Evaluation of Bioaerosol Bacterial Components of a Wastewater Treatment Plant Through an Integrate Approach and In Vivo Assessment


 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Experimental
2.3. Sample Preparation for Chemical Analyses
2.4. Isolation of Cultivable Bacteria
2.5. DNA Extraction and Bacterial Isolates Identification
2.6. Metagenomic DNA Extraction and Analysis
2.7. Determination of Antibiotic Susceptibility
2.8. Caenorhabditis Elegans Growth and Maintenance
2.9. Nematodes Infection
2.10. Life Span Analysis
2.11. Nematodes Brood Size Assays
2.12. Measurement of Nematodes Body Length
2.13. Body Bend Analysis
2.14. Pharyngeal Pumping Assay
2.15. Statistical Analysis
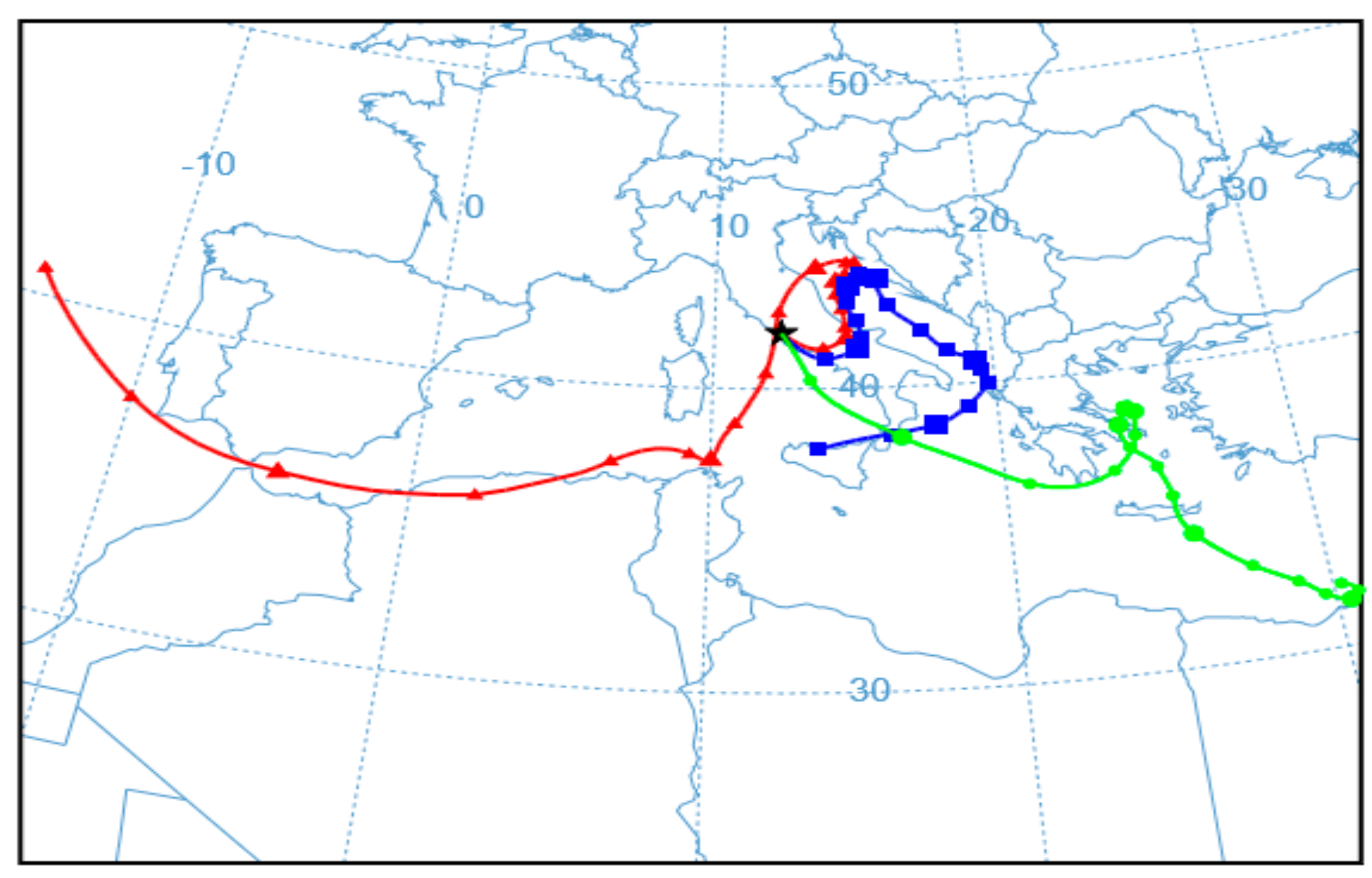
3. Results and Discussion
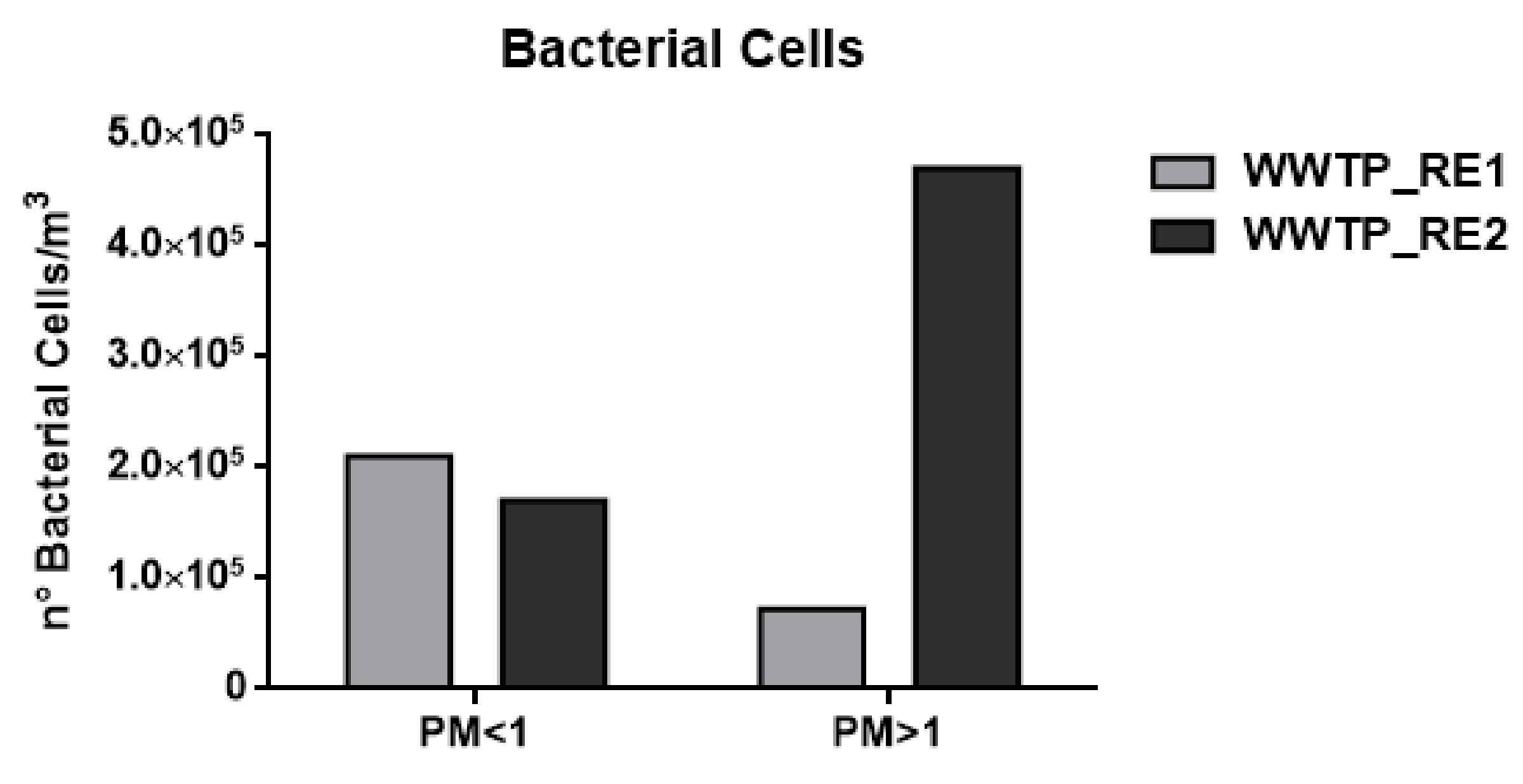
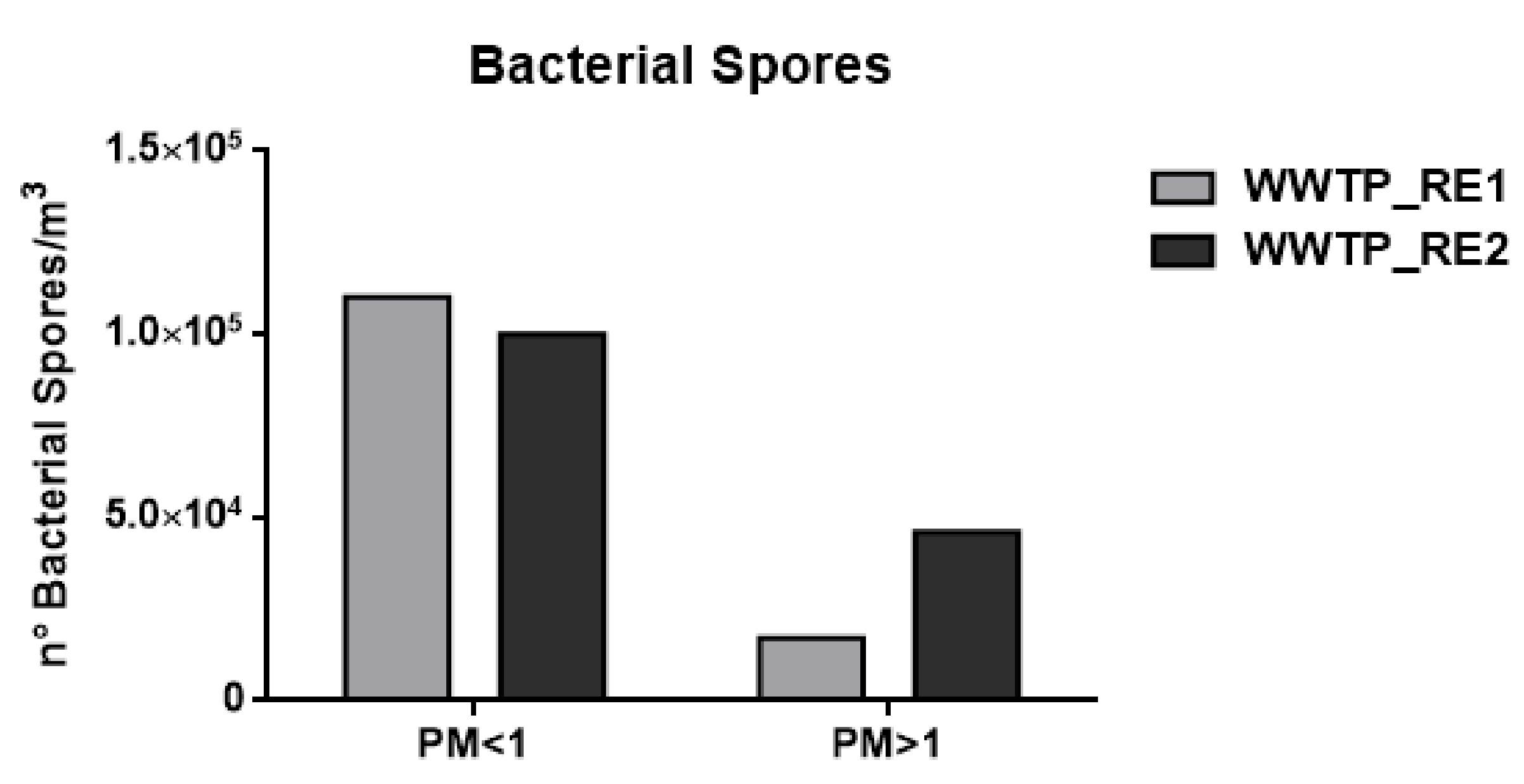
3.1. Bacterial Cells and Fungal and Bacterial Spores by Chemical Approach
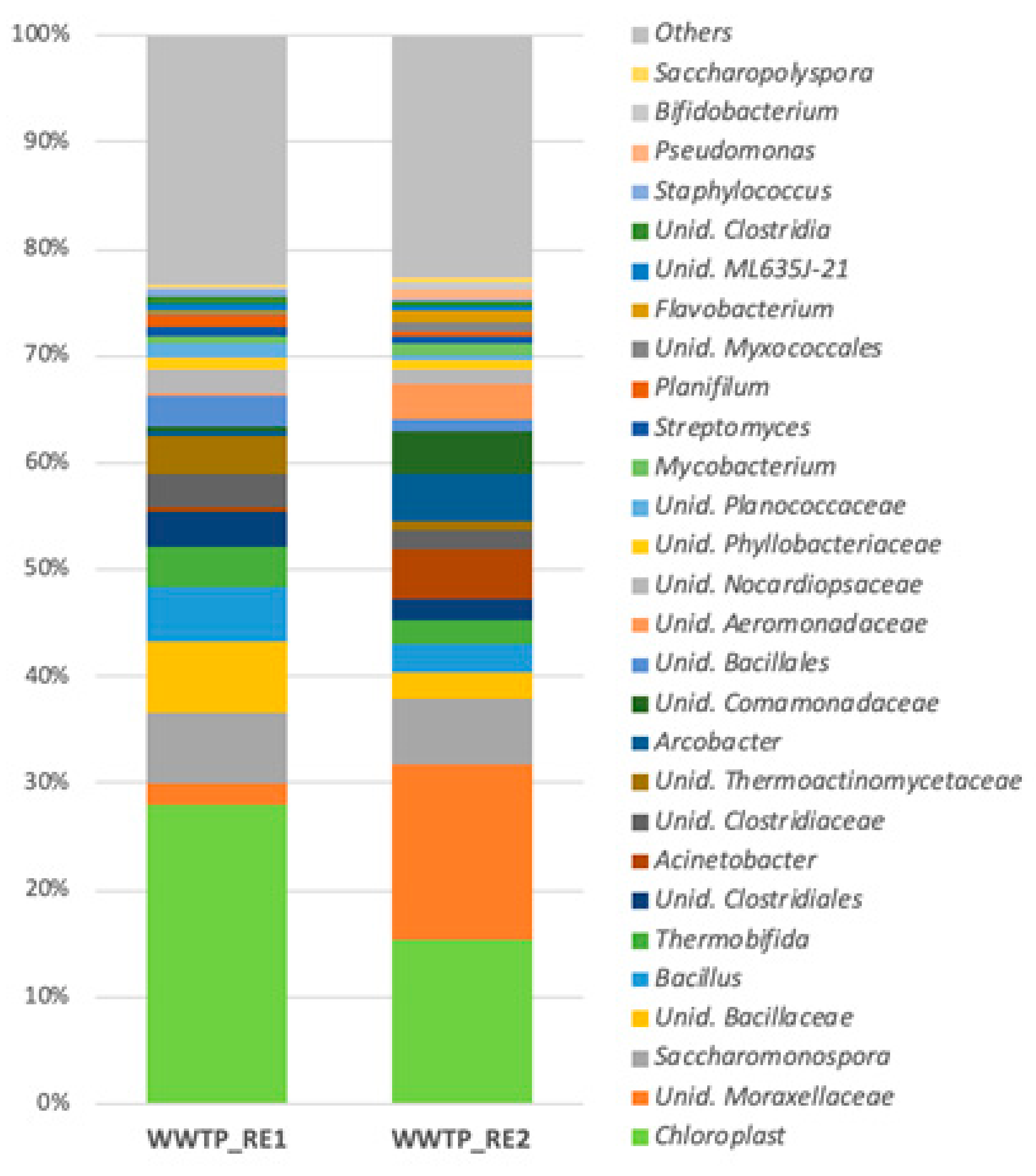
3.2. Bacterial Bioaerosol Diversity and Taxonomic Composition
3.3. Isolation of Viable Bacteria
3.4. Identification of Cultivable Bacteria by 16S rDNA Sequencing and Antibiotic Susceptibility Test
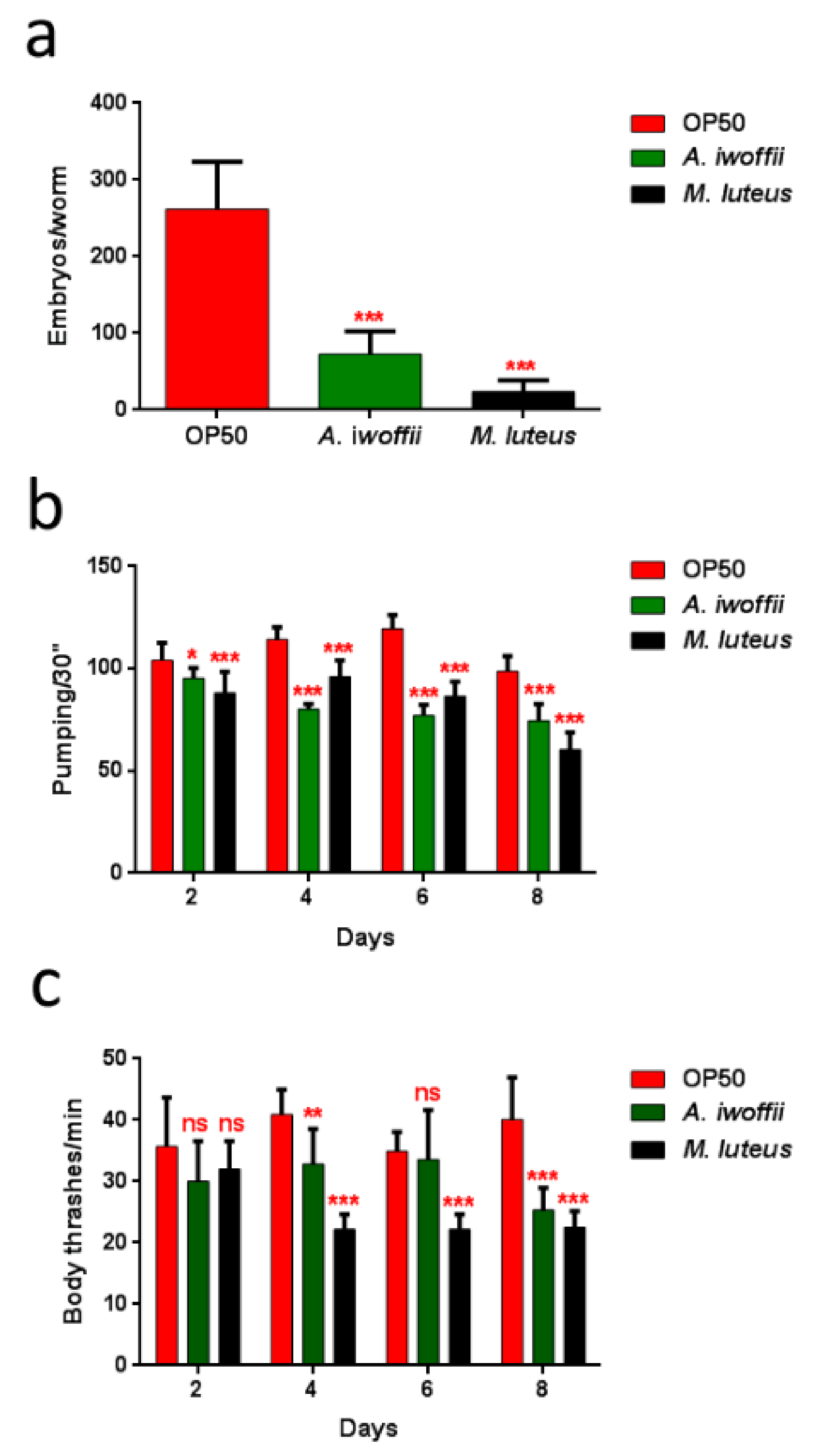
3.5. In Vivo Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fröhlich-Nowoisky, J.; Kampf, C.J.; Weber, B.; Huffman, J.A.; Pöhlker, C.; Andreae, M.O.; Lang-Yona, N.; Burrows, S.M.; Gunthe, S.S.; Elbert, W.; et al. Bioaerosols in the Earth system: Climate, health, and ecosystem interactions. Atmos. Res. 2016, 182, 346–376. [Google Scholar] [CrossRef]
- Bonazza, A.; De Nuntiis, P.; Mandrioli, P.; Sabbioni, C. Aerosol Impact on Cultural Heritage: Deterioration Processes and Strategies for Preventive Conservation. In Atmospheric Aerosols: Life Cycles and Effects on Air Quality and Climate; Tomasi, C., Fuzzi, S., Kokhanovsky, A., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2017; pp. 645–670. [Google Scholar] [CrossRef]
- Eduard, W.; Heederik, D.; Duchaine, C.; Green, B.J. Bioaerosol exposure assessment in the workplace: The past, present and recent advances. J. Environ. Monit. 2012, 14, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.D.; Ricke, S.C. Bioaerosols from municipal and animal wastes: Background and contemporary issues. Can. J. Microbiol. 2002, 48, 681–696. [Google Scholar] [CrossRef] [PubMed]
- Vitezova, M.; Mach, P.; Vitez, T.; Losak, T. Development of microbial community in the course of composting of garden waste. Acta Univ. Agric. Silvic. Mendel. Brun. 2012, 60, 225–232. [Google Scholar] [CrossRef]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef]
- Brągoszewska, E.; Pastuszka, J.S. Influence of meteorological factors on the level and characteristics of culturable bacteria in the air in Gliwice, Upper Silesia (Poland). Aerobiologia 2018, 34, 241–255. [Google Scholar] [CrossRef]
