The Nitrogen-Removal Efficiency of a Novel High-Efficiency Salt-Tolerant Aerobic Denitrifier, Halomonas Alkaliphile HRL-9, Isolated from a Seawater Biofilter

Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Media
2.2. Strain Isolation
2.3. SEM Analysis and Identification of Strain HRL-9
2.4. Single-Factor Affecting Aerobic Denitrification
2.5. Estimation of Aerobic Denitrification Ability of Strain HRL-9
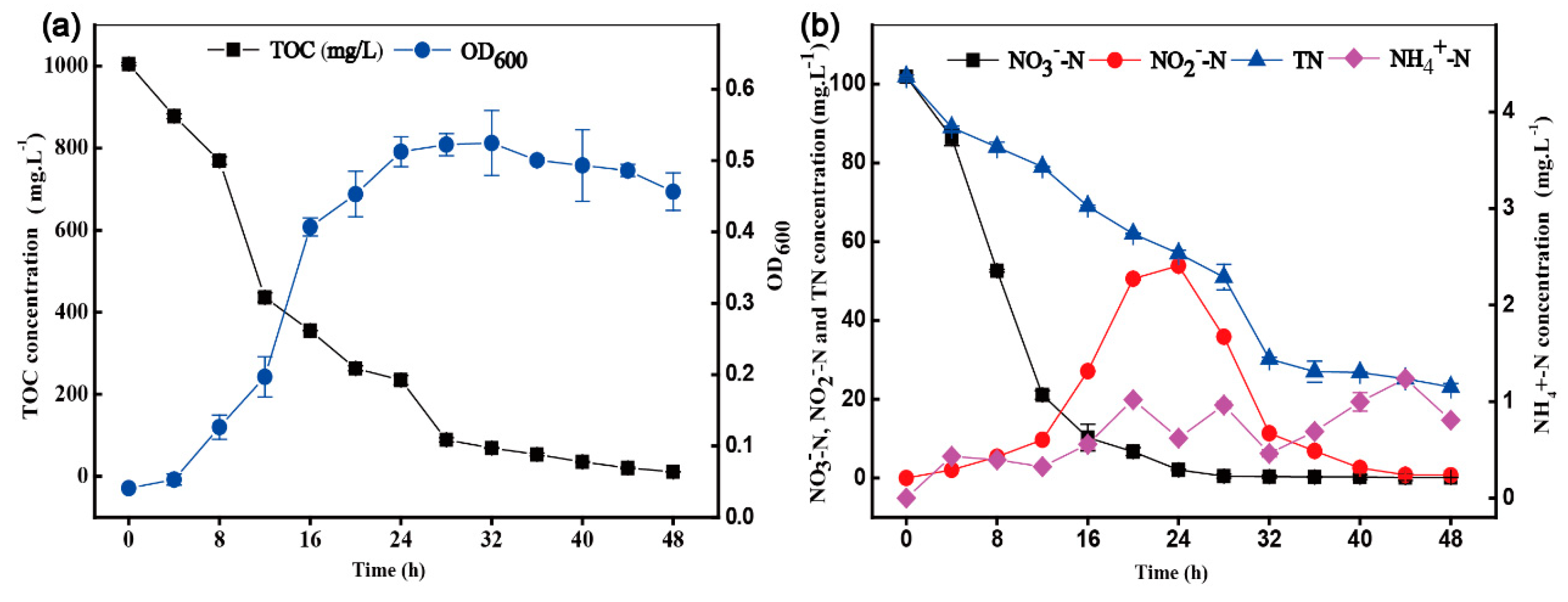
2.6. Nitrogen Balance Analysis
2.7. Amplification of napA and narG Gene
2.8. Analytical Methods
3. Results
3.1. Identification of Strain HRL-9
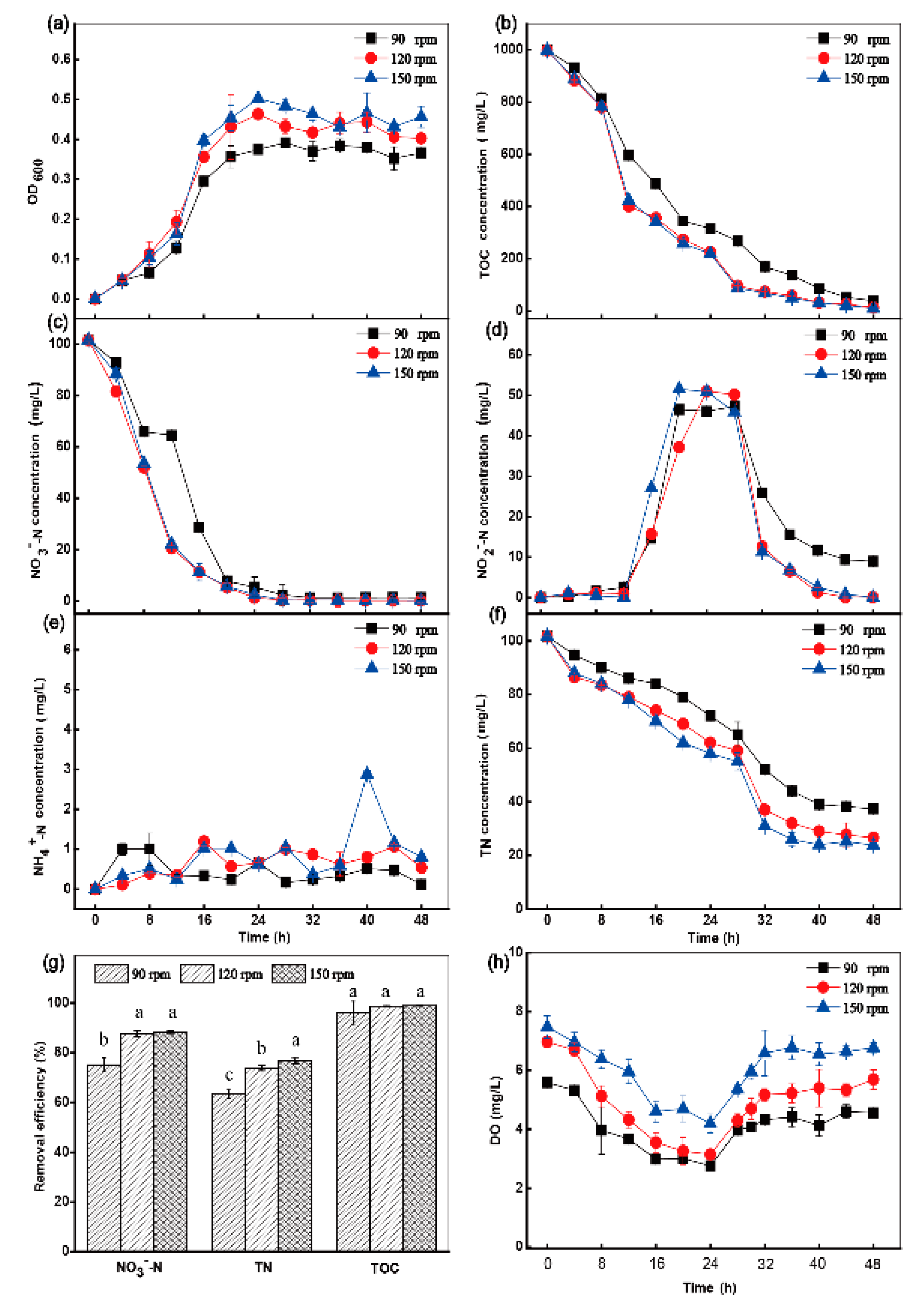
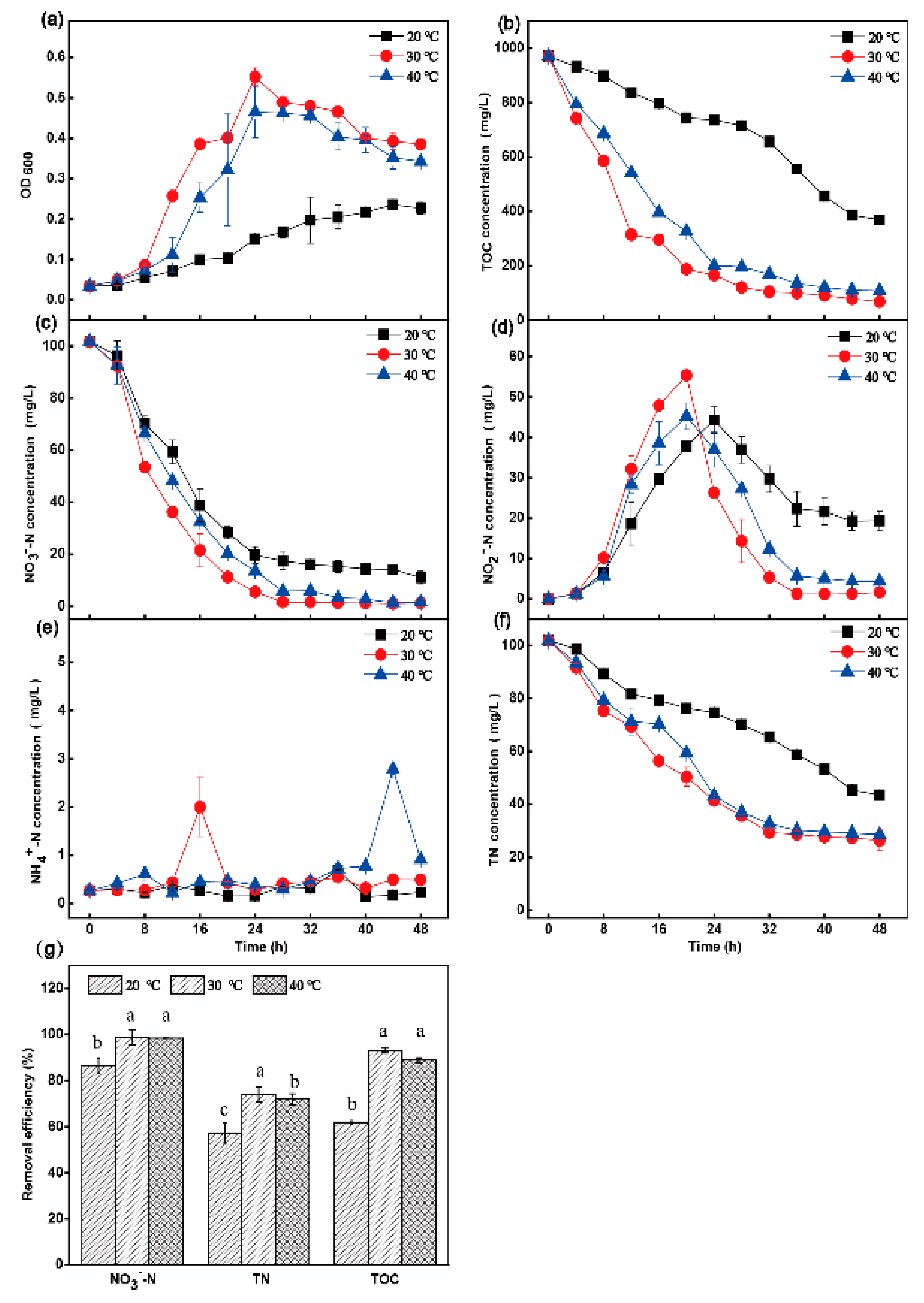
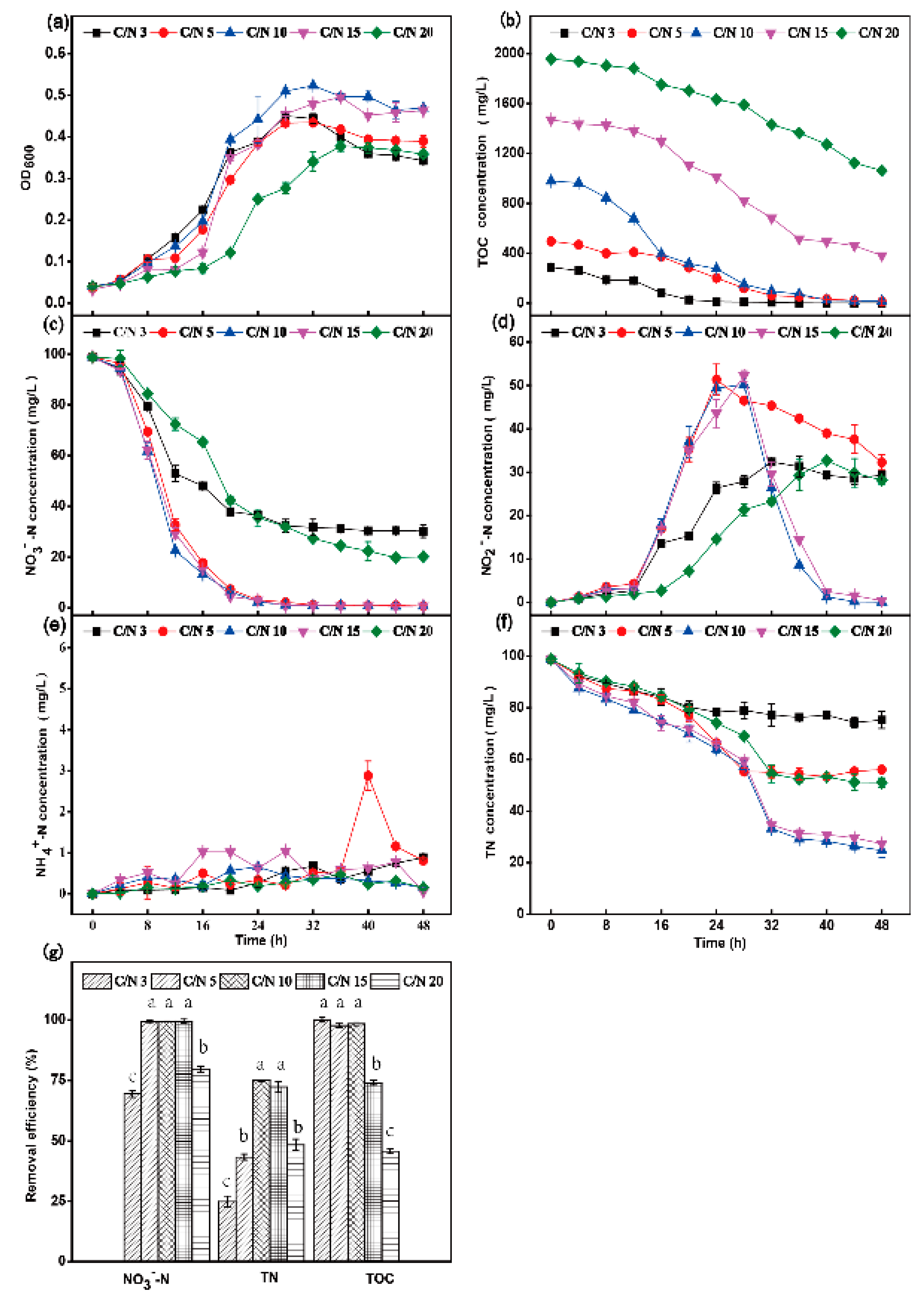
3.2. The Influence of Shaking Speed, Temperature and C/N Ratio on Aerobic Denitrification
3.2.1. Effect of Shaking Speed
3.2.2. Effect of Temperature
3.2.3. Effect of C/N
3.3. Aerobic Denitrification and Nitrogen Balance of Strain HRL-9
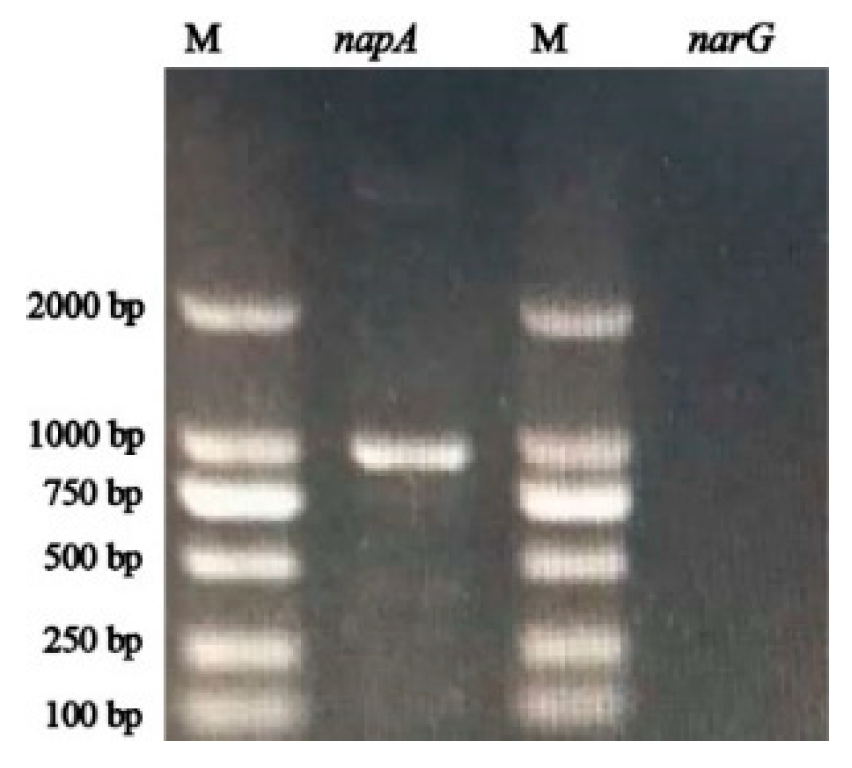
3.4. Amplification of napA and narG Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buschmann, A.; Riquelme, V.A.; Hernándezgonzález, M.C.; Varela, D.; Jiménez, J.E.; Henríquez, L.A.; Vergara, P.A. A review of the impacts of salmonid farming on marine coastal ecosystems in the southeast Pacific. ICES J. Mar. Sci. 2006, 63, 1338–1345. [Google Scholar] [CrossRef]
