Molecular Modification of Fluoroquinolone-Biodegrading Enzymes Based on Molecular Docking and Homology Modelling


Abstract
1. Introduction
2. Materials and Methods
2.1. Molecular Docking Method
2.2. Homology Modelling Method
2.3. Gaussian Calculation Method
3. Results and Discussion
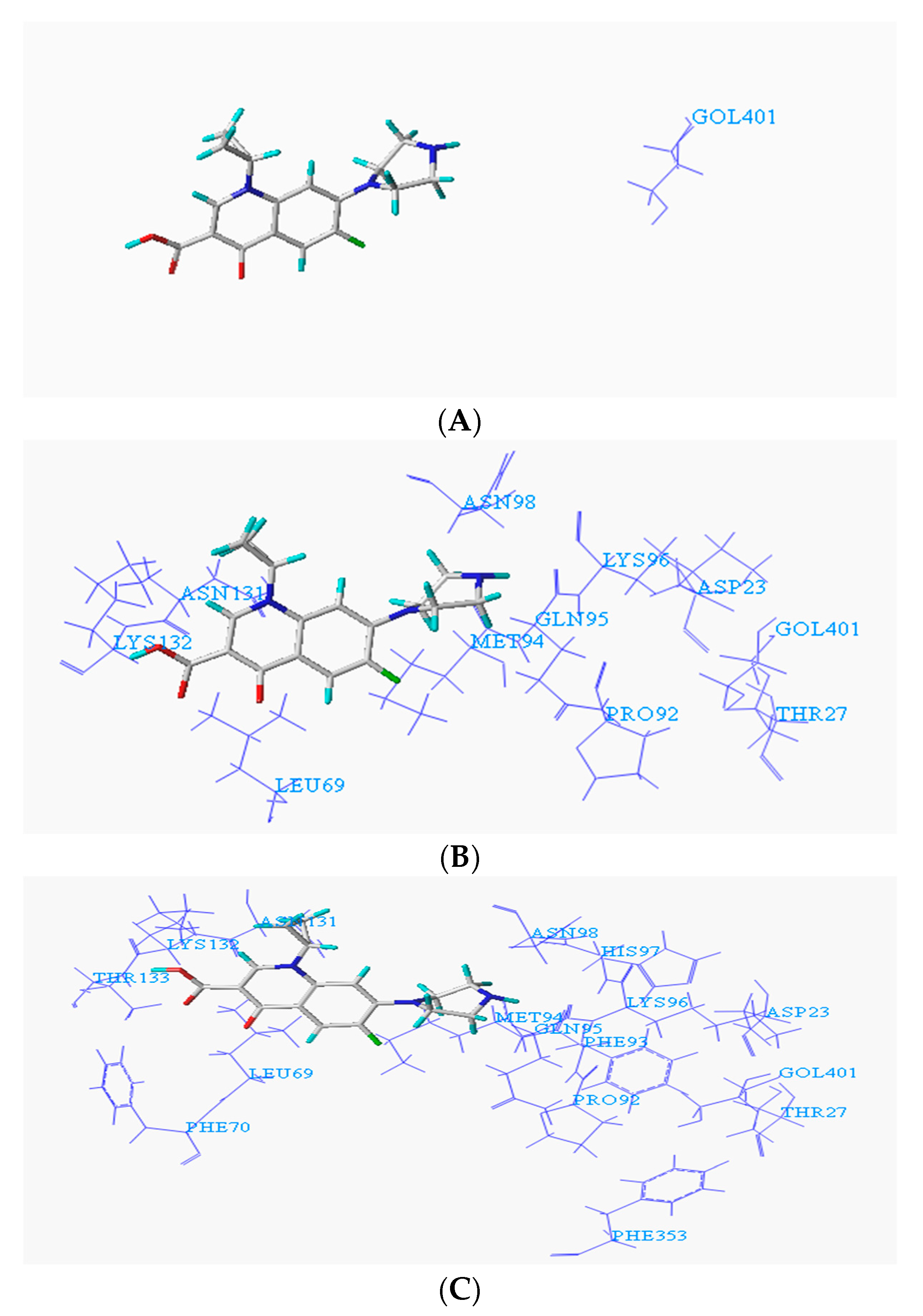
3.1. Determination of Key Amino Acid Residues of Fluoroquinolone Degrading Enzymes Based on Molecular Docking Technology
3.2. Modification of Fluoroquinolone Degrading Enzymes Based on Homology Modelling
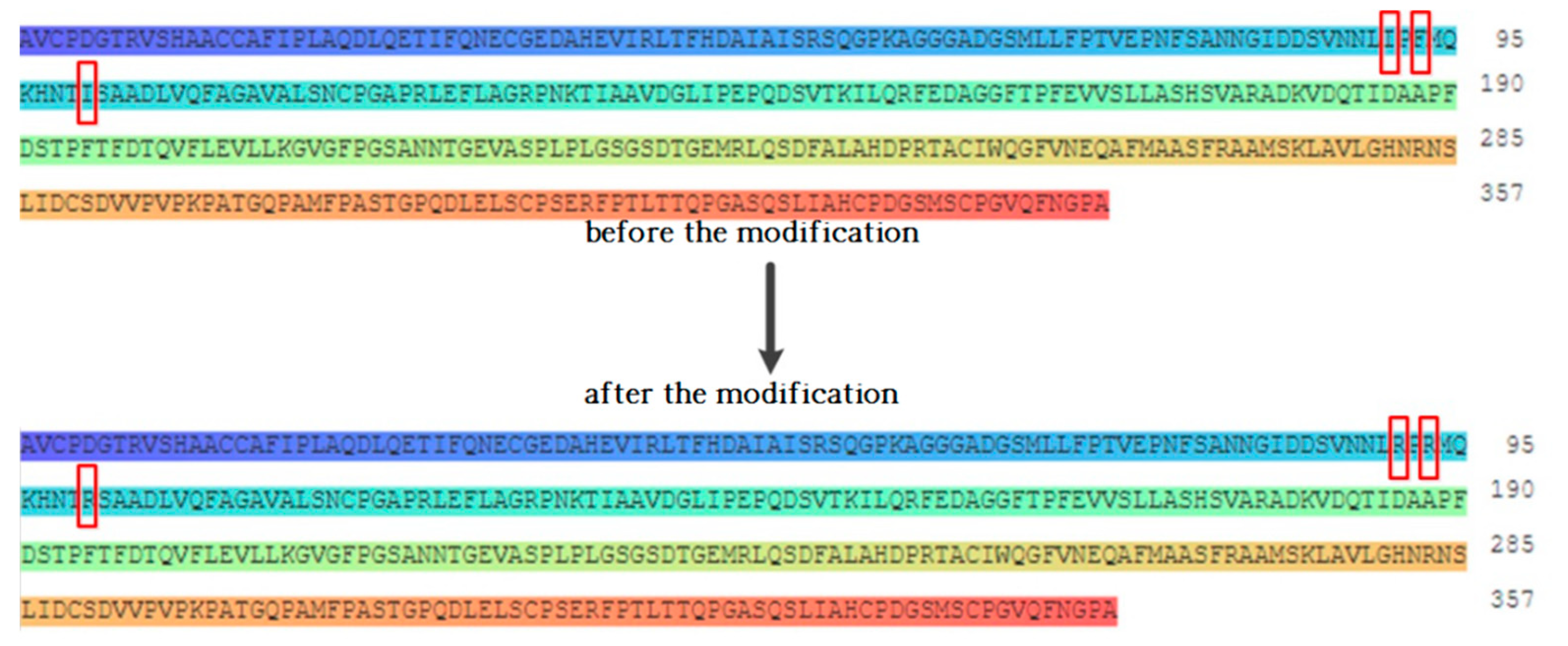
3.2.1. Design of the Modification Scheme for Fluoroquinolone Degrading Enzymes
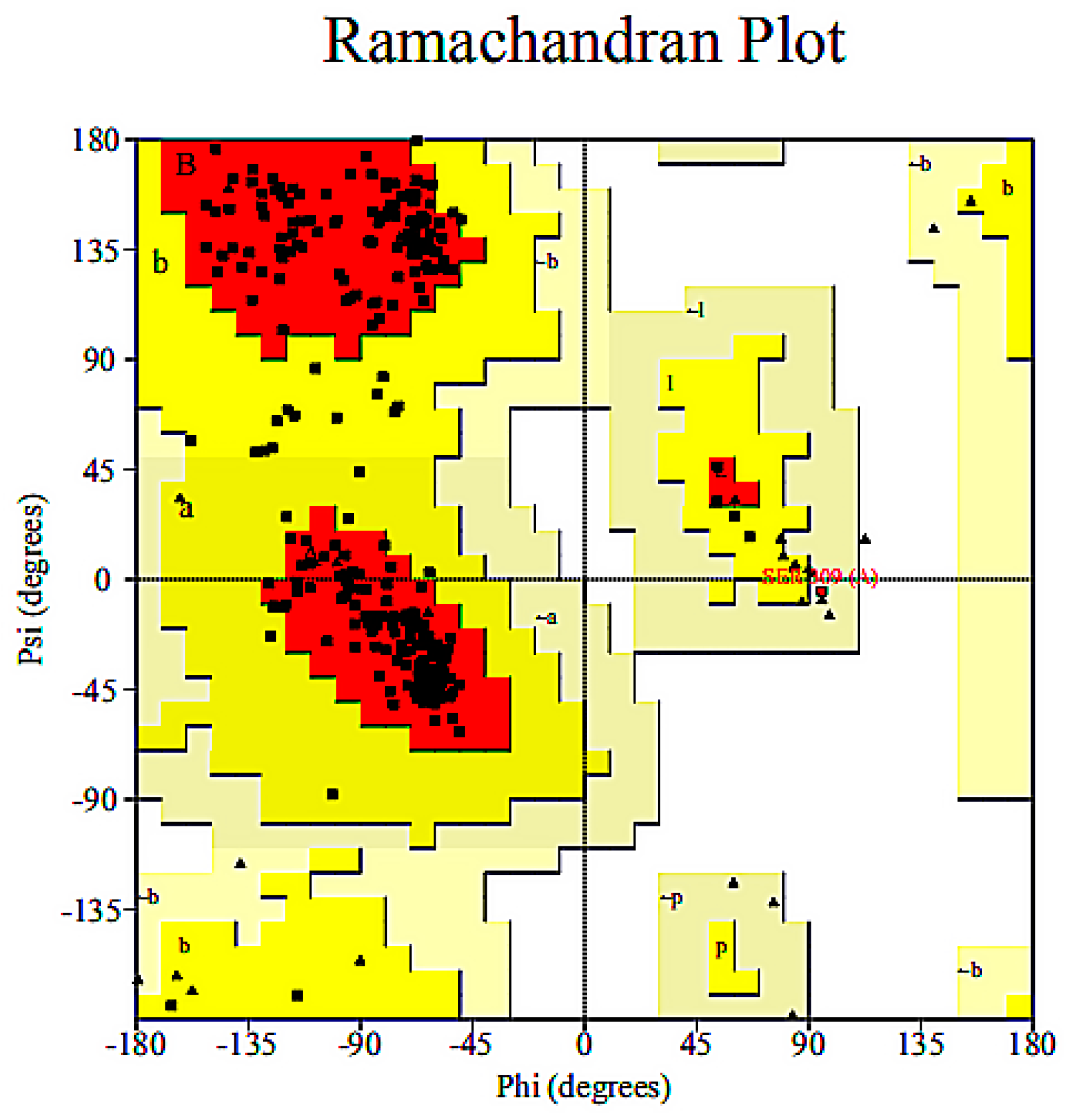
3.2.2. Homology Modelling and Model Validation of Fluoroquinolone Molecular Degrading Enzymes
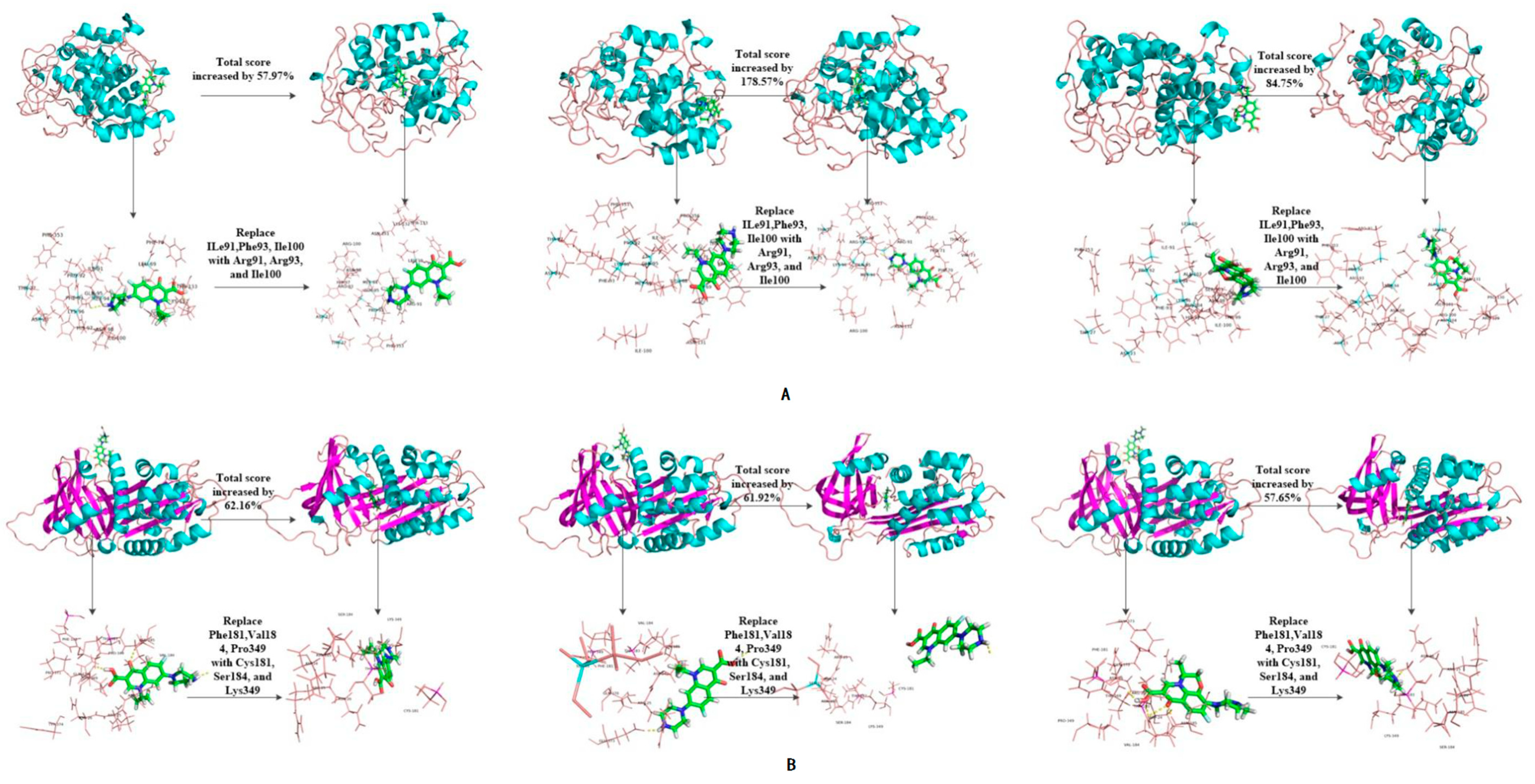
3.3. Evaluation of the Binding Affinity between Novel Enzymes and Target Fluoroquinolones
3.4. Calculation of the Energy Barriers for the Degradation of the Target Fluoroquinolone by Novel Degrading Enzymes
3.5. Analysis of the Effect of the Degrading Enzymes before and after Modification on the Degradation of Organic Matter
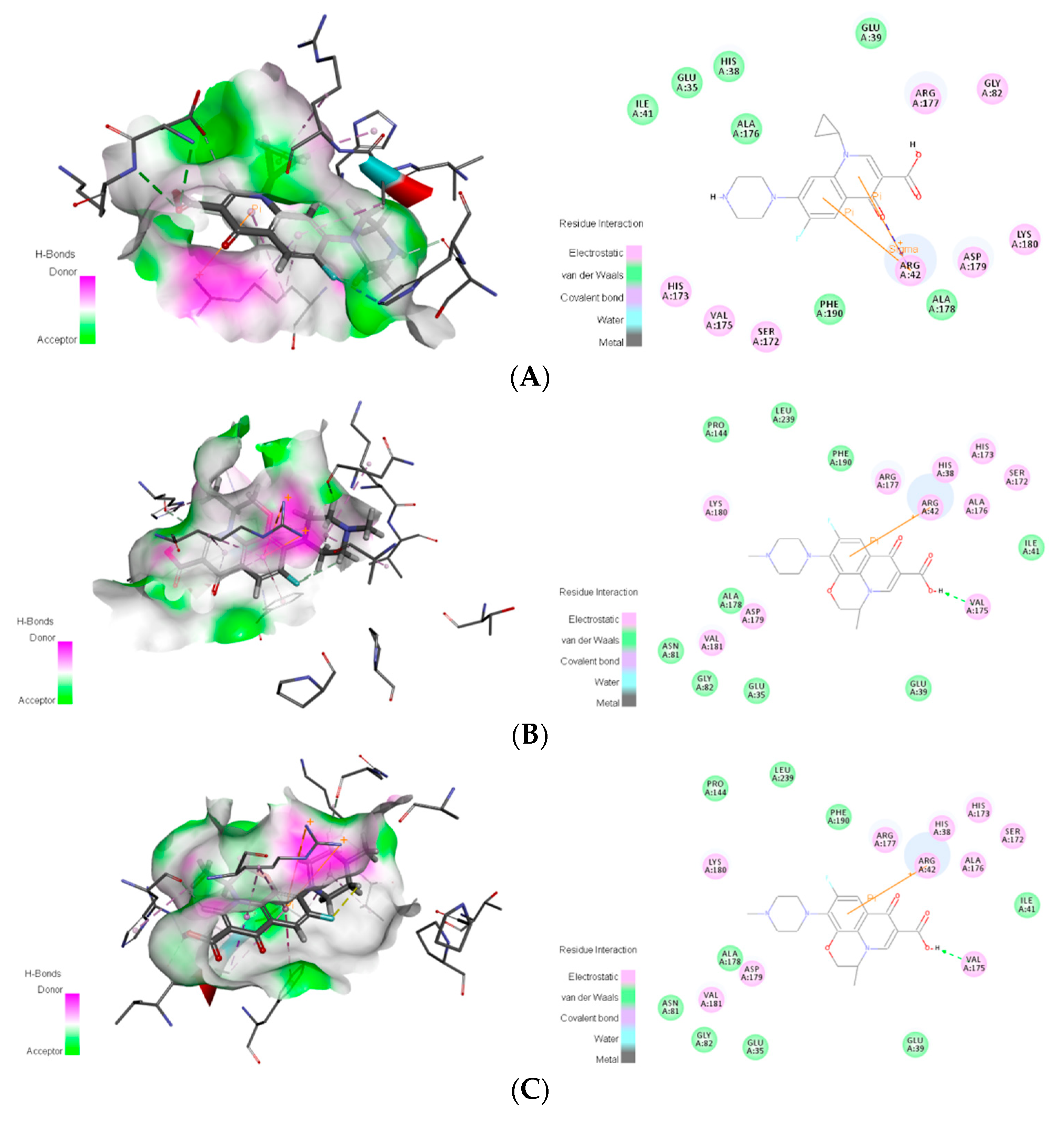
3.6. Mechanism Analysis of the Binding of the Novel Degrading Enzymes and Target Fluoroquinolones
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rusu, A.; Hancu, G.; Uivaroşi, V. Fluoroquinolone pollution of food, water and soil, and bacterial resistance. Environ. Chem. Lett. 2015, 13, 21–36. [Google Scholar] [CrossRef]
- Łuczkiewicz, A.; Jankowska, K.; Fudala-Książek, S.; Olańczuk-Neyman, K. Antimicrobial resistance of fecal indicators in municipal wastewater treatment plant. Water Res. 2010, 44, 5089–5097. [Google Scholar] [CrossRef] [PubMed]
- Van Doorslaer, X.; Dewulf, J.; Van Langenhove, H.; Demeestere, K. Fluoroquinolone antibiotics: An emerging class of environmental micropollutants. Sci. Total Environ. 2014, 500–501, 250–269. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Xie, Y.J.; Yu, Y.J.; Liu, X.H.; Zhao, S.N.; Cui, B.S.; Bai, J.H. Occurrence and Partitioning of Antibiotics in the Water Column and Bottom Sediments from the Intertidal Zone in the Bohai Bay, China. Wetlands 2016, 36, 167–179. [Google Scholar] [CrossRef]
- Meng, F.G.; Gao, G.H.; Yang, T.T.; Chen, X.; Chao, Y.Q.; Na, G.S.; Ge, L.; Huang, L.N. Effects of fluoroquinolone antibiotics on reactor performance and microbial community structure of a membrane bioreactor. Chem. Eng. J. 2015, 280, 448–458. [Google Scholar] [CrossRef]
- Gartiser, S.; Urich, E.; Alexy, R.; Kümmerer, K. Ultimate biodegradation and elimination of antibiotics in inherent tests. Chemosphere 2007, 67, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhang, Y.L.; Zhou, X.F. Occurrence and removal of fluoroquinolones in municipal sewage: A review. Environ. Pollut. Control 2016, 38, 75–80. [Google Scholar]
- Jia, A.; Wan, Y.; Xiao, Y.; Hu, J. Occurrence and fate of quinolone and fluoroquinolone antibiotics in a municipal sewage treatment plant. Water Res. 2012, 46, 387–394. [Google Scholar] [CrossRef]
- Amorim, C.L.; Moreira, I.S.; Maia, A.S.; Tiritan, M.E.; Castro, P.M.L. Biodegradation of ofloxacin, norfloxacin, and ciprofloxacin as single and mixed substrates by Labrys portucalensis F11. Appl. Microbiol. Biotechnol. 2014, 98, 3181–3190. [Google Scholar] [CrossRef]
- Martens, R.; Wetzstein, H.G.; Zadrazil, F.; Capelari, M.; Hoffmann, P.; Schmeer, N. Degradation of the fluoroquinolone enrofloxacin by wood-rotting fungi. Appl. Environ. Microbiol. 1996, 62, 4206–4209. [Google Scholar]
- Zhang, X.Y.; He, G.Q.; Chen, Q.H. Using of the Rational and Irrational Protein Design Strategies in Enzyme Engineering. Bull. Sci. Technol. 2007, 23, 191–197. [Google Scholar]
