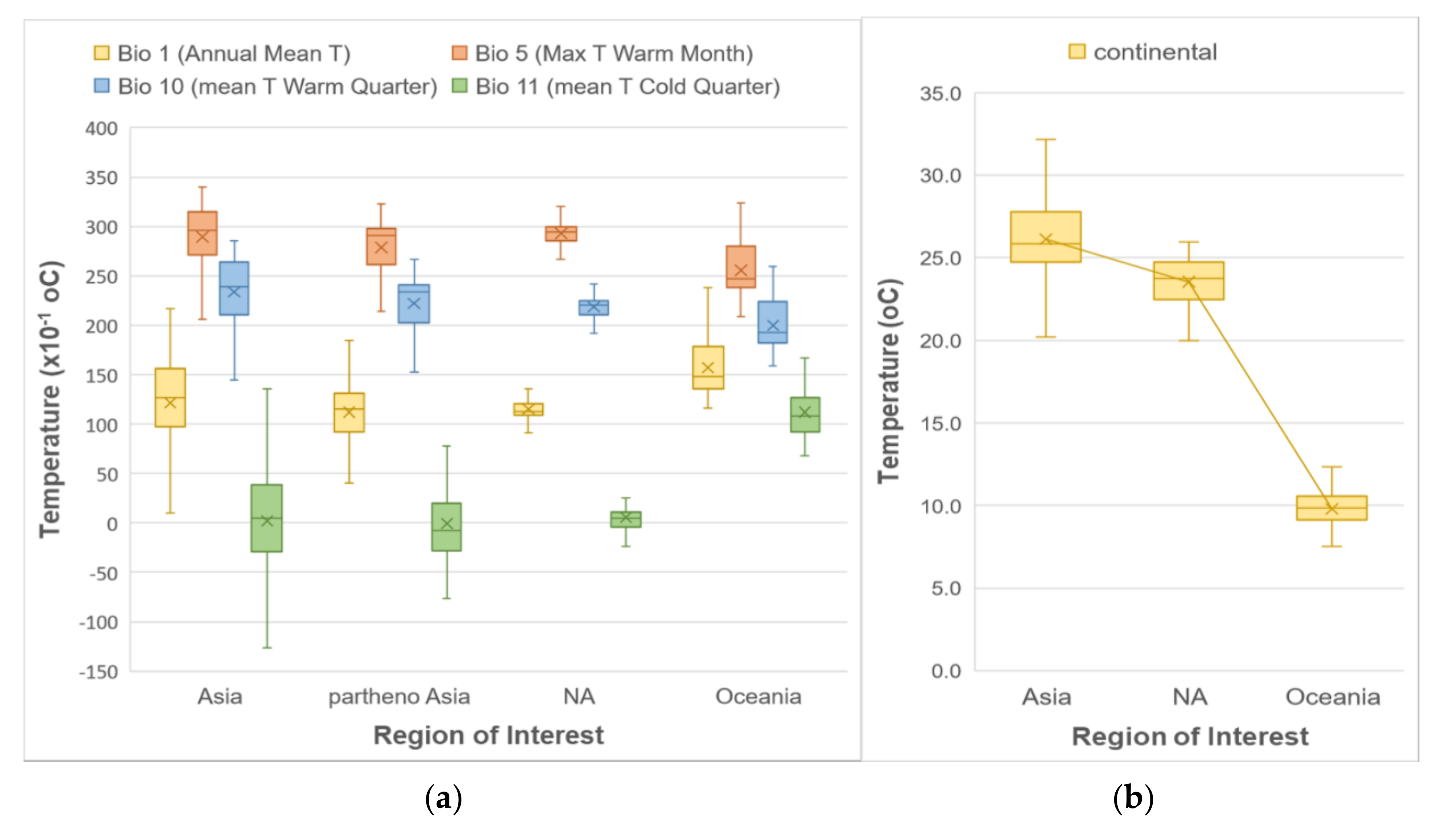
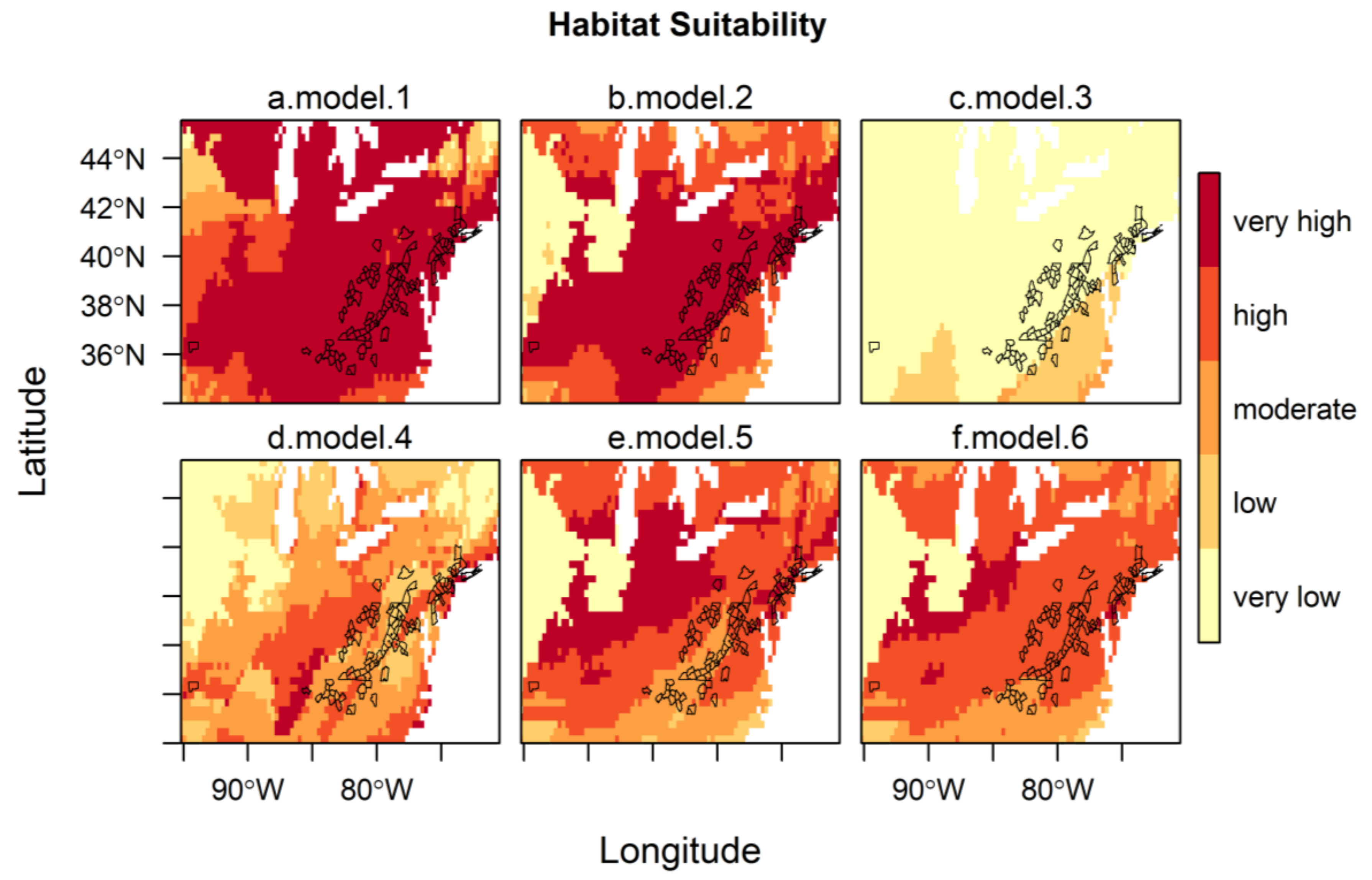
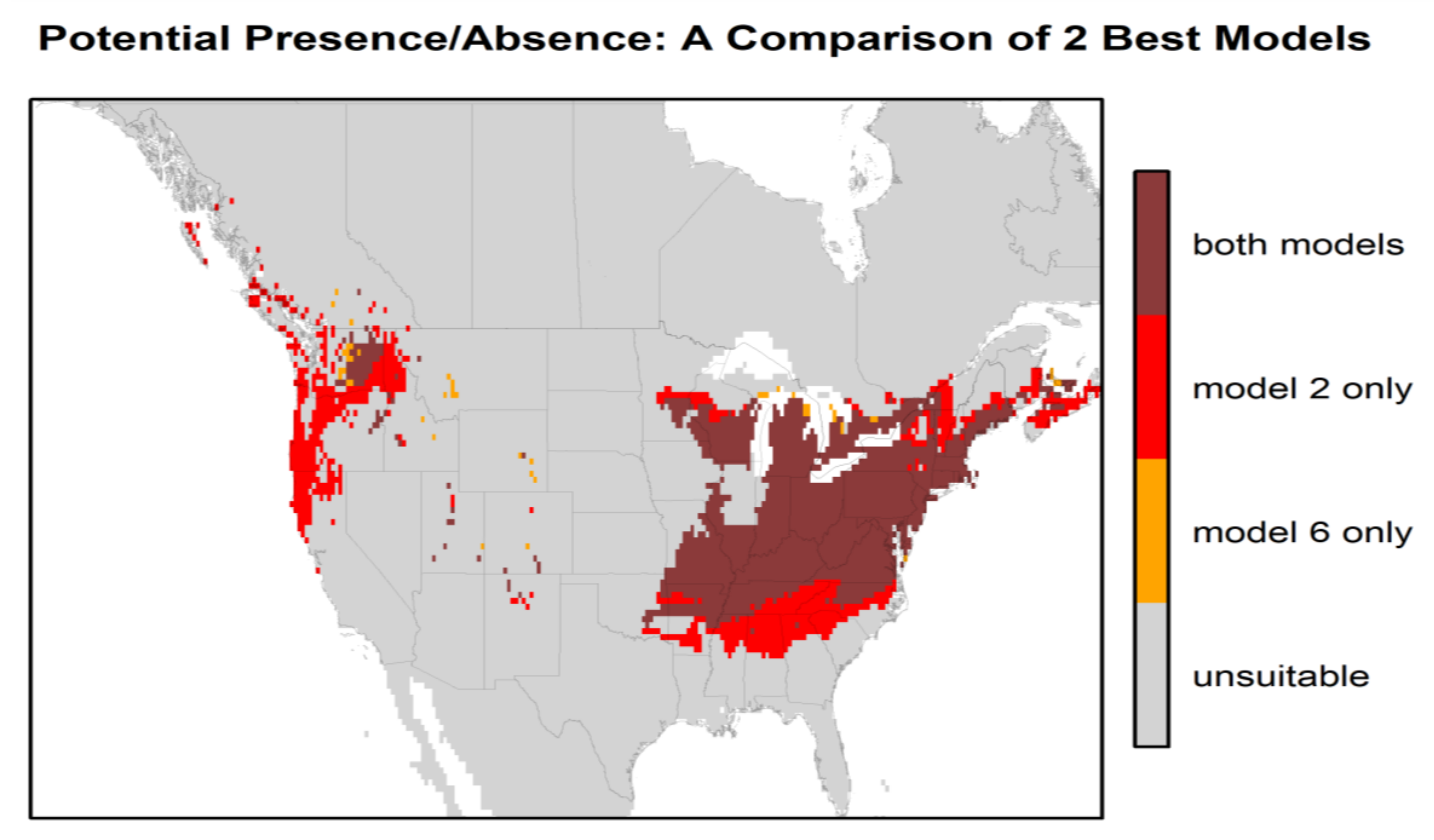
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoogstraal, H.; Roberts, F.H.S.; Kohls, G.M.; Tipton, V.J. Review of Haemaphysalis (Kaiseriana) longicornis Neumann (Resurrected) of Australia, New Zealand, New Caledonia, Fiji, Japan, Korea, and Northeastern China and USSR, and Its Parthenogenetic and Bisexual Populations (Ixodoidea, Ixodidae). J. Parasitol. 1968, 54, 1197–1213. [Google Scholar] [CrossRef] [PubMed]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Heath, A.C.G. Biology, ecology and distribution of the tick, Haemaphysalis longicornis Neumann (Acari: Ixodidae) in New Zealand. N. Z. Vet. J. 2016, 64, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Jiang, Z.; Shin, W.-J.; Jung, J.U. Severe Fever with Thrombocytopenia Syndrome Virus NSs Interacts with TRIM21 To Activate the p62-Keap1-Nrf2 Pathway. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-J.; Liang, M.-F.; Zhang, S.-Y.; Liu, Y.; Li, J.-D.; Sun, Y.-L.; Zhang, L.; Zhang, Q.-F.; Popov, V.L.; Li, C.; et al. Fever with Thrombocytopenia Associated with a Novel Bunyavirus in China. N. Engl. J. Med. 2011, 364, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Yi, J.; Kim, G.; Choi, S.J.; Jun, K.I.; Kim, N.