- Marx Sander, E.; Virdis, B.; Freguia, S. Bioelectrochemical denitrification for the treatment of saltwater recirculating aquaculture streams. ACS Omega 2018, 3, 4252–4261. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.I.M.; Eding, E.H.; Verdegem, M.C.J.; Heinsbroek, L.T.N.; Schneider, O.; Blancheton, J.P.; d’Orbcastel, E.R.; Verreth, J.A.J. New developments in recirculating aquaculture systems in Europe: A perspective on environmental sustainability. Aquac. Eng. 2010, 43, 83–93. [Google Scholar] [CrossRef]
- Martins, C.I.M.; Ochola, D.; Ende, S.S.W.; Eding, E.H.; Verreth, J.A.J. Is growth retardation present in Nile tilapia Oreochromis niloticus cultured in low water exchange recirculating aquaculture systems? Aquaculture 2009, 298, 43–50. [Google Scholar] [CrossRef]
- Rijn, J.V.; Tal, Y.; Schreier, H.J.J.A.E. Denitrification in recirculating systems: Theory and applications. Aquac. Eng. 2006, 34, 364–376. [Google Scholar] [CrossRef]
- Gutierrez-Wing, M.T.; Malone, R.F. Biological filters in aquaculture: Trends and research directions for freshwater and marine applications. Aquac. Eng. 2006, 34, 163–171. [Google Scholar] [CrossRef]
- Hamlin, H.J.; Moore, B.C.; Edwards, T.M.; Larkin, I.L.V.; Boggs, A.; High, W.J.; Main, K.L.; Guillette, L.J., Jr. Nitrate-induced elevations in circulating sex steroid concentrations in female Siberian sturgeon (Acipenser baeri) in commercial aquaculture. Aquaculture. 2008, 281, 118–125. [Google Scholar] [CrossRef]
- Tsai, S.J.; Chen, J.C. Acute toxicity of nitrate on penaeus monodon juveniles at different salinity levels. Aquacultural 2002, 213, 163–170. [Google Scholar] [CrossRef]