- Zhang, S.; Barr, B.K.; Wilson, D.B. Effects of noncatalytic residue mutations on substrate specificity and ligand binding of Thermobifida fusca endocellulase Cel6A. Eur. J. Biochem. 2000, 267, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, T.; Hakamada, Y.; Hatada, Y.J.; Kobayashi, T.; Shirai, T. Thermostabilization by replacement of specific residues with lysine in a Bacillus alkaline cellulase: Building a structural model and implications of newly formed double intrahelical salt bridges. Protein Eng. Des. Sel. 2001, 14, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Vassiliadis, V.S.; Raúl, C. Powell method Powell Method. Encycl. Optim. 2001, 2001–2003. [Google Scholar]
- Li, X.L.; Ye, L.; Wang, X.X.; Wang, X.Z.; Liu, H.L.; Qian, X.P.; Zhu, Y.L.; Yu, H.X. Molecular docking, molecular dynamics simulation, and structure-based 3D-QSAR studies on estrogenic activity of hydroxylated polychlorinated biphenyls. Sci. Total Environ. 2012, 441, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Güette-Fernández, J.R.; Meléndez, E.; Maldonado-Rojas, W.; Ortega-Zúñiga, C.; Olivero-Verbel, J.; Parés-Matos, E.I. A molecular docking study of the interactions between human transferrin and seven metallocene dichlorides. J. Mol. Graph. Model. 2017, 75, 250–265. [Google Scholar] [CrossRef] [PubMed]
- Bordoli, L.; Kiefer, F.; Arnold, K.; Benkert, P.; Battey, J.; Schwede, T. Protein structure homology modeling using SWISS-MODEL workspace. Nat. Protoc. 2008, 4, 1–13. [Google Scholar] [CrossRef]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Macarthur, M.W.; Moss, D.S.; Thornton, J. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revision E.01; Gaussian Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Qu, R.J.; Liu, H.X.; Feng, M.B.; Yang, X.; Wang, Z.Y. Investigation on Intramolecular Hydrogen Bond and Some Thermodynamic Properties of Polyhydroxylated Anthraquinones. J. Chem. Eng. Data 2012, 57, 2442–2455. [Google Scholar] [CrossRef]
- Yang, W.H.; Wang, Z.Y.; Liu, H.L.; Yu, H.X. Exploring the binding features of polybrominated diphenyl ethers as estrogen receptor antagonists: Docking studies. Sar Qsar Environ. Res. 2010, 21, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, J.M.; Mant, C.T.; Hodges, R.S. Determination of intrinsic hydrophilicity/hydrophobicity of amino acid side chains in peptides in the absence of nearest-neighbor or conformational effects. Biopolymers 2006, 84, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C.; Chu, Z.H.; Du, X.Y.; Qiu, Y.L.; Li, Y. Combined molecular docking, homology modelling and density functional theory studies to modify dioxygenase to efficiently degrade aromatic hydrocarbons. RSC Adv. 2019, 9, 11465–11475. [Google Scholar] [CrossRef]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.B.; Liu, C.; Ren, L.R.; Tong, H.; Li, W.M.; Wu, D. Studies on pyrolysis mechanism of syringol as lignin model compound by quantum chemistry. J. Fuel Chem. Technol. 2013, 41, 657–666. [Google Scholar] [CrossRef]
- Wetzstein, H.G.; Schmeer, N.; Karl, W. Degradation of the fluoroquinolone enrofloxacin by the brown rot fungus Gloeophyllum striatum: Identification of metabolites. Appl. Environ. Microbiol. 1998, 64, 4272–4281. [Google Scholar]
- Wetzstein, H.G.; Stadler, M.; Tichy, H.V.; Dalhoff, A.; Karl, W. Degradation of ciprofloxacin by basidiomycetes and identification of metabolites generated by the brown rot fungus Gloeophyllum striatum. Appl. Environ. Microbiol. 1999, 65, 1556. [Google Scholar]
- Foglar, L.; BrišKi, F. Wastewater denitrification process—The influence of methanol and kinetic analysis. Process Biochem. 2003, 39, 95–103. [Google Scholar] [CrossRef]