- Brandi, G.; Sisti, M.; Amagliani, G. Evaluation of the environmental impact of microbial aerosols generated by wastewater treatment plants utilizing different aeration systems. J. Appl. Microbiol. 2000, 88, 845–852. [Google Scholar] [CrossRef]
- Carducci, A.; Tozzi, E.; Rubulotta, E.; Casini, B.; Cantiani, L.; Rovini, E.; Muscillo, M.; Pacini, R. Assessing airborne biological hazard from urban wastewater treatment. Water Res. 2000, 34, 1173–1178. [Google Scholar] [CrossRef]
- Sanchez-Monedero, M.A.; Stentiford, E.I.; Mondini, C. Biofiltration at composting facilities: Effectiveness for bioaerosol control. Environ. Sci. Technol. 2003, 37, 4299–4303. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Xue, S.; Han, Y.; Yang, K. Characteristics and formation mechanism of intestinal bacteria particles emitted from aerated wastewater treatment tanks. Water Res. 2019, 163, 114862. [Google Scholar] [CrossRef]
- Caicedo, C.; Beutel, S.; Scheper, T.; Rosenwinkel, K.H.; Nogueira, R. Occurrence of Legionella in wastewater treatment plants linked to wastewater characteristics. Environ. Sci. Pollut. Res. 2016, 23, 16873–16881. [Google Scholar] [CrossRef]
- Higgins, P.G.; Hrenovic, J.; Seifert, H.; Dekic, S. Characterization of Acinetobacter baumannii from water and sludge line of secondary wastewater treatment plant. Water Res. 2018, 140, 261–267. [Google Scholar] [CrossRef]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef]
- Guo, J.; Li, J.; Chen, H.; Bond, P.L.; Yuan, Z. Metagenomic analysis reveals wastewater treatment plants as hotspots of antibiotic resistance genes and mobile genetic elements. Water Res. 2017, 123, 468–478. [Google Scholar] [CrossRef]
- Gaviria-Figueroa, A.; Preisner, E.C.; Hoque, S.; Feigley, C.E.; Norman, R.S. Emission and dispersal of antibiotic resistance genes through bioaerosols generated during the treatment of municipal sewage. Sci. Total Environ. 2019, 686, 402–412. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef]
- Wolf, R.; El-Haddad, I.; Slowik, J.G.; Dällenbach, K.; Bruns, E.; Vasilescu, J.; Baltensperger, U.; Prévôt, A.S.H. Contribution of bacteria-like particles to PM 2.5 aerosol in urban and rural environments. Atmos. Environ. 2017, 160, 97–106. [Google Scholar] [CrossRef]
- Zukiewicz-Sobczak, W.A. The role of fungi in allergic diseases. Postep. Dermatol. Alergol. 2013, 30, 42–45. [Google Scholar] [CrossRef]
- Xu, Z.; Yao, M. Analysis of culturable bacterial and fungal aerosol diversity obtained using different samplers and culturing methods. Aerosol Sci. Technol. 2011, 45, 1143–1153. [Google Scholar] [CrossRef]
- Simon, X.; Duquenne, P. Assessment of workers’ exposure to bioaerosols in a French cheese factory. Ann. Occup. Hyg. 2014, 58, 677–692. [Google Scholar] [CrossRef][Green Version]