-H.; Choe, P.G.; Kim, N.-J.; Lee, J.-K.; Oh, M. Severe Fever with Thrombocytopenia Syndrome, South Korea, 2012. Emerg. Infect. Dis. 2013, 19, 1892–1894. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Maeda, K.; Suzuki, T.; Ishido, A.; Shigeoka, T.; Tominaga, T.; Kamei, T.; Honda, M.; Ninomiya, D.; Sakai, T.; et al. The First Identification and Retrospective Study of Severe Fever with Thrombocytopenia Syndrome in Japan. J. Infect. Dis. 2014, 209, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-W.; Ryou, J.; Choi, W.-Y.; Han, M.-G.; Lee, W.-J. Epidemiological and Clinical Features of Severe Fever with Thrombocytopenia Syndrome during an Outbreak in South Korea, 2013–2015. Am. J. Trop. Med. Hyg. 2016, 95, 1358–1361. [Google Scholar] [CrossRef]
- Brault, A.C.; Savage, H.M.; Duggal, N.K.; Eisen, R.J.; Staples, J.E. Heartland Virus Epidemiology, Vector Association, and Disease Potential. Viruses 2018, 10, 498. [Google Scholar] [CrossRef]
- McMullan, L.K.; Folk, S.M.; Kelly, A.J.; MacNeil, A.; Goldsmith, C.S.; Metcalfe, M.G.; Batten, B.C.; Albarino, C.G.; Zaki, S.R.; Rollin, P.E.; et al. A New Phlebovirus Associated with Severe Febrile Illness in Missouri. N. Engl. J. Med. 2012, 367, 834–841. [Google Scholar] [CrossRef]