- Yan, Y.W.; Xu, P.P.; Huang, X.A.; Lai, Y.N.; Li, X.Y.; Fu, L.C. Study on the Anti-influenza Mechanism of Paeoniae Radix Rubra Base on Molecular Docking Simulation. J. Chin. Med. Mater. 2017, 40, 430–435. [Google Scholar]
- Chu, Z.H.; Li, Y. Designing modified polybrominated diphenyl ether BDE-47, BDE-99, BDE-100, BDE-183 and BDE-209 molecules with decreased estrogenic activities using 3D-QSAR, pharmacophore models coupled with resolution V of the 210−3 fractional factorial design and molecular docking. J. Hazard. Mater. 2019, 364, 151–162. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Degrading Bacteria Type | PDB ID | Bacterium | Amino Acid Residues within 5 Å Range of the Target Fluoroquinolone Molecule | ||
|---|---|---|---|---|---|
| Combined with CIP | Combined with NOR | Combined with OFL | |||
| Aerobic bacteria | 1ARP | Arthromyces ramosus | Ile81, Ala82, His83, Ser84, Arg104, Ile108, His110, Val112, Ser113, Phe144, Asn143 | Ile81, Ala82, His83, Ser84, Asn85, Ile86, Glu101, Arg104, Ile108, Phe114 | Ile81, Ala82, His83, Ser84, Asn85, Ile86, Leu88, Thr97, Ile100, Glu101, Arg104 |
| 1EI5 | Ochrobactrum anthropi | Asn155, Asp481, Cys61, Ser62, Ala482, Ala290, Leu509, Tyr153, Pro483, Ser152, Arg295, Cys296, His287, Tyr151, Trp508, Thr375, Glu409, Ser376, Ser379, Phe373, Pro377, Arg406, Val392, Glu378 | Cys296, Gln297, Arg295, Ala290, His287, Ala318, Glu315, Asn275, Ser319, Tyr270, Tyr153, Pro483, Tyr151, Leu509, Thr375, Glu409, Trp508, Ser376, Arg406 | Trp299, Gln297, Lys65, His287, Glu278, Leu276, Ser62, His277, Asn275, Ser319, Tyr270, Ala290, Pro269, Asn106, Tyr153, Ser408, Tyr151, Val392, Trp508 | |
| 1GYC | Trametes versicolor | Asn340, Asn341, Ala329, Phe338, Ser343, Asn333, Asn336, Thr335, Met57, Thr56, Asn54, Leu158, Ala155, Ala24, Tyr152, Thr154, Asp23, His153, Arg22, Phe20, Pro319, Ala325, Leu326, Glu381, Asn327, Thr383, Thr438, Asn436, Pro385, Asp435, Ala386, Ile203, Arg121, Ala1, Ser33, Thr144, His216, Val30, Leu174, Pro123, Val145, Pro34, Asn249, Ala186, Ile298, Leu35, Asp234, Thr147, Phe31, Ile188, Val187, Leu232, Val297, Ile146, Pro32, Asn172, Leu185, Thr219, Asn217, Glu142, Ser143, Asn251, Gln252 | Pro319, Leu326, Thr383, Asn340, Pro385, Ile298, Thr219, Asn217, Asn249, Asn251, Gln252, Ala325, Thr438, Ala329, Asp435, Leu232, Val297, Asp234, His216, Glu142, Ser143, Glu381, Asn327, Asn436, Asn341, Ala386, Pro123, Val145, Ser33, Pro32, Asn172, Phe31, Asn189, Leu174, Phe338, Ser343, Thr335, Arg121, Thr147, Leu35, Pro34, Ile171, Ile188, Phe31, Val30, Ala186, Asn333, Asn336, Met57, Thr56, Tyr152, Asn54, His153, Asp23, Phe20, Leu158, Ala155, Thr154, Ala24, Arg22 | Pro319, Asn436, Pro385, Asn340, Asn341, Thr335, Gln252, Val145, Pro123, Val187, Pro32, Leu174, Glu381, Ala325, Thr383, Asp435, Ala386, Phe338, Asn333, Asn249, Thr219, Leu232, Ile188, His216, Asp234, Ile298, Thr438, Leu326, Asn327, Ala329, Ser343, Asn336, Asn251, Thr144, Asn189, Pro34, Asn217, Ser33, Asn172, Ala24, Met57, Arg22, Tyr152, Thr56, His153, Glu142, Ser143, Leu35, Ile188, Ile146, Thr147, Val297, Asn54, Asp23, Phe20, Ala155, Thr154, Leu158 | |
| 1YZP | Phanerochaete chrysosporium | Phe70, Thr133, Leu69, Lys132, Phe353, Asn131, Pro92, Met94, Gln95, Phe93, Lys96, Asn98, Thr27, His97, Asp23 | Val73, Thr72, Pro356, Pro71, Phe70, Leu69, Leu68, Phe353, Ile91, Asn131, Pro92, Met94, Gln95, Phe93, Lys96, Thr27, Asp23 | Leu69, Phe353, Ala102, Asn131, Pro130, Arg129, Pro92, Met94, Ser101, Asp104, Phe93, Ile100, Asn98, Lys96, Thr27, His97, Thr99, Asp23 | |