- Shaffer, B.T.; Lighthart, B. Survey of culturable airborne bacteria at four diverse locations in Oregon: Urban, rural, forest, and coastal. FEMS Microbiol. Ecol. 1997, 34, 167–177. [Google Scholar] [CrossRef]
- Chang, C.W.; Chung, H.; Huang, C.F.; Su, H.J. Exposure of Workers to Airborne Microorganisms in Open-air Swine Houses. J. Appl. Environ. Microbiol. 2001, 67, 155–161. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic Identification and in Situ Detection of Individual Microbial Cells without Cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar]
- Pace, N.R. A molecular view of microbial diversity and the biosphere. Science 1997, 276, 734–740. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar]
- Albrecht, A.; Witzenberger, R.; Bernzen, U.; Jackel, U. Detection of airborne microbes in a composting facility by cultivation based and cultivation independent methods. Ann. Agric. Environ. Med. 2007, 14, 81–85. [Google Scholar]
- Alvarez, A.J.; Buttner, M.P.; Toranzos, G.A.; Dvorsky, E.A.; Toro, A.; Heikes, T.B.; Mertikas-Pifer, L.E.; Stetzenbach, L.D. Use of solid-phase PCR for enhanced detection of airborne microorganisms. Appl. Environ. Microbiol. 1994, 60, 374–376. [Google Scholar]
- Han, Y.; Li, L.; Liu, J. Characterization of the airborne bacteria community at different distances from the rotating brushes in a wastewater treatment plant by 16S rRNA gene clone libraries. J. Environ. Sci. 2013, 25, 5–15. [Google Scholar] [CrossRef]
- Degois, J.; Clerc, F.; Simon, X.; Bontemps, C.; Leblond, P.; Duquenne, P. First metagenomic survey of the microbial diversity in bioaerosols emitted in waste sorting plants. Ann. Work Expo. Health 2017, 61, 1076–1086. [Google Scholar] [CrossRef]
- Hall, R.J.; Leblanc-Maridor, M.; Wang, J.; Ren, X.; Moore, N.E.; Brooks, C.R.; Peacey, M.; Douwes, J.; McLean, D.J. Metagenomic detection of viruses in aerosol samples from workers in animal slaughterhouses. PLoS ONE 2013, 8, e72226. [Google Scholar] [CrossRef]
- Olsen, R.H.; Bakken, L.R. Viability of Soil Bacteria: Optimization of Plate-Counting Technique and Comparison between Total Counts and Plate Counts within Different Size Groups. Microb. Ecol. 1987, 13, 59–74. [Google Scholar] [CrossRef]
- Jannasch, H.W.; Jones, G.E. Bacterial populations in seawater as determined by different methods of enumeration. Limnol. Oceanogr. 1959, 4, 128–139. [Google Scholar] [CrossRef]
- Buiarelli, F.; Gallo, V.; Di Filippo, P.; Pomata, D.; Riccardi, C. Development of a method for the analysis of underivatized amino acids by liquid chromatography/tandem mass spectrometry: Application on Standard Reference Material 1649a (urban dust). Talanta 2013, 115, 966–972. [Google Scholar] [CrossRef]
- Buiarelli, F.; Canepari, S.; Di Filippo, P.; Perrino, C.; Pomata, D.; Riccardi, C.; Speziale, R. Extraction and analysis of fungal spore biomarkers in atmospheric bioaerosol by HPLC–MS–MS and GC–MS. Talanta 2013, 105, 142–151. [Google Scholar] [CrossRef]