- Guo, L.; Chen, Q.; Fang, F.; Hu, Z.; Wu, J.; Miao, A.; Yang, L. Application potential of a newly isolated indigenous aerobic denitrifier for nitrate and ammonium removal of eutrophic lake water. Bioresour. Technol. 2013, 142, 45–51. [Google Scholar] [CrossRef]
- Lycus, P.; Lovise Bothun, K.; Bergaust, L.; Peele Shapleigh, J.; Reier Bakken, L.; Frostegard, A. Phenotypic and genotypic richness of denitrifiers revealed by a novel isolation strategy. ISME J. 2017, 11, 2219–2232. [Google Scholar] [CrossRef]
- Wang, X.; An, Q.; Zhao, B.; Guo, J.S.; Huang, Y.S.; Tian, M. Auto-aggregation properties of a novel aerobic denitrifier Enterobacter sp. strain FL. Appl. Microbiol. Biotechnol. 2018, 102, 2019–2030. [Google Scholar] [CrossRef]
- Wang, H.; Wang, T.; Yang, S.; Liu, X.; Wen, P. Nitrogen removal in oligotrophic reservoir water by a mixed aerobic denitrifying consortium: Influencing factors and immobilization effects. Int. J. Environ. Res. Pub. Health. 2019, 16, 583. [Google Scholar] [CrossRef]
- Ferguson, S.J. Denitrification and its control. Antonie Van Leeuwenhoek 1994, 66, 89–110. [Google Scholar] [CrossRef]
- Wan, C.; Yang, X.; Lee, D.J.; Du, M.; Wan, F.; Chen, C. Aerobic denitrification by novel isolated strain using as nitrogen source. Bioresour. Technol. 2011, 102, 7244–7248. [Google Scholar] [CrossRef]
- Lesley, A.R.; Gijs, K.J. Thiosphaera pantotropha gen. nov. sp. nov. a facultatively anaerobic, facultatively autotrophic sulphur bacterium. Microbiology 1983, 129, 2847–2855. [Google Scholar]