| 4DTE | Nocardia | Phe181, Pro371, Glu373, Gln370, Pro349, Thr183, Thr369, Asn26, Val184, Asn185, Arg25, Asp24 | Ser55, Glu373, Gln370, Arg25, Asn185, Asp24, Thr183, Val184 | Glu373, Gln370, Asn26, Arg25, Asp24, Asn185, Thr369, Thr183, Phe181, Val184, Pro349 | |
| Anaerobic bacteria | 3NQA | Methanothermobacter thermautotrophicus | Pro46, Asp20, Asp70, Lys42, Tyr206, Arg203, Ala18, Lys72, Ile96, Leu123, Val20, Ile200, Met126, His128, Pro180, Ala184, Val155, Ser127, Gln125, Ser179, Val182, Pro157, Ser158 | Pro46, Tyr45, Asp20, Lys72, Asp70, Ile96, Lys42, Tyr206, Ala18, Arg203, Val201, Ser204, Ile200, Met126, Val155, Ser127, Pro180, Ala184, Gln185, Val182 | Pro46, Asp20, Asp70, Lys72, Ile96, Lys42, Tyr206, Ala18, Arg203, Leu123, Val201, Ile200, Thr124, Met126, Val155, His128, Pro180, Ala184, Glu125, Ser127, Ser179, Gln185, Val182, Pro157, Ser158 |
| 1E1D | Desulfovibrio vulgaris | Ala435, Tyr437, Cys459, Cys406, Ala497, Asp407, Cys434, Tyr493, Lys496, Glu494, Tyr161, Ser291, Glu268, Met269, Trp292, Trp293, Thr71, Ile70, His244, Gln294, Asn287, Tyr288, His266, Cys312, Asn311, Ser242, Gln295, Asn311, Cys312, Thr310, Glu298, Phe299, Leu308, Leu313, Val314 | Tyr161, Thr310, Leu308, Asn311, His244, Lys496, Cys312, Ser242, Glu494, Met296, Ala497, Cys459, Glu268, His266, Phe299, Tyr493, Trp292, Gln295, Cys406, Cys434, Trp293, Asp407, Ser291, Ala435 | Ile70, Tyr61, Lys496, Glu494, Thr310, Ala497, Asn311, Tyr493, Cys312, Leu313, Cys459, His244, Leu308, Cys406, Glu268, Ser242, Asp407, Cys434, Ala435, Trp292, Trp293, His266, Ser291, Gln294, Tyr437, Phe299, Glu298 | |
| 3EZX | Methanosarcina barkeri | Met48, Lys49, Met72, Val46, Leu44, Ser45, His106, Ile108, Ile40, Leu155, Pro187, Ala186, His110, Leu112, Glu203, Asn204, Val113, Ala205, Ala202, Ala206, Ala208 | Met48, His110, Leu155, Ala154, Ser153, Val113, Ala186, Asn204, Val151, Val188, Glu203, Phe183, Ala208, Ala202, Met182, Thr201 | Ser37, Ile105, Met66, Asp107, Leu62, His106, Ile108, Ile65, His110, Leu155, Met48, Val51, Lys49, Ala186, Asp53 | |
| 5D5P | Methanococcus maripaludis | Leu87, Lys119, Met116, Asn84, Leu83, Arg121, Ile122, Leu83, Met80 | Lys119, Leu87, Leu83, Ile122, Met80, Arg79 | Arg79, Thr78, Leu83, Asn84, Leu81 | |
| 1BFM | Methanothermus fervidus | Phe67, Lys68, Val64, Lys69, Ile39, Asp38, Lys68, Met35, Glu34, Ile31 | Val64, Ile39, Leu32, Asp38, Phe67, Lys68, Lys69, Glu34, Met35 | Val64, Ile39, Lys68, Lys69, Phe67, Asp38, Met35, Glu34 | |
| Facultative bacteria | 1USH | Escherichia coli | Val32, Asn497, His43, Trp291, Thr518, Glu290, His117, Asp84, Asp41, Asn517, Gln254, Asp255, Val516, Asn116, His252, Ser253, Ile178, Tyr515, His217, Asn229, Tyr221 | Val432, Asn497, Phe520, His43, Asp510, Thr518, Asn517, Gln254, Asp255, Lys265, Val516, Tyr515, His117, Asp84, Ser253, His252, Ile178, Asn116, Asn229, Tyr221, His220 | Ile521, His43, Val432, His117, Trp291, Thr518, Asp84, Lys293, Asn497, Asn116, Glu290, Ile178, Asn517, Val516, Gln254, His252, His289, Tyr515, Ser253, Asp255 |
| 1BLI | Bacillus licheniformis | Phe190, Met197, Leu196, Tyr193, His105, Glu189, His235, Ser334, Tyr56, Lys234, Ala232, Leu335, Cal233, Trp263, Asp231, Asp328, Tyr262 | Val102, Met197, Tyr56, Leu196, Leu230, Asp231, Arg229, Val233, Ala232, Lys234, Glu261, Trp13, Tyr262, Trp263, His327, Asp328, Ser334, Gln333, Leu335 | Tyr56, Leu196, His235, Asp231, Arg229, Val233, Ala232, Tyr193, Lys234, Phe190, Glu261, Trp13, Glu189, Tyr262, His327, Trp263, Asp328, Ser334, Gln333, Ser264, Leu335 | |
| 2DYT | Saccharomyces cerevisiae | Val250, Gln252, Arg251, Asp133, Lys256, Arg254, Glu134, Ile136, Arg255, Asp279, Gln137, Leu258, Glu140, Ile141 | Val250, Asp133, Arg251, Ile136, Glu134, Arg254, Arg255, Gln137, Met139, Asp279, Asp138, Glu140, Ile141, Lys142 | Asp133, Glu134, Arg251, Arg254, Glu257, Gln137, Asp138, Arg255, Leu258, Ser280, Asp279 | |
