Hormonal Changes in High-Level Aerobic Male Athletes during a Sports Season

 , ,
, ,  ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Nutritional Evaluation
2.3. Anthropometrics Measurements
2.4. Exercise Test until Exhaustion
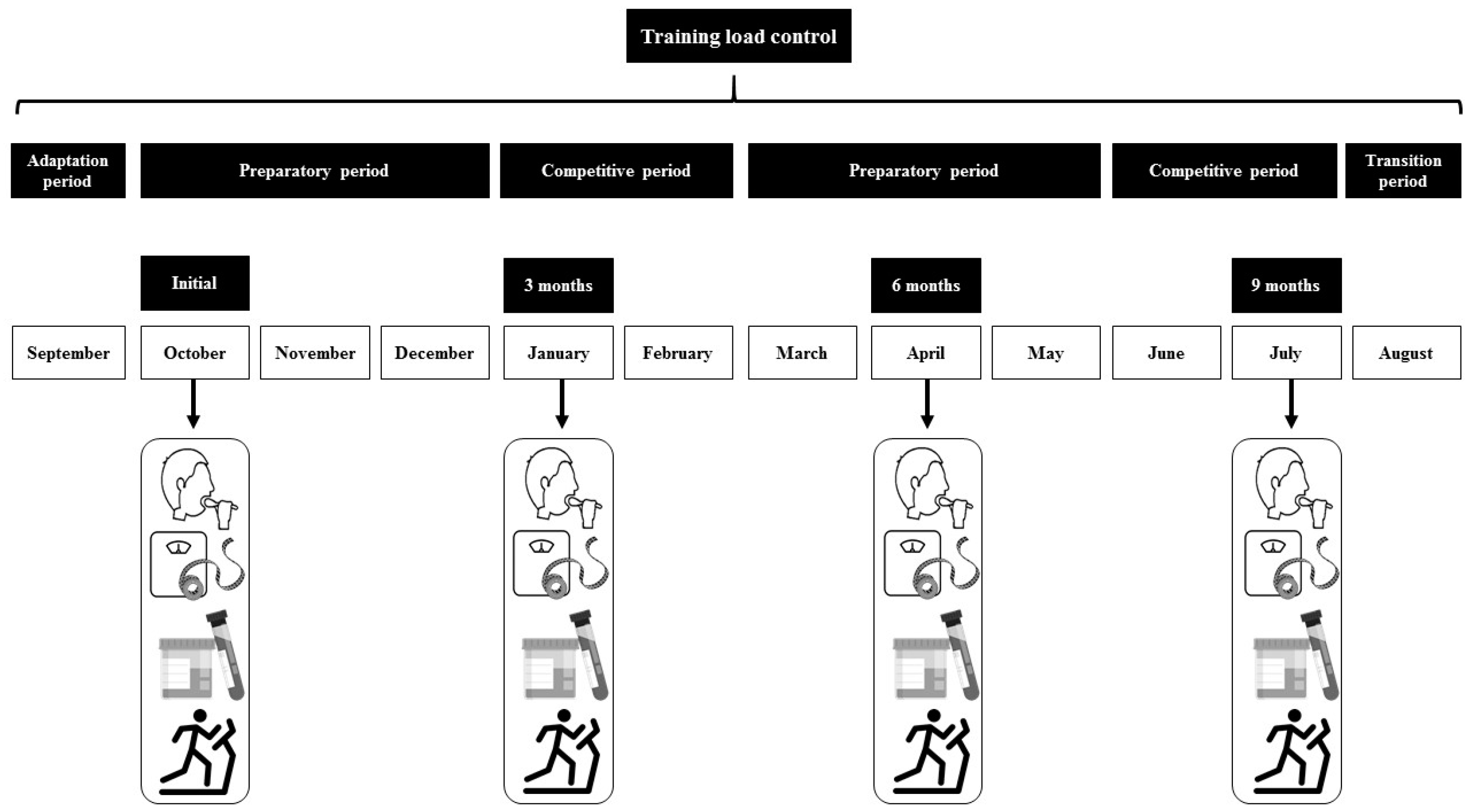
2.5. Training Characteristics
2.6. Sample Collection
2.7. Analytical Determination
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zinner, C.; Wahl, P.; Achtzehn, S.; Reed, J.L.; Mester, J. Acute hormonal responses before and after 2 weeks of HIT in well trained junior triathletes. Int. J. Sports Med. 2014, 35, 316–322. [Google Scholar] [CrossRef]
- Hackney, A.C.; Lane, A.R. Exercise and the regulation of endocrine hormones. In Molecular and Cellular Regulation of Adaptation to Exercise; Bouchard, C., Ed.; Elsevier Academic Press: San Diego, CA, USA, 2015; Volume 135, pp. 293–311. ISBN 1877-1173. [Google Scholar]
- Brownlee, K.K.; Moore, A.W.; Hackney, A.C. Relationship between circulating cortisol and testosterone: Influence of physical exercise. J. Sports Sci. Med. 2005, 4, 76. [Google Scholar]
- Duclos, M.; Tabarin, A. Exercise, training, and the hypothalamo–pituitary–adrenal axis. In Hormone Use and Abuse by Athletes; Springer: Boston, MA, USA, 2011; pp. 9–15. [Google Scholar]
- Viru, A.; Viru, M. Cortisol-essential adaptation hormone in exercise. Int. J. Sports Med. 2004, 25, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.; Lane, A.R.; Hackney, A.C. Cortisol and testosterone dynamics following exhaustive endurance exercise. Eur. J. Appl. Physiol. 2016, 116, 1503–1509. [Google Scholar] [CrossRef]
- Zitzmann, M.; Nieschlag, E. Testosterone levels in healthy men and the relation to behavioural and physical characteristics: Facts and constructs. Eur. J. Endocrinol. 2001, 144, 183–197. [Google Scholar] [CrossRef]
- Enoki, T.; Yoshida, Y.; Lally, J.; Hatta, H.; Bonen, A. Testosterone increases lactate transport, monocarboxylate transporter (MCT) 1 and MCT4 in rat skeletal muscle. J. Physiol. 2006, 577, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Daly, W.; Seegers, C.A.; Rubin, D.A.; Dobridge, J.D.; Hackney, A.C. Relationship between stress hormones and testosterone with prolonged endurance exercise. Eur. J. Appl. Physiol. 2005, 93, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Hackney, A.C. Effects of endurance exercise on the reproductive system of men: The “exercise-hypogonadal male condition”. J. Endocrinol. Investig. 2008, 31, 932–938. [Google Scholar] [CrossRef]