- Zhao, B.; Cheng, D.Y.; Tan, P.; An, Q.; Guo, J.S.J.B.T. Characterization of an aerobic denitrifier Pseudomonas stutzeri strain XL-2 to achieve efficient nitrate removal. Bioresour. Technol. 2017, 250, 564–573. [Google Scholar] [CrossRef]
- Zheng, M. Aerobic denitrification characteristics and mechanism of Pseudomonas stutzeri PCN-1. In Nitrogen Removal Characteristics of Aerobic Denitrifying Bacteria and Their Applications in Nitrogen Oxides Emission Mitigation; Springer: Singapore, 2019; pp. 51–69. [Google Scholar]
- Zhang, Y.; Shi, Z.; Chen, M.; Dong, X.; Zhou, J. Evaluation of simultaneous nitrification and denitrification under controlled conditions by an aerobic denitrifier culture. Bioresour. Technol. 2018, 175, 602–605. [Google Scholar] [CrossRef]
- Medhi, K.; Mishra, A.; Thakur, I.S. Genome sequence of a heterotrophic nitrifier and aerobic denitrifier, paracoccus denitrificans strain ISTOD1, isolated from wastewater. Genome Announc. 2018, 6, e00210-18. [Google Scholar] [CrossRef]
- Feng, Y.; Feng, J.; Shu, Q.L. Isolation and characterization of heterotrophic nitrifying and aerobic denitrifying Klebsiella pneumoniae and Klebsiella variicola strains from various environments. J. Appl. Microbiol. 2018, 124, 1195–1211. [Google Scholar] [CrossRef]
- Liu, Y.; Ai, G.M.; Miao, L.L.; Liu, Z.P. Marinobacter strain NNA5, a newly isolated and highly efficient aerobic denitrifier with zero N2O emission. Bioresour. Technol. 2016, 206, 9–15. [Google Scholar] [CrossRef]