| 4GW3 | Proteus mirabilis | Phe143, Thr138, Ile139, Tyr24, Ile140, Phe16, Ile255, Met219, Ala135, Gis254, Phe136, Leu222, Leu13, His11, His78, Leu234, Ala153, Ser79, Leu44, Ala46, Gln80, Ala156, Phe47, Val105, Leu157, Leu160, Agr53 | Asp149, Ser144, Ile19, Phe143, Ile140, Asp152, Val20, Ile139, Phe136, Ala153, Phe16, Leu13, Phe47, Ala156, Tyr24, Leu157, Ile255, His254, Leu234, Ser79 | His146, Asp149, Ser144, Phe143, Ile140, Ile139, Phe136, Asp152, Ile19, Val20, Ala153, Phe16, Ile255, Tyr24, Leu13, Ala156, Phe47, Leu157, Leu234, His254, Ala46, Ser79, Leu160, Gln80 | |
| 1EFP | Paracoccus denitrificans | Ala155, Asp153, Ala154, Phe152, Lys21, Ser151, Asp17, Thr149, Lys272, Glu251, Ala150, Arg194, Pro250, Ser192, Ala249, Ala193, Thr191, Val248 | Asp153, Ala154, Phe152, Lys21, Ser151, Asp17, Thr149, Lys272, Glu251, Ala150, Arg194, Pro250, Ser192, Ala249, Ala193, Thr191, Val248 | Asp270, Ser271, Lys272, Val196, Val248, Ala249, Glu251, Pro250, Ser151, Ala150, Leu190, Thr191, Arg194, Ala193, Thr149, Glu189, Phe152, Ser192, Asp153, Lys21, Ala154 | |
| Degrading Bacteria Type | PDB ID | Novel Degrading Enzyme Number | Hydrophobic Amino Acid Residues (Before Modification) | Hydrophilic Amino Acid Residues (After Modification) |
|---|---|---|---|---|
| Aerobic bacteria | 1GYC | 1-1 | Ala325 | Asp |
| 1-2 | Pro385 | Glu | ||
| 1-3 | Pro34 | His | ||
| Leu232 | Lys | |||
| 1-4 | Pro123 | Glu | ||
| Leu232 | Glu | |||
| Val297 | His | |||
| 1-5 | Val187 | Cys | ||
| Ile298 | Gln | |||
| Val297 | Asp | |||
| 1-6 | Val187 | Gln | ||
| Ile298 | Gln | |||
| Val297 | Gln | |||
| 1-7 | Pro32 | Ser | ||
| Tyr152 | Thr | |||
| Ala386 | Lys | |||
| 1ARP | 2-1 | Ile100 | Cys | |
| Phe114 | Asn | |||
| 2-2 | Ile86 | Cys | ||
| Val112 | His | |||
| 2-3 | Leu88 | Asn | ||
| Ile108 | Thr | |||
| 2-4 | Leu88 | Gln | ||
| Ile108 | Cys | |||
| 2-5 | Leu88 | Ser | ||
| Ile108 | His | |||
| 2-6 | Leu88 | Lys | ||
| Ile108 | Asn | |||
| 2-7 | Leu88 | Arg | ||
| Ile108 | Asp | |||
| 2-8 | Leu88 | Cys | ||
| Ile108 | Thr | |||
| 2-9 | Leu88 | Cys | ||
| Phe114 | Thr | |||
| Ile100 | Thr | |||
| 1YZP | 3-1 | Ile91 | Lys | |
| Phe93 | Lys | |||
| Ile100 | Lys | |||
| 3-2 | Ile91 | Asp | ||
| Phe93 | Asp | |||
| Ile100 | Asp | |||
| 3-3 | Ile91 | Glu | ||
| Phe93 | Glu | |||
| Ile100 | Glu | |||
| 3-4 | Phe70 | Lys | ||
| Ile91 | Lys | |||
| Ile100 | Lys | |||
| 3-5 | Phe70 | Glu | ||
| Ile91 | Glu | |||
| Ile100 | Glu | |||
| 3-6 | Ile91 | Arg | ||
| Phe93 | Arg | |||
| Ile100 | Arg | |||
| 3-7 | Ile91 | Asp | ||
| Phe93 | Asp | |||
| Ile100 | Asp | |||
| 3-8 | Phe70 | Arg | ||
| Ile91 | Arg | |||
| Ile100 | Arg | |||
| 3-9 | Phe70 | Asp | ||
| Ile91 | Asp | |||
| Ile100 | Asp | |||
| 1EI5 | 4-1 | Ala290 | Lys | |
| Phe373 | Lys | |||
| Val392 | Lys | |||
| 4-2 | Ala290 | Asn | ||
| Phe373 | Asn | |||
| Val392 | Asn | |||
| 4-3 | Ala290 | Lys | ||
| Phe373 | Lys | |||
| 4-4 | Ala290 | Asp | ||
| Phe373 | Asn | |||
| 4DTE | 5-1 | Phe181 | Cys | |
| Val184 | Cys | |||
| Pro349 | His | |||
| 5-2 | Phe181 | Cys | ||
| Val184 | Ser | |||
| Pro349 | Lys | |||
| Anaerobic bacteria | 3NQA | 6-1 | Val155 | Lys |
| Ile200 | Lys | |||
| Val201 | Lys | |||
| 6-2 | Val155 | Asp | ||
| Ile200 | Asp | |||
| Val201 | Asp | |||
| 6-3 | Val155 | Asp | ||
| Ile200 | Lys | |||
| Val201 | Lys | |||
| 6-4 | Val155 | Asp | ||
| Ile200 | Asp | |||
| Val201 | Lys | |||
| 6-5 | Val155 | Asp | ||
| Ile200 | Asp | |||
| Val201 | Arg | |||
| 6-6 | Val155 | Arg | ||
| Ile200 | Lys | |||
| Val201 | Asp | |||
| 6-7 | Val155 | Asn | ||
| Ile200 | Asn | |||
| Val201 | Lys | |||
| 6-8 | Val155 | Asn | ||
| Ile200 | Asp | |||
| Val201 | Lys | |||