- Cumming, D.C.; Quigley, M.E.; Yen, S.S.C. Acute suppression of circulating testosterone levels by cortisol in men. J. Clin. Endocrinol. Metab. 1983, 57, 671–673. [Google Scholar] [CrossRef]
- Vervoorn, C.; Quist, A.M.; Vermulst, L.J.M.; Erich, W.B.M.; De Vries, W.R.; Thijssen, J.H.H. The behaviour of the plasma free testosterone/cortisol ratio during a season of elite rowing training. Int. J. Sports Med. 1991, 12, 257–263. [Google Scholar] [CrossRef]
- Hackney, A.C.; Szczepanowska, E.; Viru, A.M. Basal testicular testosterone production in endurance-trained men is suppressed. Eur. J. Appl. Physiol. 2003, 89, 198–201. [Google Scholar] [CrossRef]
- Raju, G.A.R.; Chavan, R.; Deenadayal, M.; Gunasheela, D.; Gutgutia, R.; Haripriya, G.; Govindarajan, M.; Patel, N.H.; Patki, A.S. Luteinizing hormone and follicle stimulating hormone synergy: A review of role in controlled ovarian hyper-stimulation. J. Hum. Reprod. Sci. 2013, 6, 227. [Google Scholar] [CrossRef] [PubMed]
- Millar, R.P.; Lu, Z.-L.; Pawson, A.J.; Flanagan, C.A.; Morgan, K.; Maudsley, S.R. Gonadotropin-releasing hormone receptors. Endocr. Rev. 2004, 25, 235–275. [Google Scholar] [CrossRef]
- An, E.; Wilson, A. Exercise and gonadal function. Hum. Reprod. 1993, 8, 1747–1761. [Google Scholar]
- Elias, A.N.; Wilson, A.F.; Pandian, M.R.; Chune, G.; Utsumi, A.; Kayaleh, R.; Stone, S.C. Corticotropin releasing hormone and gonadotropin secretion in physically active males after acute exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1991, 62, 171–174. [Google Scholar] [CrossRef]
- Lehmann, M.; Gastmann, U.; Petersen, K.G.; Bachl, N.; Seidel, A.; Khalaf, A.N.; Fischer, S.; Keul, J. Training-overtraining: Performance, and hormone levels, after a defined increase in training volume versus intensity in experienced middle-and long-distance runners. Br. J. Sports Med. 1992, 26, 233–242. [Google Scholar] [CrossRef] [PubMed]
- De Luccia, T. Use of the testosterone/cortisol ratio variable in sports. Open Sports Sci. J. 2016, 9, 104–113. [Google Scholar] [CrossRef]