- Duan, J.; Fang, H.; Su, B.; Chen, J.; Lin, J. Characterization of a halophilic heterotrophic nitrification-aerobic denitrification bacterium and its application on treatment of saline wastewater. Bioresour. Technol. 2015, 179, 421–428. [Google Scholar] [CrossRef]
- Vyrides, I.; Stuckey, D. Adaptation of anaerobic biomass to saline conditions: Role of compatible solutes and extracellular polysaccharides. Enzym. Microb. Technol. 2009, 44, 46–51. [Google Scholar] [CrossRef]
- Guo, Y.; Zhou, X.; Li, Y.; Li, K.; Wang, C.; Liu, J.; Yan, D.; Liu, Y.; Yang, D.; Xing, J. Heterotrophic nitrification and aerobic denitrification by a novel Halomonas campisalis. Biotechnol. Lett. 2013, 35, 2045–2049. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.Y.; Liu, Y.; Gao, X.Y.; Ai, G.M.; Miao, L.L.; Liu, Z.P. Characterization of a marine origin aerobic nitrifying–denitrifying bacterium. J. Biosci. Bioeng. 2012, 114, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Roussel-Delif, L.; Tarnawski, S.; Hamelin, J.; Philippot, L.; Aragno, M.; Fromin, N.J.M.E. Frequency and diversity of nitrate reductase genes among nitrate-dissimilating Pseudomonas in the rhizosphere of perennial grasses grown in field conditions. Microb. Ecol. 2005, 49, 63–72. [Google Scholar] [CrossRef]
- Angela, H.; Shapleigh, J.P. Physiological roles for two periplasmic nitrate reductases in Rhodobacter sphaeroides 2.4.3 (ATCC 17025). J. Bacteriol. 2011, 193, 6483–6489. [Google Scholar]