| 6-9 | Val155 | Asp | ||
| Ile200 | Asn | |||
| Val201 | Lys | |||
| 6-10 | Val182 | Lys | ||
| Ile200 | Lys | |||
| Val201 | Lys | |||
| 1E1D | 7-1 | Trp292 | Lys | |
| Trp293 | Lys | |||
| Tyr493 | Lys | |||
| 7-2 | Trp292 | Arg | ||
| Trp293 | Lys | |||
| Tyr493 | Lys | |||
| 7-3 | Trp292 | Arg | ||
| Trp293 | Lys | |||
| Tyr493 | Arg | |||
| 7-4 | Trp292 | Arg | ||
| Trp293 | Arg | |||
| Tyr493 | Arg | |||
| 7-5 | Trp292 | Arg | ||
| Trp293 | Arg | |||
| Tyr493 | Lys | |||
| 7-6 | Trp292 | Arg | ||
| Trp293 | Asp | |||
| Tyr493 | Lys | |||
| 7-7 | Trp292 | Asp | ||
| Trp293 | Asp | |||
| Tyr493 | Lys | |||
| 7-8 | Trp292 | Asp | ||
| Trp293 | Asp | |||
| Tyr493 | Asp | |||
| 7-9 | Trp292 | Lys | ||
| Trp293 | Asp | |||
| Tyr493 | Asp | |||
| 7-10 | Trp292 | Lys | ||
| Trp293 | Lys | |||
| Tyr493 | Asp | |||
| 3EZX | 8-1 | Leu155 | Arg | |
| Ala186 | Lys | |||
| Ala202 | Asp | |||
| 8-2 | Leu155 | Asn | ||
| Ala186 | Lys | |||
| Ala202 | Asp | |||
| 8-3 | Leu155 | Asn | ||
| Ala186 | Lys | |||
| Ala202 | Asn | |||
| 8-4 | Leu155 | Arg | ||
| Ala186 | Lys | |||
| Ala202 | Asn | |||
| 5D5P | 9-1 | Met80 | Arg | |
| Leu83 | Arg | |||
| Leu87 | Arg | |||
| 9-2 | Met80 | Asp | ||
| Leu83 | Lys | |||
| Leu87 | Lys | |||
| 9-3 | Met80 | Arg | ||
| Leu83 | Lys | |||
| Leu87 | Glu | |||
| 9-4 | Met80 | Thr | ||
| Leu83 | Arg | |||
| Leu87 | Arg | |||
| 1BFM | 10-1 | Met35 | Glu | |
| Ile39 | Gln | |||
| Phe67 | Glu | |||
| Facultative bacteria | 1USH | 11-1 | Val432 | Arg |
| Tyr515 | Arg | |||
| Val516 | Arg | |||
| 11-2 | Val432 | Lys | ||
| Tyr515 | Lys | |||
| Val516 | Lys | |||
| 11-3 | Val432 | Arg | ||
| Tyr515 | Lys | |||
| Val516 | Lys | |||
| 11-4 | Val432 | Arg | ||
| Tyr515 | Arg | |||
| Val516 | Lys | |||
| 11-5 | Val432 | Arg | ||
| Tyr515 | Lys | |||
| Val516 | Arg | |||
| 11-6 | Val432 | Lys | ||
| Tyr515 | Lys | |||
| Val516 | Arg | |||
| 11-7 | Val432 | Lys | ||
| Tyr515 | Arg | |||
| Val516 | Lys | |||
| 11-8 | Val432 | Lys | ||
| Tyr515 | Arg | |||
| Val516 | Arg | |||
| 11-9 | Val432 | Asp | ||
| Tyr515 | Asp | |||
| Val516 | Asp | |||
| 11-10 | Val432 | Arg | ||
| Tyr515 | Asp | |||
| Val516 | Asp | |||
| 1BLI | 12-1 | Leu196 | Arg | |
| Ala232 | Asp | |||
| Val233 | Arg | |||
| 2DYT | 13-1 | Ile136 | Arg | |
| Met139 | Arg | |||
| Leu258 | Arg | |||
| 13-2 | Ile136 | Glu | ||
| Met139 | Glu | |||
| Leu258 | Glu | |||
| 13-3 | Ile136 | Lys | ||
| Met139 | Lys | |||
| Leu258 | Lys | |||
| 13-4 | Ile136 | Arg | ||
| Met139 | Arg | |||
| Leu258 | Lys | |||
| 13-5 | Ile136 | Arg | ||
| Met139 | Lys | |||
| Leu258 | Lys | |||
| 13-6 | Ile136 | Lys | ||
| Met139 | Arg | |||
| Leu258 | Arg | |||
| 13-7 | Ile136 | Glu | ||
| Met139 | Arg | |||
| Leu258 | Arg | |||
| 13-8 | Ile136 | Glu | ||
| Met139 | Glu | |||
| Leu258 | Arg | |||
| 13-9 | Ile136 | Arg | ||
| Met139 | Glu | |||
| Leu258 | Lys | |||
| 4GW3 | 14-1 | Phe136 | Arg | |
| Ile139 | Arg | |||
| Ile140 | Arg | |||
| 14-2 | Phe136 | Lys | ||
| Ile139 | Lys | |||
| Ile140 | Lys | |||
| 14-3 | Phe136 | Arg | ||
| Ile139 | Lys | |||
| Ile140 | Lys | |||
| 14-4 | Phe136 | Arg | ||
| Ile139 | Lys | |||
| Ile140 | Arg | |||
| 14-5 | Phe136 | Lys | ||
| Ile139 | Lys | |||
| Ile140 | Arg | |||
| 14-6 | Phe136 | Asp | ||
| Ile139 | Lys | |||
| Ile140 | Arg | |||
| 14-7 | Phe136 | Arg | ||
| Ile139 | Arg | |||
| Ile140 | Asp | |||
| 14-8 | Phe136 | Arg | ||
| Ile139 | Asp | |||
| Ile140 | Arg | |||
| 1EFP | 15-1 | Ala150 | Arg | |
| Phe152 | Arg | |||
| Pro250 | Arg | |||
| 15-2 | Ala150 | Lys | ||
| Phe152 | Lys | |||
| Pro250 | Lys | |||
| 15-3 | Ala150 | Arg | ||
| Phe152 | Lys | |||
| Pro250 | Lys | |||
| 15-4 | Ala150 | Arg | ||
| Phe152 | Arg | |||
| Pro250 | Lys | |||