- Meeusen, R.; Duclos, M.; Foster, C.; Fry, A.; Gleeson, M.; Nieman, D.; Raglin, J.; Rietjens, G.; Steinacker, J.; Urhausen, A. Prevention, diagnosis, and treatment of the overtraining syndrome: Joint consensus statement of the European College of Sport Science and the American College of Sports Medicine. Med. Sci. Sports Exerc. 2013, 45, 186. [Google Scholar] [CrossRef]
- Comitato, R.; Saba, A.; Turrini, A.; Arganini, C.; Virgili, F. Sex hormones and macronutrient metabolism. Crit. Rev. Food Sci. Nutr. 2015, 55, 227–241. [Google Scholar] [CrossRef]
- Horton, T.J.; Grunwald, G.K.; Lavely, J.; Donahoo, W.T. Glucose kinetics differ between women and men, during and after exercise. J. Appl. Physiol. 2006, 100, 1883–1894. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; He, J.; Watkins, S.; Kelley, D.E. Skeletal muscle lipid content and insulin resistance: Evidence for a paradox in endurance-trained athletes. J. Clin. Endocrinol. Metab. 2001, 86, 5755–5761. [Google Scholar] [CrossRef] [PubMed]
- Popovic, B.; Popovic, D.; Macut, D.; Antic, I.B.; Isailovic, T.; Ognjanovic, S.; Bogavac, T.; Kovacevic, V.E.; Ilic, D.; Petrovic, M. Acute Response to Endurance Exercise Stress: Focus on Catabolic/Anabolic Interplay Between Cortisol, Testosterone, and Sex Hormone Binding Globulin in Professional Athletes. J. Med. Biochem. 2019, 38, 6–12. [Google Scholar] [PubMed]
- Wahl, P.; Mathes, S.; Köhler, K.; Achtzehn, S.; Bloch, W.; Mester, J. Acute metabolic, hormonal, and psychological responses to different endurance training protocols. Horm. Metab. Res. 2013, 45, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Yassin, A.A.; Doros, G. Testosterone therapy in hypogonadal men results in sustained and clinically meaningful weight loss. Clin. Obes. 2013, 3, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Moreiras, O. Tablas de Composición de Alimentos, 16th ed.; Pirámide: Madrid, Spain, 2013. [Google Scholar]
- Jackson, A.S.; Pollock, M.L. Practical Assessment of Body Composition. Phys. Sportsmed. 1985, 13, 76–90. [Google Scholar] [CrossRef]
- Stewart, A.; Marfell-Jones, M.; Olds, T.; de Ridder, H. International Standards for Anthropometric Assessment; ISAK: Lower Hutt, New Zealand, 2011. [Google Scholar]
- Skinner, J.S.; Mclellan, T.H.; McLellan, T.H. The Transition from Aerobic to Anaerobic Metabolism. Res. Q. Exerc. Sport 1980, 51, 234–248. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge Academic: New York, NY, USA, 1988. [Google Scholar]