- Berendes, F.; Gottschalk, G.; Heine-Dobbernack, E.; Moore, E.R.B.; Tindall, B.J. Halomonas desiderata sp. nov, a new alkaliphilic, halotolerant and denitrifying bacterium isolated from a municipal sewage works. Syst. Appl. Microbiol. 1996, 19, 158–167. [Google Scholar] [CrossRef]
- Shapovalova, A.A.; Khijniak, T.V.; Tourova, T.P.; Sorokin, D.Y. Halomonas chromatireducens sp. nov., a new denitrifying facultatively haloalkaliphilic bacterium from solonchak soil capable of aerobic chromate reduction. Microbiology 2009, 78, 102–111. [Google Scholar] [CrossRef]
- Ji, B.; Wang, H.; Yang, K. Tolerance of an aerobic denitrifier (Pseudomonas stutzeri) to high O2 concentrations. Biotechnol. Lett. 2014, 36, 719–722. [Google Scholar] [CrossRef]
- Ji, B.; Yang, K.; Wang, H.; Zhou, J.; Zhang, H. Aerobic denitrification by Pseudomonas stutzeri C3 incapable of heterotrophic nitrification. Bioprocess Biosyst. Eng. 2015, 38, 407–409. [Google Scholar] [CrossRef]
- Zhou, M.; Ye, H.; Zhao, B. Isolation and characterization of a novel heterotrophic nitrifying and aerobic denitrifying bacterium Pseudomonas stutzeri KTB for bioremediation of wastewater. Biotechnol. Bioprocess Eng. 2014, 19, 231–238. [Google Scholar] [CrossRef]
- Huang, T.; Guo, L.; Zhang, H.; Su, J.; Wen, G.; Zhang, K. Nitrogen-removal efficiency of a novel aerobic denitrifying bacterium, Pseudomonas stutzeri strain ZF31, isolated from a drinking-water reservoir. Bioresour. Technol. 2015, 196, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Domenech, C.M.; Martinez-Checa, F.; Quesada, E.; Bejar, V. Halomonas fontilapidosi sp. nov., a moderately halophilic, denitrifying bacterium. Int. J. Syst. Evol. Microbiol. 