| 15-5 | Ala150 | Asp | ||
| Phe152 | Arg | |||
| Pro250 | Arg | |||
| 15-6 | Ala150 | Asp | ||
| Phe152 | Asp | |||
| Pro250 | Arg | |||
| 15-7 | Ala150 | Asp | ||
| Phe152 | Asp | |||
| Pro250 | Asp | |||
| 15-8 | Ala150 | Arg | ||
| Phe152 | Lys | |||
| Pro250 | Asp |
| Degrading Bacteria Type | PDB ID | Novel Degrading Enzyme | With the CIP | With the NOR | With the OFL | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Modification Scoring Function | Modified Scoring Function | Change Rate (%) | Pre-Modification Scoring Function | Modified Scoring Function | Change Rate (%) | Pre-Modification Scoring Function | Modified Scoring Function | Change Rate (%) | |||
| Aerobic bacteria | 1GYC | 1-1 | 3.27 | 3.97 | 21.41 | 2.13 | 4.48 | 110.33 | 2.20 | 2.43 | 10.45 |
| 1-2 | 4.31 | 31.80 | 2.80 | 31.46 | 3.44 | 56.36 | |||||
| 1-3 | 4.49 | 37.31 | 2.79 | 30.99 | 3.32 | 50.91 | |||||
| 1-4 | 4.35 | 33.03 | 2.58 | 21.13 | 3.07 | 39.55 | |||||
| 1-5 | 4.00 | 22.32 | 4.38 | 105.63 | 3.83 | 74.09 | |||||
| 1-6 | 3.60 | 10.09 | 3.84 | 80.28 | 3.76 | 70.91 | |||||
| 1-7 | 4.13 | 26.30 | 2.88 | 35.21 | 2.70 | 22.73 | |||||
| 1ARP | 2-1 | 3.36 | 5.30 | 57.74 | 3.01 | 5.36 | 78.07 | 3.84 | 4.78 | 24.48 | |
| 2-2 | 6.16 | 83.33 | 7.90 | 162.46 | 4.95 | 28.91 | |||||
| 2-3 | 6.37 | 89.58 | 7.18 | 138.54 | 5.15 | 34.11 | |||||
| 2-4 | 6.29 | 87.20 | 7.05 | 134.22 | 5.07 | 32.03 | |||||
| 2-5 | 6.12 | 82.14 | 8.15 | 170.76 | 5.45 | 41.93 | |||||
| 2-6 | 7.05 | 109.82 | 7.74 | 157.14 | 5.66 | 47.40 | |||||
| 2-7 | 6.37 | 89.58 | 7.13 | 136.88 | 6.06 | 57.81 | |||||
| 2-8 | 6.07 | 80.65 | 7.49 | 148.84 | 5.11 | 33.07 | |||||
| 2-9 | 6.26 | 86.31 | 6.39 | 112.29 | 6.38 | 66.15 | |||||
| 1YZP | 3-1 | 3.64 | 5.94 | 63.19 | 2.66 | 7.29 | 174.06 | 2.95 | 5.87 | 98.98 | |
| 3-2 | 5.85 | 60.71 | 6.40 | 140.60 | 6.59 | 123.39 | |||||
| 3-3 | 6.86 | 88.46 | 5.64 | 112.03 | 6.54 | 121.69 | |||||
| 3-4 | 5.81 | 59.62 | 7.31 | 174.81 | 5.74 | 94.58 | |||||
| 3-5 | 5.77 | 58.52 | 6.09 | 128.95 | 6.62 | 124.41 | |||||
| 3-6 | 5.75 | 57.97 | 7.41 | 178.57 | 5.45 | 84.75 | |||||
| 3-7 | 5.85 | 60.71 | 6.40 | 140.60 | 6.59 | 123.39 | |||||
| 3-8 | 5.91 | 62.36 | 7.40 | 178.20 | 5.77 | 95.59 | |||||
| 3-9 | 6.01 | 65.11 | |||||||||
| 1EI5 | 4-1 | 5.08 | 6.43 | 26.57 | 3.74 | 6.07 | 62.30 | 4.44 | 5.55 | 25.00 | |
| 4-2 | 5.77 | 13.58 | 5.23 | 39.84 | 5.16 | 16.22 | |||||
| 4-3 | 5.67 | 11.61 | 4.74 | 26.74 | 4.90 | 10.36 | |||||
| 4-4 | 5.60 | 10.24 | 4.71 | 25.94 | 4.93 | 11.04 | |||||
| 4DTE | 5-1 | 3.70 | 4.07 | 10.00 | 3.23 | 3.72 | 15.17 | 3.66 | 4.21 | 15.03 | |
| 5-2 | 6.00 | 62.16 | 5.23 | 61.92 | 5.77 | 57.65 | |||||
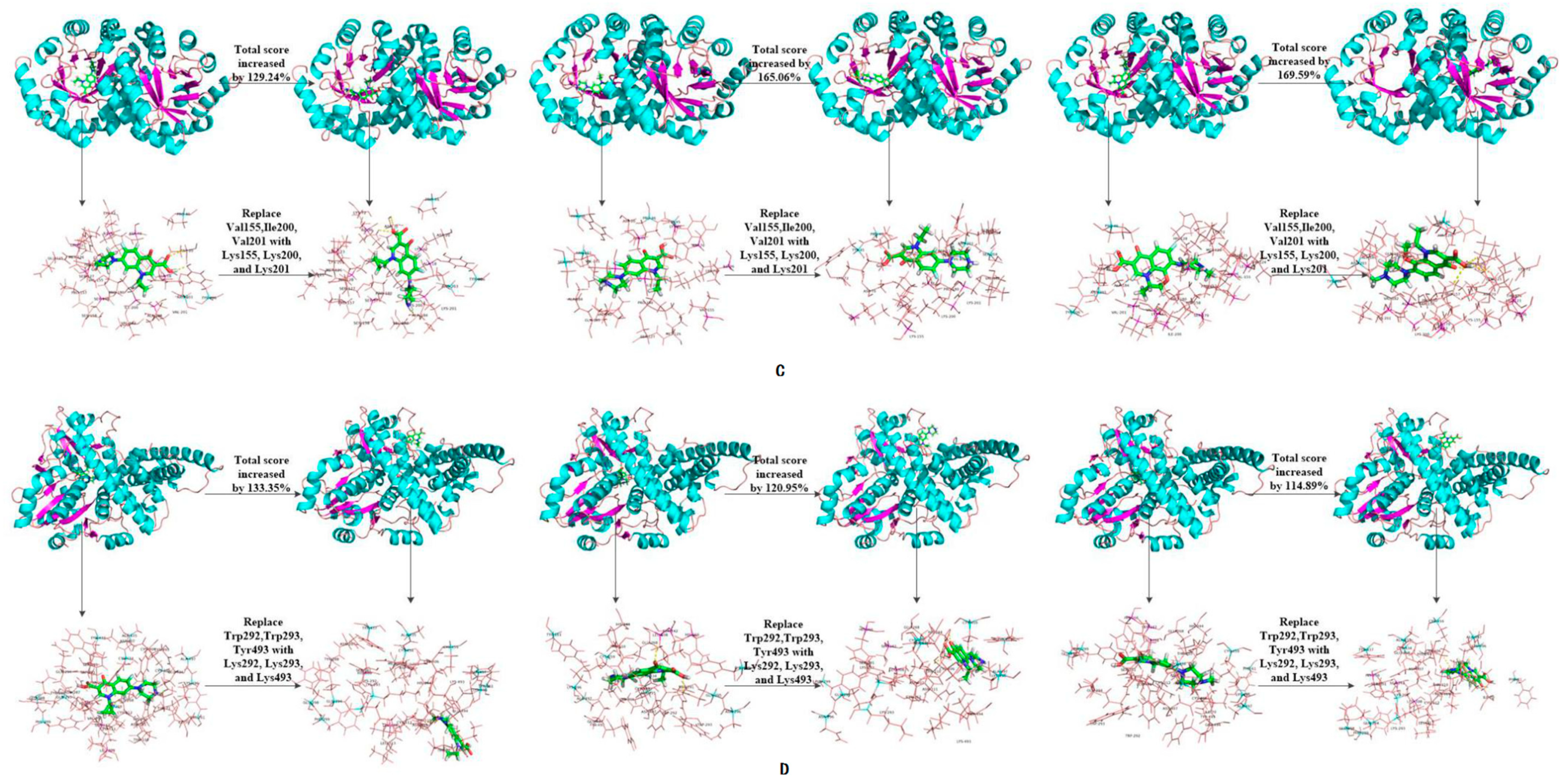
| Anaerobic bacteria | 3NQA | 6-1 | 1.71 | 3.92 | 129.24 | 2.69 | 7.13 | 165.06 | −3.42 | 2.38 | 169.59 |
| 6-2 | 2.71 | 58.48 | 3.46 | 28.62 | −0.35 | 89.77 | |||||
| 6-3 | 4.27 | 149.71 | 3.76 | 39.78 | 0.09 | 102.63 | |||||
| 6-4 | 2.42 | 41.52 | 4.00 | 48.70 | −2.62 | 23.39 | |||||
| 6-5 | 2.03 | 18.71 | 4.55 | 69.14 | −2.19 | 35.96 | |||||
| 6-6 | 2.39 | 39.77 | 5.09 | 89.22 | −1.16 | 66.08 | |||||
| 6-7 | 3.94 | 130.41 | 3.74 | 39.03 | −1.34 | 60.82 | |||||
| 6-8 | 4.24 | 147.95 | 3.73 | 38.66 | −2.86 | 16.37 | |||||
| 6-9 | 3.12 | 82.46 | 5.46 | 102.97 | 2.75 | 180.41 | |||||
| 6-10 | 3.65 | 113.45 | 4.84 | 79.93 | 2.68 | 178.36 | |||||
| 1E1D | 7-1 | −18.08 | 6.03 | 133.35 | 4.11 | 120.95 | −28.47 | 4.24 | 114.89 | ||
| 7-2 | 5.47 | 130.25 | 3.13 | 115.95 | 4.86 | 117.07 | |||||
| 7-3 | 5.27 | 129.15 | 4.01 | 120.44 | 3.12 | 110.96 | |||||
| 7-4 | 5.18 | 128.65 | 3.86 | 119.67 | 4.23 | 114.86 | |||||
| 7-5 | 5.09 | 128.15 | 3.61 | 118.40 | 4.39 | 115.42 | |||||