- Mahara, F. Japanese spotted fever: Report of 31 cases and review of the literature. Emerg. Infect. Dis. 1997, 3, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Hammer, J.F.; Emery, D.; Bogema, D.R.; Jenkins, C. Detection of Theileria orientalis genotypes in Haemaphysalis longicornis ticks from southern Australia. Parasit. Vectors 2015, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, K.E.; Summers, S.R.; Heath, A.C.G.; McFadden, A.M.J.; Pulford, D.J.; Tait, A.B.; Pomroy, W.E. Using a rule-based envelope model to predict the expansion of habitat suitability within New Zealand for the tick Haemaphysalis longicornis, with future projections based on two climate change scenarios. Vet. Parasitol. 2017, 243, 226–234. [Google Scholar] [CrossRef]
- Chu, C.; Jiang, B.-G.; Liu, W.; Zhao, Q.-M.; Wu, X.-M.; Zhang, P.-H.; Zhan, L.; Yang, H.; Cao, W.-C. Presence of pathogenic Borrelia burgdorferi sensu lato in ticks and rodents in Zhejiang, south-east China. J. Med. Microbiol. 2008, 57, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Sun, J.; Yan, J.; Wang, C.; Zhang, Z.; Zhao, L.; Han, H.; Tong, Z.; Liu, M.; Wu, Y.; et al. Detection of a Novel Ehrlichia Species in Haemaphysalis longicornis Tick from China. Vector-Borne Zoonotic Dis. 2016, 16, 363–367. [Google Scholar] [CrossRef]