2009, 59, 1290–1296. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Jeong, S.Y.; Yoon, S.J.; Cho, S.J.; Kim, Y.H.; Kim, M.J.; Ryu, E.Y.; Lee, S.J.J.J.O.B. Aerobic denitrification of Pseudomonas putida AD-21 at different C/N ratios. J. Biosci. Bioeng. 2008, 106, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.K.; Tseng, S.K. Nitrate reduction by Citrobacter diversus under aerobic environment. Appl. Microbiol. Biotechnol. 2001, 55, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Ding, W.; Feng, L.J.; Kong, Y.; Xu, J.; Xu, X.Y. Isolation of aerobic denitrifiers and characterization for their potential application in the bioremediation of oligotrophic ecosystem. Bioresour. Technol. 2012, 108, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L. Dentrifying genes in bacterial and archaeal genomes. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2002, 1577, 355–376. [Google Scholar] [CrossRef]
- Zhang, W.; Yan, Z.C.; Shen, J.; Wei, R.; Gao, Y. Characterization of aerobic denitrifying bacterium Pseudomonas mendocina strain GL6 and its potential application in wastewater treatment plant effluent. Int. J. Environ. Res. Public Health 2019, 16, 364. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Initial TN mg·L−1 | Final TN mg·L−1 | Intracellular-N/% | Nitrogen Remove/% | |||
|---|---|---|---|---|---|---|---|
| NO3−-N | NO2−-N | NH4+-N | Organic-N | ||||