- Di Filippo, P.; Pomata, D.; Riccardi, C.; Buiarelli, F.; Gallo, V.; Quaranta, A. Free and combined amino acids in size-segregated atmospheric aerosol samples. Atmos. Environ. 2014, 98, 179–189. [Google Scholar] [CrossRef]
- Di Filippo, P.; Pomata, D.; Riccardi, C.; Buiarelli, F.; Uccelletti, D.; Zanni, E. Muramic and dipicolinic acids in atmospheric particulate matter as biomarkers of bacteria and bacterial spores. Anal. Bioanal. Chem. 2017, 409, 1657–1666. [Google Scholar] [CrossRef]
- Lau, A.P.S.; Lee, A.K.Y.; Chan, C.K.; Fang, M. Ergosterol as a biomarker for the quantification of the fungal biomass in atmospheric aerosols. Atmos. Environ. 2006, 40, 249–259. [Google Scholar] [CrossRef]
- Kumar, A.; Baruah, A.; Tomioka, M.; Iino, Y.; Kalita, M.C.; Khan, M. Caenorhabditis elegans: A model to understand host-microbe interactions. Cell. Mol. Life Sci. 2019, 1–21. [Google Scholar] [CrossRef]
- Brągoszewska, E.; Mainka, A.; Pastuszka, J. Concentration and size distribution of culturable bacteria in ambient air during spring and winter in Gliwice: A typical urban area. Atmosphere 2017, 8, 239. [Google Scholar] [CrossRef]
- Di Filippo, P.; Riccardi, C.; Pomata, D.; Buiarelli, F. Concentrations of PAHs, and nitro-and methyl-derivatives associated with a size-segregated urban aerosol. Atmos. Environ. 2010, 44, 2742–2749. [Google Scholar] [CrossRef]
- Di Filippo, P.; Pomata, D.; Riccardi, C.; Buiarelli, F.; Gallo, V. Oxygenated polycyclic aromatic hydrocarbons in size-segregated urban aerosol. J. Aerosol Sci. 2015, 87, 126–134. [Google Scholar] [CrossRef]
- Buiarelli, F.; Di Filippo, P.; Massimi, L.; Pomata, D.; Riccardi, C.; Simonetti, G.; Sonego, E. Ultrafine, fine and coarse airborne particle mass concentration in workplaces. Atmos. Pollut. Res. 2019, 10, 1685–1690. [Google Scholar] [CrossRef]
- Buiarelli, F.; Sonego, E.; Uccelletti, D.; Bruni, E.; Di Filippo, P.; Pomata, D.; Riccardi, C.; Perrino, C.; Marcovecchio, F.; Simonetti, G. Determination of the main bioaerosol components using chemical markers by liquid chromatography–tandem mass spectrometry. Microchem. J. 2019, 149, 103974. [Google Scholar] [CrossRef]
- Birnboim, H.C.; Doly, J. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 1979, 7, 1513–1523. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Federici, E.; Petroselli, C.; Montalbani, E.; Casagrande, C.; Ceci, E.; Moroni, B.; La Porta, G.; Castellini, S.; Selvaggi, R.; Sebastiani, B.; et al. Airborne bacteria and persistent organic pollutants associated with an intense Saharan dust event in the Central Mediterranean. Sci. Total Environ. 2018, 645, 401–410. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schifano, E.; Zinno, P.; Guantario, B.; Roselli, M.; Marcoccia, S.; Devirgiliis, C.; Uccelletti, D. The Foodborne Strain Lactobacillus fermentum MBC2 Triggers pept-1-Dependent Pro-Longevity Effects in Caenorhabditis elegans. Microorganisms 2019, 7, 45. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. elegans (11 February 2006). In WormBook; WormBook, The C. elegans Research Community: Minneapolis, MN, USA, 1999; Volume 2, pp. 51–67. [Google Scholar] [CrossRef]