- Rodriguez, N.R.; DiMarco, N.M.; Langley, S.; American Dietetic Association; Dietitians of Canada; American College of Sports Medicine: Nutrition and Athletic Performance. Position of the American Dietetic Association, Dietitians of Canada, and the American College of Sports Medicine: Nutrition and Athletic Performance. J. Am. Diet. Assoc. 2009, 109, 509–527. [Google Scholar] [CrossRef]
- Dipla, K.; Kraemer, R.R.; Constantini, N.W.; Hackney, A.C. Relative energy deficiency in sports (RED-S): Elucidation of endocrine changes affecting the health of males and females. Hormones 2020. [Google Scholar] [CrossRef]
- Basset, F.A.; Chouinard, R.; Boulay, M.R. Training profile counts for time-to-exhaustion performance. Can. J. Appl. Physiol. 2003, 28, 654–666. [Google Scholar] [CrossRef]
- Berg, K. Endurance training and performance in runners—Research limitations and unanswered questions. Sports Med. 2003, 33, 59–73. [Google Scholar] [CrossRef]
- Rabadán, M.; Díaz, V.; Calderón, F.J.; Benito, P.J.; Peinado, A.B.; Maffulli, N. Physiological determinants of speciality of elite middle- and long-distance runners. J. Sports Sci. 2011, 29, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Hackney, A.; Sinning, W.E.; Bruot, B.C. Reproductive hormonal profiles of endurance-trained and untrained males. Med. Sci. Sports Exerc. 1988, 20, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Volek, J.S.; Ratamess, N.A.; Rubin, M.R.; Gomez, A.L.; French, D.N.; McGuigan, M.M.; Scheett, T.P.; Sharman, M.J.; Häkkinen, K.; Kraemer, W.J. The effects of creatine supplementation on muscular performance and body composition responses to short-term resistance training overreaching. Eur. J. Appl. Physiol. 2004, 91, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Mäestu, J.; Jürimäe, J.; Jürimäe, T. Hormonal reactions during heavy training stress and following tapering in highly trained male rowers. Horm. Metab. Res. 2003, 35, 109–113. [Google Scholar] [CrossRef]
- Purge, P.; Jürimäe, J.; Jürimäe, T. Hormonal and psychological adaptation in elite male rowers during prolonged training. J. Sports Sci. 2006, 24, 1075–1082. [Google Scholar] [CrossRef]
- Urhausen, A.; Gabriel, H.; Kindermann, W. Blood hormones as markers of training stress and overtraining. Sports Med. 1995, 20, 251–276. [Google Scholar] [CrossRef]