| Nitrate | 101.4 ± 0.02 | 1.24 ± 0.01 | 0.10 ± 0.01 | 0.50 ± 0.10 | 2.75 ± 0.32 | 21.7 ± 0.28 | ≈74.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, J.; Wei, C.; Ma, H.; Dai, M.; Fan, J.; Liu, Y.; Wu, Y.; Han, R. The Nitrogen-Removal Efficiency of a Novel High-Efficiency Salt-Tolerant Aerobic Denitrifier, Halomonas Alkaliphile HRL-9, Isolated from a Seawater Biofilter. Int. J. Environ. Res. Public Health 2019, 16, 4451. https://doi.org/10.3390/ijerph16224451
Ren J, Wei C, Ma H, Dai M, Fan J, Liu Y, Wu Y, Han R. The Nitrogen-Removal Efficiency of a Novel High-Efficiency Salt-Tolerant Aerobic Denitrifier, Halomonas Alkaliphile HRL-9, Isolated from a Seawater Biofilter. International Journal of Environmental Research and Public Health. 2019; 16(22):4451. https://doi.org/10.3390/ijerph16224451
Chicago/Turabian StyleRen, Jilong, Chenzheng Wei, Hongjing Ma, Mingyun Dai, Jize Fan, Ying Liu, Yinghai Wu, and Rui Han. 2019. "The Nitrogen-Removal Efficiency of a Novel High-Efficiency Salt-Tolerant Aerobic Denitrifier, Halomonas Alkaliphile HRL-9, Isolated from a Seawater Biofilter" International Journal of Environmental Research and Public Health 16, no. 22: 4451. https://doi.org/10.3390/ijerph16224451
APA StyleRen, J., Wei, C., Ma, H., Dai, M., Fan, J., Liu, Y., Wu, Y., & Han, R. (2019). The Nitrogen-Removal Efficiency of a Novel High-Efficiency Salt-Tolerant Aerobic Denitrifier, Halomonas Alkaliphile HRL-9, Isolated from a Seawater Biofilter. International Journal of Environmental Research and Public Health, 16(22), 4451. https://doi.org/10.3390/ijerph16224451