- Nawa, M.; Matsuoka, M. The Method of the Body Bending Assay Using Caenorhabditis elegans. Bio-Protocol 2012, 2, e253. [Google Scholar] [CrossRef]
- Uccelletti, D.; Pascoli, A.; Farina, F.; Alberti, A.; Mancini, P.; Hirschberg, C.B.; Palleschi, C. APY-1, a novel Caenorhabditis elegans apyrase involved in unfolded protein response signalling and stress responses. Mol. Biol. Cell. 2008, 19, 1337–1345. [Google Scholar] [CrossRef]
- Pomata, D.; Di Filippo, P.; Riccardi, C.; Buiarelli, F.; Gallo, V. Determination of non-certified levoglucosan, sugar polyols and ergosterol in NIST Standard Reference Material 1649a. Atmos. Environ. 2014, 84, 332–338. [Google Scholar] [CrossRef]
- Di Filippo, P.; Pomata, D.; Riccardi, C.; Buiarelli, F.; Perrino, C. Fungal contribution to size-segregated aerosol measured through biomarkers. Atmos. Environ. 2013, 64, 132–140. [Google Scholar] [CrossRef]
- Elbert, W.; Taylor, P.E.; Andreae, M.O.; Pöschl, U. Contribution of fungi to primary biogenic aerosols in the atmosphere: Wet and dry discharged spores, carbohydrates, and inorganic ions. Atmos. Chem. Phys. 2007, 7, 4569–4588. [Google Scholar] [CrossRef]
- Kowalski, M.; Wolany, J.; Pastuszka, J.S.; Płaza, G.; Wlazło, A.; Ulfig, K.; Malina, A. Characteristics of airborne bacteria and fungi in some Polish wastewater treatment plants. Int. J. Environ. Sci. Technol. 2017, 14, 2181–2192. [Google Scholar] [CrossRef]
- Li, Y.; Lu, R.; Li, W.; Xie, Z.; Song, Y. Concentrations and size distributions of viable bioaerosols under various weather conditions in a typical semi-arid city of Northwest China. J. Aerosol Sci. 2017, 106, 83–92. [Google Scholar] [CrossRef]
- Reche, I.; Ortega-Retuerta, E.; Romera, O.; Pulido-Villena, E.; Morales-Baquero, R.; Casamayor, E.O. Effect of Saharan dust inputs on bacterial activity and community composition in Mediterranean lakes and reservoirs. Limnol. Oceanogr. 2009, 54, 869–879. [Google Scholar] [CrossRef]
- Stein, A.F.; Draxler, R.R.; Rolph, G.D.; Stunder, B.J.B.; Cohen, M.D.; Ngan, F. NOAA’s HYSPLIT atmospheric transport and dispersion modeling system. Bull. Am. Meteorol. Soc. 2015, 96, 2059–2077. [Google Scholar] [CrossRef]
- Bauer, H.; Kasper-Giebl, A.; Löflund, M.; Giebl, H.; Hitzenberger, R.; Zibuschka, F.; Puxbaum, H. The contribution of bacterial and fungal spores to the organic carbon content of cloud water, precipitation and aerosols. Atmos. Res. 2002, 64, 109–119. [Google Scholar] [CrossRef]