- Rainey, T.; Occi, J.L.; Robbins, R.G.; Egizi, A. Discovery of Haemaphysalis longicornis (Ixodida: Ixodidae) Parasitizing a Sheep in New Jersey, United States. J. Med. Entomol. 2018, 55, 757–759. [Google Scholar] [CrossRef] [PubMed]
- Oakes, V.J.; Yabsley, M.J.; Schwartz, D.; LeRoith, T.; Bissett, C.; Broaddus, C.; Schlater, J.L.; Todd, S.M.; Boes, K.M.; Brookhart, M.; et al. Theileria orientalis Ikeda Genotype in Cattle, Virginia, USA. Emerg. Infect. Dis. J. 2019, 25. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.T.; Dominguez, K.; Cleveland, C.A.; Dergousoff, S.J.; Doi, K.; Falco, R.C.; Greay, T.; Irwin, P.; Lindsay, L.R.; Liu, J.; et al. Molecular Characterization of Haemaphysalis Species and a Molecular Genetic Key for the Identification of Haemaphysalis of North America. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- USDA National Haemaphysalis longicornis (Asian Longhorned Tick) Situation Report, U.S. Department of Agriculture’s Animal and Plant Health Inspection Service. 2020. Available online: https://www.aphis.usda.gov/animal_health/animal_diseases/tick/downloads/longhorned-tick-sitrep.pdf (accessed on 23 October 2020).