- Petibois, C.; Cazorla, G.; Deleris, G. The biological and metabolic adaptations to 12 months training in elite rowers. Int. J. Sports Med. 2003, 24, 36–42. [Google Scholar] [CrossRef]
- Elliott-Sale, K.J.; Tenforde, A.S.; Parziale, A.L.; Holtzman, B.; Ackerman, K.E. Endocrine effects of relative energy deficiency in sport. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 335–349. [Google Scholar] [CrossRef]
- Lane, A.R.; Anderson, T.; Hackney, A.C. Relationship Between Cortisol and Free Testosterone in Response to Exhaustive Endurance Exercise. Age 2015, 22, 19–28. [Google Scholar]
- Shahani, S.; Braga-Basaria, M.; Maggio, M.; Basaria, S. Androgens and erythropoiesis: Past and present. J. Endocrinol. Investig. 2009, 32, 704–716. [Google Scholar] [CrossRef]
- Hackney, A.C.; Lane, A.R. Low testosterone in male endurance-trained distance runners: Impact of years in training. Hormones 2018, 17, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Krsmanovic, L.Z.; Hu, L.; Leung, P.-K.; Feng, H.; Catt, K.J. The hypothalamic GnRH pulse generator: Multiple regulatory mechanisms. Trends Endocrinol. Metab. 2009, 20, 402–408. [Google Scholar] [CrossRef] [PubMed]
- MacConnie, S.E.; Barkan, A.; Lampman, R.M.; Schork, M.A.; Beitins, I.Z. Decreased hypothalamic gonadotropin-releasing hormone secretion in male marathon runners. N. Engl. J. Med. 1986, 315, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.R.; Kraemer, W.J.; Saenz, C.; Schill, K.E.; Focht, B.C.; Volek, J.S.; Maresh, C.M. The presence of symptoms of testosterone deficiency in the exercise-hypogonadal male condition and the role of nutrition. Eur. J. Appl. Physiol. 2017, 117, 1349–1357. [Google Scholar] [CrossRef]
- De Souza, M.J.; Arce, J.C.; Pescatello, L.S.; Scherzer, H.S.; Luciano, A.A. Gonadal hormones and semen quality in male runners. Int. J. Sports Med. 1994, 15, 383–391. [Google Scholar] [CrossRef]
- Flynn, M.G.; Pizza, F.X.; Brolinson, P.G. Hormonal responses to excessive training: Influence of cross training. Int. J. Sports Med. 1997, 18, 191–196. [Google Scholar] [CrossRef]
- Hackney, A.C. Hypogonadism in Exercising Males: Dysfunction or Adaptive-Regulatory Adjustment? Front. Endocrinol. (Lausanne) 2020, 11, 1–16. [Google Scholar] [CrossRef]