- China, S.; Wang, B.; Weis, J.; Rizzo, L.; Brito, J.; Cirino, G.G.; Kovarik, L.; Artaxo, P.; Gilles, M.K.; Laskin, A. Rupturing of Biological Spores As a Source of Secondary Particles in Amazonia. Environ. Sci. Technol. 2016, 50, 12179–12186. [Google Scholar] [CrossRef]
- Madelin, T.M.; Johnson, H.E. Fungal and actinomycete spore aerosols measured at different humidities with an aerodynamic particle sizer. J. Appl. Bacteriol. 1992, 72, 400–409. [Google Scholar] [CrossRef]
- Polymenakou, P.N.; Mandalakis, M.; Stephanou, E.G.; Tselepides, A. Particle size distribution of airborne microorganisms and pathogens during an Intense African dust event in the eastern Mediterranean. Environ. Health Perspect. 2008, 116, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.C.; Hsueh, P.R.; Yang, P.C.; Luh, K.T. Clinical and microbiological characteristics of bacteremia caused by Acinetobacter lwoffii. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Regalado, N.G.; Martin, G.; Antony, S.J. Acinetobacter iwoffii: Bacteremia associated with acute gastroenteritis. Travel Med. Infect. Dis. 2009, 7, 316–317. [Google Scholar] [CrossRef] [PubMed]
- Seifert, H.; Kaltheuner, M.; Perdreau-Remington, F. Micrococcus luteus endocarditis: Case report and review of the literature. Zentralbl. Bakteriol. 1995, 282, 431–435. [Google Scholar] [CrossRef]
- Souhami, L.; Feld, R.; Tuffnell, P.G.; Fellner, T. Micrococcus luteus pneumonia: A case report and review of the literature. Med. Pediatr. Oncol. 1979, 7, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Gravato-Nobre, M.J.; Hodgkin, J. Caenorhabditis elegans as a model for innate immunity to pathogens. Cell Microbiol. 2005, 7, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Albertson, D.; Natsios, G.A.; Gleckman, R. Septic Shock with Micrococcus luteus. Arch. Intern. Med. 1978, 138, 487–488. [Google Scholar] [CrossRef]
- Von Eiff, C.; Kuhn, N.; Herrmann, M.; Weber, S.; Peters, G. Micrococcus luteus as a cause of recurrent bacteremia. Pediatr. Infect. Dis. J. 1996, 15, 711–713. [Google Scholar] [CrossRef]
- Mittal, S.; Sharma, M.; Yadav, A.; Bala, K.; Chaudhary, U. Acinetobacter iwoffii an emerging pathogen in neonatal ICU. Infect. Disord. Drug Targets 2015, 15, 184–188. [Google Scholar] [CrossRef]
- Tas, M.Y.; Oguz, M.M.; Ceri, M. Acinetobacter iwoffii Peritonitis in a Patient on Automated Peritoneal Dialysis: A Case Report and Review of the Literature. Case Rep. Nephrol. 2017, 2017, 5760254. [Google Scholar] [CrossRef]
- Ewbank, J.J.; Pujol, N. Local and long-range activation of innate immunity by infection and damage in C. elegans. Curr. Opin. Immunol. 2016, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Acronym | Da (µm) | Volume m3 | PM (µg/m3) |
|---|---|---|---|---|
| 5–11/04/18 | WWTP_RE1 | <1 | 84 | 10 |
| 1–10 | 84 | 13 | ||
| 11–16/04/18 | WWTP_RE2 | <1 | 72 | 12 |
| 1–10 | 72 | 37 |
| Standards and Reagents | Company |
|---|---|