- Hutcheson, H.J.; Dergousoff, S.J.; Lindsay, L.R. Haemaphysalis longicornis: A tick of considerable veterinary importance, now established in North America. Can. Vet. J. 2019, 60, 27–28. [Google Scholar] [CrossRef] [PubMed]
- Public Health Ontario. The Asian Longhorned Tick: Assessing Public Health Implications for Ontario; Ontario Agency for Health Protection and Promotion (Public Health Ontario): Toronto, ON, Canada, 2019. [Google Scholar]
- Wormser, G.P.; McKenna, D.; Piedmonte, N.; Vinci, V.; Egizi, A.M.; Backenson, B.; Falco, R.C. First Recognized Human Bite in the United States by the Asian Longhorned Tick, Haemaphysalis longicornis. Clin. Infect. Dis. 2020, 70, 314–316. [Google Scholar] [CrossRef]
- Oliver, J.H.; Tanaka, K.; Sawada, M. Cytogenetics of ticks (Acari: Ixodoidea). Chromosoma 1973, 42, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Egizi, A.; Bulaga-Seraphin, L.; Alt, E.; Bajwa, W.I.; Bernick, J.; Bickerton, M.; Campbell, S.R.; Connally, N.; Doi, K.; Falco, R.C.; et al. First glimpse into the origin and spread of the Asian longhorned tick, Haemaphysalis longicornis, in the United States. Zoonoses Public Health 2020, 67, 637–650. [Google Scholar] [CrossRef]
- Herrin, C.S.; Oliver Jr, J.H. Numerical taxonomic studies of parthenogenetic and bisexual populations of Haemaphysalis longicornis and related species (Acari: Ixodidae). J. Parasitol. 1974, 1025–1036. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, X.; Bu, F.; Yang, X.; Liu, J. Morphological, biological and molecular characteristics of bisexual and parthenogenetic Haemaphysalis longicornis. Vet. Parasitol. 2012, 189, 344–352. [Google Scholar] [CrossRef]
- Magori, K. Preliminary prediction of the potential distribution and consequences of Haemaphysalis longicornis using a simple rule-based climate envelope model. bioRxiv 2018, 389940. [Google Scholar] [CrossRef]
- Rochlin, I. Modeling the Asian Longhorned Tick (Acari: Ixodidae) Suitable Habitat in North America. J. Med. Entomol. 2018, 56, 384–391. [Google Scholar] [CrossRef]
- Raghavan, R.K.; Barker, S.C.; Cobos, M.E.; Barker, D.; Teo, E.J.M.; Foley, D.H.; Nakao, R.; Lawrence, K.; Heath, A.C.G.; Peterson, A.T. Potential Spatial Distribution of the Newly Introduced Long-horned Tick, Haemaphysalis longicornis in North America. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zheng, D.; Ian, Y.T.; Li, S. A dataset of distribution and diversity of ticks in China. Sci. Data 2019, 6, 105. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Title, P.O.; Bemmels, J.B. ENVIREM: An expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 2018, 41, 291–307. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Hijmans, R.J. {raster: Geographic Data Analysis and Modeling}, R package version 3.1-5; 2020; Available online: https://CRAN.R-project.org/package=raster (accessed on 4 November 2020).
- Bivand, R.; Keitt, T.; Rowlingson, B. rgdal: Bindings for the “Geospatial” Data Abstraction Library, R package version 1.4-8; 2019; Available online: https://CRAN.R-project.org/package=rgdal (accessed on 4 November 2020).