- De Souza, M.J.; Koltun, K.J.; Williams, N.I. The Role of Energy Availability in Reproductive Function in the Female Athlete Triad and Extension of its Effects to Men: An Initial Working Model of a Similar Syndrome in Male Athletes. Sports Med. 2019, 49, 125–137. [Google Scholar] [CrossRef]
- Bellastella, G.; Pane, E.; Iorio, S.; De Bellis, A.; Sinisi, A.A. Seasonal variations of plasma gonadotropin, prolactin, and testosterone levels in primary and secondary hypogonadism: Evidence for an independent testicular role. J. Endocrinol. Investig. 2013, 36, 339–342. [Google Scholar]
- Chen, C.; Zhai, H.; Cheng, J.; Weng, P.; Chen, Y.; Li, Q.; Wang, C.; Xia, F.; Wang, N.; Lu, Y. Causal Link between Vitamin D and Total Testosterone in Men: A Mendelian Randomization Analysis. J. Clin. Endocrinol. Metab. 2019, 104, 3148–3156. [Google Scholar] [CrossRef]
- Lombardi, G.; Vitale, J.A.; Logoluso, S.; Logoluso, G.; Cocco, N.; Cocco, G.; Cocco, A.; Banfi, G. Circannual rhythm of plasmatic vitamin D levels and the association with markers of psychophysical stress in a cohort of Italian professional soccer players. Chronobiol. Int. 2017, 34, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.K.; Hoermann, R.; Grossmann, M. Reversible male hypogonadotropic hypogonadism due to energy deficit. Clin. Endocrinol. (Oxf.) 2019, 91, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Cumming, D.C.; Wheeler, G.D.; McColl, E.M. The effects of exercise on reproductive function in men. Sports Med. 1989, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Hackney, A.C.; Viru, A. Twenty-four-hour cortisol response to multiple daily exercise sessions of moderate and high intensity. Clin. Physiol. 1999, 19, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Flynn, M.G.; Pizza, F.X.; Boone, J.B.; Andres, F.F.; Michaud, T.A.; Rodriguez-Zayas, J.R. Indices of training stress during competitive running and swimming seasons. Int. J. Sports Med. 1994, 15, 21–26. [Google Scholar] [CrossRef]
- Gillespie, C.A.; Edgerton, V.R. The role of testosterone in exercise-induced glycogen supercompensation. Horm. Metab. Res. 1970, 2, 364–366. [Google Scholar] [CrossRef]
- Kuoppasalmi, K.; Adlercreutz, H. Interaction between catabolic and anabolic steroid hormones in muscular exercise. Exerc. Endocrinol. 1985, 65–98. [Google Scholar] [CrossRef]
- Smith, R.P.; Coward, R.M.; Kovac, J.R.; Lipshultz, L.I. The evidence for seasonal variations of testosterone in men. Maturitas 2013, 74, 208–212. [Google Scholar] [CrossRef]