| Acetonitrile (AcN) | Romil LTD (Cambridge, UK) |
| Methanol (MeOH | Romil LTD (Cambridge, UK) |
| Ethanol (EtOH) | Romil LTD (Cambridge, UK) |
| Dichloromethane (DCM) | Romil LTD (Cambridge, UK) |
| 2,2,4-Trimethylpentane (iso-octane, ISO) | Romil LTD (Cambridge, UK) |
| Propan-2-ol (IPA) | Romil LTD (Cambridge, UK) |
| Water (H2O) | Romil LTD (Cambridge, UK) |
| Hydrochloric acid (HCl) 30% | Merck S.p.a. (Vimodrone, Milan, Italy), |
| 2,6-Pyridinedicarboxylic acid (Dipicolinic acid, DPA) | Sigma-Aldrich S.r.l. (Milan, Italy) |
| Muramic acid (MUR) | Sigma-Aldrich S.r.l. (Milan, Italy) |
| Ergosterol (ERG) | Sigma-Aldrich S.r.l. (Milan, Italy) |
| Analyte | Column | Brand | HPLC | MS-MS | Software |
|---|---|---|---|---|---|
| MUR DPA | SeQuant® ZIC®- HILIC column (5 µm polymer 150 × 2.1 mm) + guard column (20 × 2.1 mm) | Merk s.p.a (Vimodrone, Milan, Italy). | 1260 Infinity II system (Agilent Technologies Italy S.p.A. Cernusco sul Naviglio MI, Italy) | API 2000 + ESI source (AB SCIEX S.r.l. Forster City, CA, USA) | Analyst 1.6.2 |
| ERG | Discovery C8 (50 × 2.1 mm, 5 µm) + guard column (20 × 2.1 mm) | Phenomenex (Torrance, CA, USA) | 1290 Infinity system (Agilent Technologies Italy S.p.A. Cernusco sul Naviglio MI, Italy) | G 6460 +APCI source (Agilent Technologies Italy S.p.A. Cernusco sul Naviglio MI, Italy) | Mass Hunter Workstation software B.06.00 |
| n° Microorganisms/m3 | WWTP_RE1 | WWTP_RE2 | |
|---|---|---|---|
| Bacterial Cells | PM < 1 | 2.10 × 105 | 1.70 × 105 |
| PM > 1 | 7.20 × 104 | 4.70 × 105 | |
| Bacterial Spores | PM < 1 | 1.10 × 105 | 1.00 × 105 |
| PM > 1 | 1.70 × 104 | 4.60 × 104 | |
| Fungal Spores | PM < 1 | * n.d. | * n.d. |
| PM > 1 | 2.90 × 103 | 1.10 × 104 |
| Isolate Number | Strain ID | Bacterial Species | BLAST Percent Identity | GenBank Accession Number |
|---|---|---|---|---|
| 1 | AW 25 | Staphylococcus warneri | 99% | NR_025922.1 |
| 2 | CMS 76or | Kocuria polaris | 98% | NR_028924.1 |
| 7 | LMG 7040 | Pseudomonas oryzihabitans | 99% | NR_117269.1 |
| 8 | C58 | Agrobacterium fabrum | 98% | NR_074266.1 |
| 9 | ZS207 | Acinetobacter iwoffii | 93% | CP019143.1 |
| 10 | TA68 | Kocuria rhizophila | 99% | NR_026452.1 |
| 12 | NBRC 12092 | Bacillus pumilus | 99% | NR_112637.1 |
| 13 | DSM 11821 | Bacillus mycoides | 99% | NR_024697 |
| 14 | DSM 13 | Bacillus licheniformis | 99% | NR_118996.1 |
| 15 | DSM 6998 | Moraxella osloensis | 98% | NR_113392.1 |
| 16 | DM 122 | Staphylococcus hominis | 99% | NR_036956.1 |
| 18 | ODN7 | Sphingomonas hankookensis | 98% | NR_116570.1 |
| 19 | G2-1 | Paenarthrobacter nitroguajacolicus | 99% | NR_027199.1 |
| 20 | NCTC 2665 | Micrococcus luteus | 99% | NR_075062.2 |
| 23 | ICB 89 | Stenotrophomonas pavanii | 99% | NR_116793.1 |
| 29 | DSM 20578 | Microbacterium oxydans | 99% | NR_044931.1 |
| 30 | IAM 12423 | Stenotrophomonas maltophilia | 99% | NR_041577.1 |