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. dismo: Species Distribution Modeling, R package version 1.1-4; 2017; Available online: https://CRAN.R-project.org/package=dismo (accessed on 4 November 2020).
- Vollering, J.; Halvorsen, R.; Mazzoni, S. The MIAmaxent R package: Variable transformation and model selection for species distribution models. Ecol. Evol. 2019, 9, 12051–12068. [Google Scholar] [CrossRef] [PubMed]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2019, 43–57. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference: Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental Niche Equivalency Versus Conservatism: Quantitative Approaches to Niche Evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef]
- Zuliani, A.; Massolo, A.; Lysyk, T.; Johnson, G.; Marshall, S.; Berger, K.; Cork, S.C. Modelling the Northward Expansion of Culicoides sonorensis (Diptera: Ceratopogonidae) under Future Climate Scenarios. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Zhao, L.; Li, J.; Cui, X.; Jia, N.; Wei, J.; Xia, L.; Wang, H.; Zhou, Y.; Wang, Q.; Liu, X.; et al. Distribution of Haemaphysalis longicornis and associated pathogens: Analysis of pooled data from a China field survey and global published data. Lancet Planet. Health 2020, 4, e320–e329. [Google Scholar] [CrossRef]
- Miao, D.; Dai, K.; Zhao, G.-P.; Li, X.-L.; Shi, W.-Q.; Zhang, J.S.; Yang, Y.; Liu, W.; Fang, L.-Q. Mapping the global potential transmission hotspots for severe fever with thrombocytopenia syndrome by machine learning methods. Emerg. Microbes Infect. 2020, 9, 817–826. [Google Scholar] [CrossRef]
- Peng, S.-H.; Yang, S.-L.; Tang, S.-E.; Wang, T.-C.; Hsu, T.-C.; Su, C.-L.; Chen, M.-Y.; Shimojima, M.; Yoshikawa, T.; Shu, P.-Y. Human Case of Severe Fever with Thrombocytopenia Syndrome Virus Infection, Taiwan, 2019. Emerg. Infect. Dis. 2020, 26, 1612–1614. [Google Scholar] [CrossRef]
- Tran, X.C.; Yun, Y.; Van An, L.; Kim, S.-H.; Thao, N.T.P.; Man, P.K.C.; Yoo, J.R.; Heo, S.T.; Cho, N.-H.; Lee, K.H. Endemic Severe Fever with Thrombocytopenia Syndrome, Vietnam. Emerg. Infect. Dis. 2019, 25, 1029–1031. [Google Scholar] [CrossRef] [PubMed]
- Ben Beard, C.; Occi, J.; Bonilla, D.L.; Egizi, A.M.; Fonseca, D.M.; Mertins, J.W.; Backenson, B.P.; Bajwa, W.I.; Barbarin, A.M.; Bertone, M.A.; et al. Multistate Infestation with the Exotic Disease-Vector Tick Haemaphysalis longicornis - United States, August 2017-September 2018. Mmwr-Morb. Mortal. Wkly. Rep. 2018, 67, 1310–1313. [Google Scholar] [CrossRef]
- Stanley, H.M.; Ford, S.L.; Snellgrove, A.N.; Hartzer, K.; Smith, E.B.; Krapiunaya, I.; Levin, M.L. The Ability of the Invasive Asian Longhorned Tick Haemaphysalis longicornis (Acari: Ixodidae) to Acquire and Transmit Rickettsia rickettsii (Rickettsiales: Rickettsiaceae), the Agent of Rocky Mountain Spotted Fever, Under Laboratory Conditions. J. Med. Entomol. 2020. [Google Scholar] [CrossRef]
- Lawrence, K.E.; Lawrence, B.L.; Hickson, R.E.; Hewitt, C.A.; Gedye, K.R.; Fermin, L.M.; Pomroy, W.E. Associations between Theileria orientalis Ikeda type infection and the growth rates and haematocrit of suckled beef calves in the North Island of New Zealand. N. Z. Vet. J. 2019, 67, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Tufts, D.M.; VanAcker, M.C.; Fernandez, M.P.; DeNicola, A.; Egizi, A.; Diuk-Wasser, M.A. Distribution, Host-Seeking Phenology, and Host and Habitat Associations of Haemaphysalis longicornis Ticks, Staten Island, New York, USA. Emerg. Infect. Dis. 2019, 25, 792–796. [Google Scholar] [CrossRef]