- Jürimäe, J.; Purge, P.; Jürimäe, T. Adiponectin and stress hormone responses to maximal sculling after volume-extended training season in elite rowers. Metabolism 2006, 55, 13–19. [Google Scholar] [CrossRef]
- Hartley, L.H.; Mason, J.W.; Hogan, R.P.; Jones, L.G.; Kotchen, T.A.; Mougey, E.H.; Wherry, F.E.; Pennington, L.L.; Ricketts, P.T. Multiple hormonal responses to prolonged exercise in relation to physical training. J. Appl. Physiol. 1972, 33, 607–610. [Google Scholar] [CrossRef]
- Borghouts, L.B.; Keizer, H.A. Exercise and insulin sensitivity: A review. Int. J. Sports Med. 2000, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Training Load | Initial | 3 Months | 6 Months | 9 Months |
|---|---|---|---|---|
| Total (km/week) | 85.71 ± 13.62 | 105.9 ± 16.85 | 93.33 ± 14.34 | 74.76 ± 14.09 |
| >VT2 (km/week) | 4.29 ± 0.68 | 12.71 ± 2.02 | 18.67 ± 2.86 | 16.45 ± 3.10 |
| <VT2 (km/week) | 81.43 ± 12.94 | 93.24 ± 14.83 | 74.67 ± 11.47 | 58.31 ± 10.99 |
| Parameters | Initial | 3 Months | 6 Months | 9 Months | ηp2 |
|---|---|---|---|---|---|
| VO2 max (mL/min/kg) | 68.12 ± 4.63 | 67.53 ± 9.54 | 68.55 ± 6.97 | 68.60 ± 7.36 | 0.05 |
| VT2 (% VO2 max.) | 91.02 ± 2.43 | 92.43 ± 3.59 | 91.02 ± 3.08 | 90.96 ± 2.07 | 0.06 |
| RER | 1.05 ± 0.03 | 1.05 ± 0.04 | 1.05 ± 0.05 | 1.04 ± 0.04 | 0.01 |
| HR maximum | 190.4 ± 9.48 | 193.1 ± 7.80 | 193.5 ± 9.06 | 193.8 ± 7.19 | 0.04 |
| Weight (kg) | 65.50 ± 7.30 | 65.45 ± 7.36 | 64.67 ± 7.03 * | 64.80 ± 7.34 * | 0.07 |
| Fat mass (%) | 8.18 ± 1.04 | 8.23 ± 1.04 | 8.19 ± 1.29 | 8.21 ± 1.07 | 0.04 |
| Fat-free mass (kg) | 60.15 ± 6.70 | 60.07 ± 6.75 | 59.38 ± 6.45 | 59.48 ± 6.73 | 0.02 |
| ∑6 skinfold (mm) | 45.65 ± 10.88 | 46.49 ± 10.69 | 44.92 ± 8.16 | 45.37 ± 9.11 | 0.03 |
| Parameters | Initial | 3 Months | 6 Months | 9 Months |
|---|---|---|---|---|
| Energy (kcal/d) | 2855.21 ± 511.32 | 2515.48 ± 427.18 | 2902.37 ± 522.62 | 3108.78 ± 770.12 |
| EA (kcal/kg/FFM/d) | 43.58 ± 4.32 | 41.87 ± 3.15 | 48.88 ± 5.63 | 52.26 ± 4.87 |
| HC (g/kg/d) | 5.26 ± 1.21 | 5.18 ± 1.14 | 6.25 ± 1.38 | 6.13 ± 1.50 |
| Proteins (g/kg/d) | 1.73 ± 0.79 | 1.69 ± 0.35 | 1.85 ± 0.53 | 1.89 ± 0.63 |
| Lipids (g/kg/d) | 1.78 ± 0.40 | 1.63 ± 0.28 | 1.58 ± 0.52 | 1.72 ± 0.74 |
| Parameters | Initial | 3 Months | 6 Months | 9 Months | ηp2 |