| 32 | A1920 | Moraxella osloensis | 98% | NR_104936.1 |
| 33 | P 369/06 | Microbacterium phyllosphaerae | 99% | NR_025405.1 |
| 35 | LMG 25348 | Stenotrophomonas pavanii | 99% | NR_118008.1 |
| 36 | B6 | Aquabacterium parvum | 95% | NR_024874.1 |
| 38 | Fussel | Staphylococcus epidermidis | 99% | NR_036904.1 |
| 40 | AE-6 | Micrococcus aloeverae | 99% | NR_134088.1 |
| Antibiotics | μg | A. iwoffii | M. luteus |
|---|---|---|---|
| Ampicillin | 10 | R | 5 cm |
| Tetracycline | 30 | 2.5 cm | 4 cm |
| Chloramphenicol | 30 | 4 cm | 4.5 cm |
| Erythromycin | 15 | 3.5 cm | 3 cm |
| Cephalothin | 30 | R | 3 cm |
| Clindamycin | 2 | R | 3 cm |
| Cefotaxime | 30 | R | 3 cm |
| Cefuroxima | 30 | R | 3 cm |
| Rifamycin | 30 | 2 cm | 3 cm |
| Oxacillin | 1 | R | 1.5 cm |
| Mezlocillin | 75 | 1 cm | 3 cm |
| Amikacin | 30 | 1 cm | 1.5 cm |
| Fosfomycin | 50 | R | R |
| Aztreonam | 30 | 1 cm | R |
| Gentamycin | 10 | 1 cm | 2 cm |
| Tobramycin | 10 | 1 cm | 1.5 cm |
| Carbenicillin | 100 | 1 cm | 3 cm |
| Streptomycin | 25 | 2 cm | 3 cm |
| Penicillin | 10 u | 1 cm | 4 cm |
| Vancomycin | 30 | 1 cm | 3 cm |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruni, E.; Simonetti, G.; Bovone, B.; Casagrande, C.; Castellani, F.; Riccardi, C.; Pomata, D.; Di Filippo, P.; Federici, E.; Buiarelli, F.; et al. Evaluation of Bioaerosol Bacterial Components of a Wastewater Treatment Plant Through an Integrate Approach and In Vivo Assessment. Int. J. Environ. Res. Public Health 2020, 17, 273. https://doi.org/10.3390/ijerph17010273
Bruni E, Simonetti G, Bovone B, Casagrande C, Castellani F, Riccardi C, Pomata D, Di Filippo P, Federici E, Buiarelli F, et al. Evaluation of Bioaerosol Bacterial Components of a Wastewater Treatment Plant Through an Integrate Approach and In Vivo Assessment. International Journal of Environmental Research and Public Health. 2020; 17(1):273. https://doi.org/10.3390/ijerph17010273
Chicago/Turabian StyleBruni, Erika, Giulia Simonetti, Beatrice Bovone, Chiara Casagrande, Federica Castellani, Carmela Riccardi, Donatella Pomata, Patrizia Di Filippo, Ermanno Federici, Francesca Buiarelli, and et al. 2020. "Evaluation of Bioaerosol Bacterial Components of a Wastewater Treatment Plant Through an Integrate Approach and In Vivo Assessment" International Journal of Environmental Research and Public Health 17, no. 1: 273. https://doi.org/10.3390/ijerph17010273
APA StyleBruni, E., Simonetti, G., Bovone, B., Casagrande, C., Castellani, F., Riccardi, C., Pomata, D., Di Filippo, P., Federici, E., Buiarelli, F., & Uccelletti, D. (2020). Evaluation of Bioaerosol Bacterial Components of a Wastewater Treatment Plant Through an Integrate Approach and In Vivo Assessment. International Journal of Environmental Research and Public Health, 17(1), 273. https://doi.org/10.3390/ijerph17010273