- White, S.A.; Bevins, S.N.; Ruder, M.G.; Shaw, D.; Vigil, S.L.; Randall, A.; Deliberto, T.J.; Dominguez, K.; Thompson, A.T.; Mertins, J.W.; et al. Surveys for ticks on wildlife hosts and in the environment at Asian longhorned tick (Haemaphysalis longicornis)-positive sites in Virginia and New Jersey, 2018. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Neilson, F.J.A. An investigation into the ecology, biology, distribution and control of Haemaphysalis longicornis Neumann, 1901: A thesis presented in partial fulfilment of the requirements for the degree ofMaster of Veterinary Science at Massey University. Master’s Thesis, Massey University, Palmerston North, New Zealand, 1980. [Google Scholar]
- Zheng, H.; Yu, Z.; Zhou, L.; Yang, X.; Liu, J. Seasonal abundance and activity of the hard tick Haemaphysalis longicornis (Acari: Ixodidae) in North China. Exp. Appl. Acarol. 2012, 56, 133–141. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Zone of Influence | GEZ | bio 1 | bio 12 | bio 5 | Features (rm) |
|---|---|---|---|---|---|---|
| 1 | Entire distribution | 10.34 | 77.63 | 9.62 | 2.40 | LQ (4.0) |
| 2 | East Asia | 33.60 | 41.20 | 13.46 | 11.74 | LQ (0.5) |
| 3 | Oceania | 5.33 | 6.12 | 13.73 | 74.82 | LQ (1.0) |
| 4 | Native parthenogenetic | 26.54 | 51.48 | 5.57 | 16.41 | LQ (0.5) |
| Model | Model on | AUC | Correlation | Parameters | AICc | ΔΑΙCc |
|---|---|---|---|---|---|---|
| 1 | Entire range | 0.95 | 0.64 | 11 | 2420.7 | 17.5 |
| 2 | East Asia | 0.97 | 0.68 | 8 | 2403.2 | 0 |
| 3 | Oceania | 0.87 | 0.16 | 9 | 2662.3 | 259 |
| 4 | Parthenogenetic | 0.75 | 0.22 | 8 | 2631.3 | 228 |
| 5 | East Asia | 0.95 | 0.62 | 8 | 2421.6 | 18.3 |
| 6 | East Asia | 0.96 | 0.64 | 8 | 2418.7 | 15.5 |
| Model 5 | Model 6 | ||
|---|---|---|---|
| GEZ | 45.44 | GEZ | 43.01 |
| bio 1 | 27.72 | bio 10 | 10.63 |
| continentality | 26.84 | bio 11 | 46.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Namgyal, J.; Couloigner, I.; Lysyk, T.J.; Dergousoff, S.J.; Cork, S.C. Comparison of Habitat Suitability Models for Haemaphysalis longicornis Neumann in North America to Determine Its Potential Geographic Range. Int. J. Environ. Res. Public Health 2020, 17, 8285. https://doi.org/10.3390/ijerph17218285
Namgyal J, Couloigner I, Lysyk TJ, Dergousoff SJ, Cork SC. Comparison of Habitat Suitability Models for Haemaphysalis longicornis Neumann in North America to Determine Its Potential Geographic Range. International Journal of Environmental Research and Public Health. 2020; 17(21):8285. https://doi.org/10.3390/ijerph17218285
Chicago/Turabian StyleNamgyal, Jamyang, Isabelle Couloigner, Tim J. Lysyk, Shaun J. Dergousoff, and Susan C. Cork. 2020. "Comparison of Habitat Suitability Models for Haemaphysalis longicornis Neumann in North America to Determine Its Potential Geographic Range" International Journal of Environmental Research and Public Health 17, no. 21: 8285. https://doi.org/10.3390/ijerph17218285
APA StyleNamgyal, J., Couloigner, I., Lysyk, T. J., Dergousoff, S. J., & Cork, S. C. (2020). Comparison of Habitat Suitability Models for Haemaphysalis longicornis Neumann in North America to Determine Its Potential Geographic Range. International Journal of Environmental Research and Public Health, 17(21), 8285. https://doi.org/10.3390/ijerph17218285