|---|---|---|---|---|---|
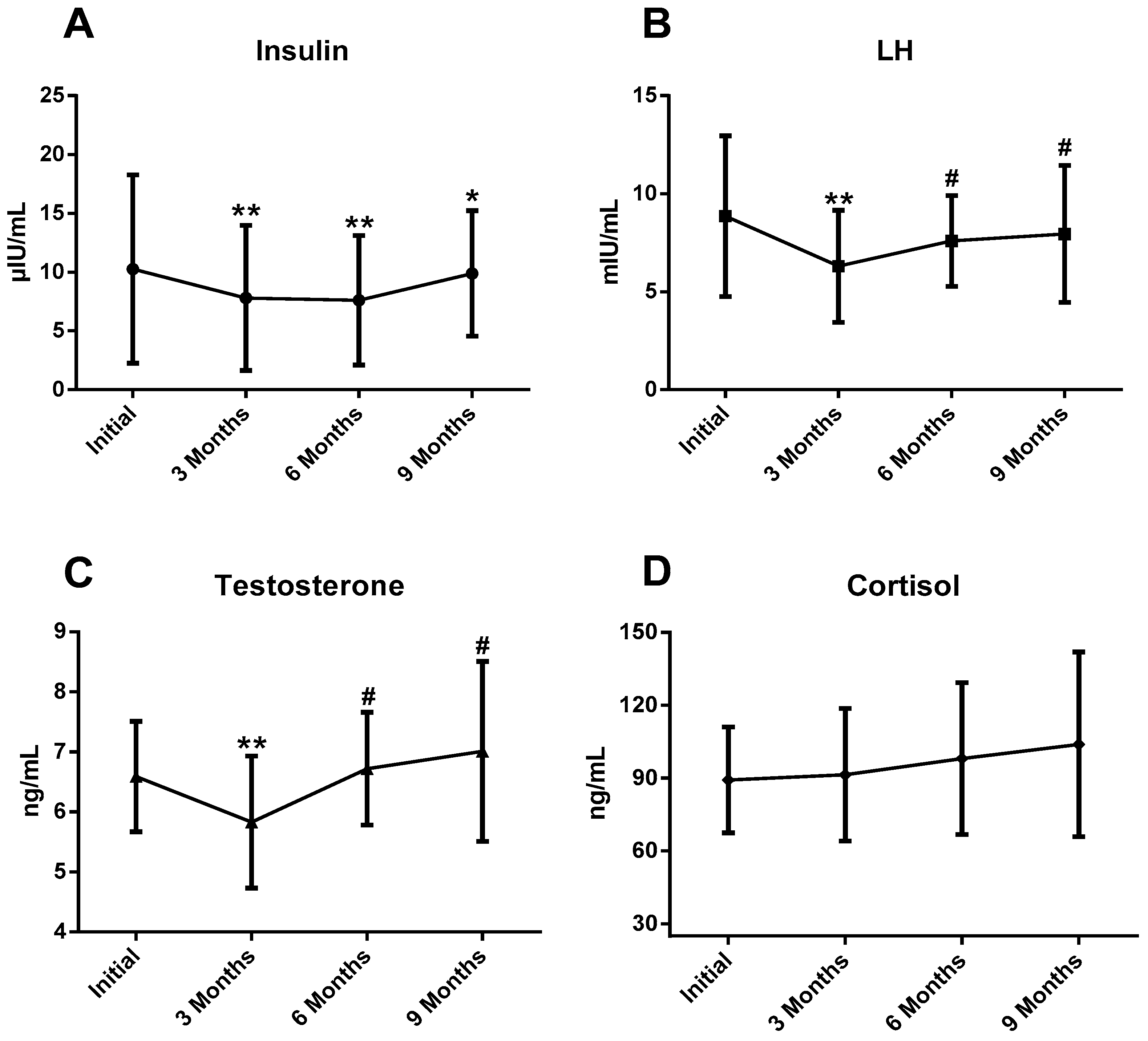
| Insulin (μIU/mL) | 10.25 ± 7.99 | 7.81 ± 6.15 ** | 7.62 ± 5.50 ** | 9.89 ± 5.32 * | 0.51 |
| LH (mIU/mL) | 8.85 ± 4.10 | 6.30 ± 2.86 ** | 7.59 ± 2.32 # | 7.95 ± 3.49 # | 0.29 |
| Testosterone (ng/mL) | 6.59 ± 0.92 | 5.83 ± 1.10 ** | 6.72 ± 0.94 # | 7.01 ± 1.50 # | 0.32 |
| Cortisol (ng/mL) | 89.26 ± 21.85 | 91.41 ± 27.32 | 98.06 ± 31.28 | 103.9 ± 38.04 | 0.05 |
| T/C | 0.07 ± 0.01 | 0.06 ± 0.02 | 0.07 ± 0.02 | 0.07 ± 0.02 | 0.03 |
| Hormones | β (95% CI) | SE | r | R2 | p |
|---|---|---|---|---|---|
| Insulin | −0.577 (−0.984/0.030) | 0.255 | 0.363 | 0.142 | 0.017 |
| LH | −1.105 (−2.769/−0.440) | 0.585 | 0.113 | 0.025 | 0.302 |
| Testosterone | −2.145 (−5.247/−3.543) | 1.109 | 0.131 | 0.028 | 0.213 |
| Cortisol | 0.310 (−0.124/0.544) | 0.167 | 0.202 | 0.075 | 0.043 |
| Hormones | β (95% CI) | SE | r | R2 | p |
|---|---|---|---|---|---|
| Insulin | −0.442 (−0.579/−0.306) | 0.069 | 0.580 | 0.336 | 0.000 |
| LH | −0.320 (−0.707/0.068) | 0.195 | 0.178 | 0.032 | 0.104 |
| Testosterone | 0.276 (−0.812/1.364) | 3.638 | 0.056 | 0.003 | 0.615 |
| Cortisol | 0.238 (−0.004/0.081) | 2.150 | 0.193 | 0.037 | 0.079 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, J.; Toro, V.; Barrientos, G.; Bartolomé, I.; Muñoz, D.; Maynar, M. Hormonal Changes in High-Level Aerobic Male Athletes during a Sports Season. Int. J. Environ. Res. Public Health 2020, 17, 5833. https://doi.org/10.3390/ijerph17165833
Alves J, Toro V, Barrientos G, Bartolomé I, Muñoz D, Maynar M. Hormonal Changes in High-Level Aerobic Male Athletes during a Sports Season. International Journal of Environmental Research and Public Health. 2020; 17(16):5833. https://doi.org/10.3390/ijerph17165833
Chicago/Turabian StyleAlves, Javier, Víctor Toro, Gema Barrientos, Ignacio Bartolomé, Diego Muñoz, and Marcos Maynar. 2020. "Hormonal Changes in High-Level Aerobic Male Athletes during a Sports Season" International Journal of Environmental Research and Public Health 17, no. 16: 5833. https://doi.org/10.3390/ijerph17165833
APA StyleAlves, J., Toro, V., Barrientos, G., Bartolomé, I., Muñoz, D., & Maynar, M. (2020). Hormonal Changes in High-Level Aerobic Male Athletes during a Sports Season. International Journal of Environmental Research and Public Health, 17(16), 5833. https://doi.org/10.3390/ijerph17165833