| 7-6 | 4.62 | 125.55 | 3.04 | 115.49 | 4.33 | 115.21 | |||||
| 7-7 | 5.61 | 131.03 | 2.85 | 114.53 | 4.23 | 114.86 | |||||
| 7-8 | 5.28 | 129.20 | 3.90 | 119.88 | 4.30 | 115.10 | |||||
| 7-9 | 5.03 | 127.82 | 2.84 | 114.48 | 4.18 | 114.68 | |||||
| 7-10 | 5.25 | 129.04 | 3.68 | 118.76 | 2.97 | 110.43 | |||||
| 3EZX | 8-1 | 4.09 | 6.63 | 62.10 | 4.00 | 4.51 | 12.75 | 4.60 | 5.24 | 13.91 | |
| 8-2 | 6.62 | 61.86 | 5.93 | 48.25 | 5.97 | 29.78 | |||||
| 8-3 | 4.62 | 12.96 | 6.33 | 58.25 | 5.34 | 16.09 | |||||
| 8-4 | 4.56 | 11.49 | 4.91 | 22.75 | 5.39 | 17.17 | |||||
| 5D5P | 9-1 | 4.58 | 5.11 | 11.57 | 2.74 | 4.28 | 56.20 | 4.17 | 5.22 | 25.18 | |
| 9-2 | 6.92 | 51.09 | 3.10 | 13.14 | 5.47 | 31.18 | |||||
| 9-3 | 6.02 | 31.44 | 4.28 | 56.20 | 5.78 | 38.61 | |||||
| 9-4 | 5.04 | 10.04 | 5.23 | 90.88 | 4.90 | 17.51 | |||||
| 1BFM | 10-1 | 3.12 | 4.02 | 28.85 | 3.46 | 4.38 | 26.59 | 3.05 | 4.29 | 40.66 | |
| Facultative bacteria | 1USH | 11-1 | 3.78 | 5.97 | 57.94 | 3.08 | 4.53 | 47.08 | 3.37 | 5.21 | 54.60 |
| 11-2 | 5.57 | 47.35 | 4.01 | 30.19 | 4.45 | 32.05 | |||||
| 11-3 | 5.65 | 49.47 | 3.87 | 25.65 | 4.35 | 29.08 | |||||
| 11-4 | 6.11 | 61.64 | 4.27 | 38.64 | 3.72 | 10.39 | |||||
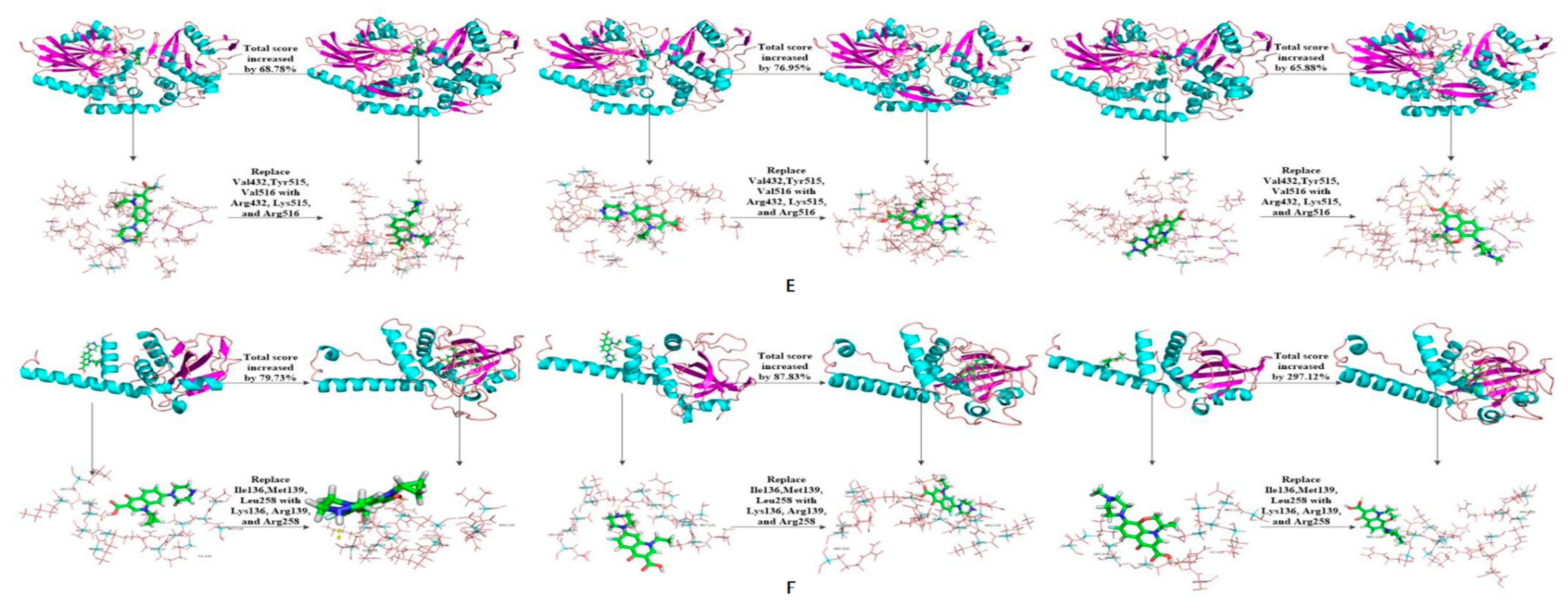
| 11-5 | 6.38 | 68.78 | 5.45 | 76.95 | 5.59 | 65.88 | |||||
| 11-6 | 5.89 | 55.82 | 5.62 | 82.47 | 5.42 | 60.83 | |||||
| 11-7 | 5.89 | 55.82 | 4.46 | 44.81 | 5.25 | 55.79 | |||||
| 11-8 | 5.63 | 48.94 | 4.88 | 58.44 | 5.30 | 57.27 | |||||
| 11-9 | 4.40 | 16.40 | 5.24 | 70.13 | 4.77 | 41.54 | |||||
| 11-10 | 4.74 | 25.40 | 6.02 | 95.45 | 5.26 | 56.08 | |||||
| 1BLI | 12-1 | 3.51 | 5.65 | 60.97 | 3.49 | 4.42 | 26.65 | 4.26 | 4.73 | 11.03 | |
| 2DYT | 13-1 | 2.96 | 3.53 | 19.26 | 2.63 | 4.01 | 52.47 | 1.39 | 3.03 | 117.99 | |
| 13-2 | 4.71 | 59.12 | 3.58 | 36.12 | 3.71 | 166.91 | |||||
| 13-3 | 5.95 | 101.01 | 3.37 | 28.14 | 4.49 | 223.02 | |||||
| 13-4 | 6.77 | 128.72 | 4.34 | 65.02 | 4.17 | 200.00 | |||||
| 13-5 | 4.30 | 45.27 | 3.74 | 42.21 | 3.92 | 182.01 | |||||
| 13-6 | 5.32 | 79.73 | 4.94 | 87.83 | 5.52 | 297.12 | |||||
| 13-7 | 4.80 | 62.16 | 4.00 | 52.09 | 3.11 | 123.74 | |||||
| 13-8 | 4.49 | 51.69 | 4.81 | 82.89 | 3.37 | 142.45 | |||||
| 13-9 | 4.69 | 58.45 | 3.00 | 14.07 | 3.14 | 125.90 | |||||
| 4GW3 | 14-1 | 3.60 | 4.70 | 30.56 | 2.61 | 5.56 | 113.03 | 3.66 | 5.36 | 46.45 | |
| 14-2 | 4.36 | 21.11 | 3.93 | 50.57 | 5.22 | 42.62 | |||||
| 14-3 | 4.53 | 25.83 | 4.77 | 82.76 | 6.41 | 75.14 | |||||
| 14-4 | 4.27 | 18.61 | 4.82 | 84.67 | 6.05 | 65.30 | |||||
| 14-5 | 4.00 | 11.11 | 4.54 | 73.95 | 5.09 | 39.07 | |||||
| 14-6 | 5.54 | 53.89 | 5.21 | 99.62 | 4.99 | 36.34 | |||||
| 14-7 | 5.28 | 46.67 | 4.87 | 86.59 | 5.27 | 43.99 | |||||
| 14-8 | 5.30 | 47.22 | 4.55 | 74.33 | 5.04 | 37.70 | |||||
| 1EFP | 15-1 | 4.48 | 5.59 | 24.78 | 3.12 | 4.36 | 39.74 | 3.32 | 5.06 | 52.41 | |
| 15-2 | 5.89 | 31.47 | 3.70 | 18.59 | 5.22 | 57.23 | |||||
| 15-3 | 5.62 | 25.45 | 5.36 | 71.79 | 3.81 | 14.76 | |||||
| 15-4 | 5.63 | 25.67 | 4.16 | 33.33 | 4.76 | 43.37 | |||||
| 15-5 | 5.70 | 27.23 | 4.61 | 47.76 | 4.94 | 48.80 | |||||
| 15-6 | 5.30 | 18.30 | 4.00 | 28.21 | 5.24 | 57.83 | |||||
| 15-7 | 4.93 | 10.04 | 4.73 | 51.60 | 4.77 | 43.67 | |||||
| 15-8 | 5.26 | 17.41 | 4.87 | 56.09 | 4.95 | 49.10 | |||||
| Molecular Target | Enzyme | Ereactant/(a.u.) | ETS/(a.u.) | ΔE/(a.u.) | ΔE/(kcal·mol−1) |
|---|---|---|---|---|---|
| CIP | 1YZP | −1148.37 | −1224.11 | −75.74 | −47,527.61 |
| 3-6 | −1148.37 | −1224.11 | −75.74 | −47,527.61 | |
| 4DTE | −1148.37 | −1224.13 | −75.75 | −47,533.88 | |
| 5-2 | −1148.37 | −1224.05 | −75.68 | −47,489.96 | |
| 3NQA | −1148.37 | −1224.12 | −75.75 | −47,533.88 | |
| 6-1 | −1148.37 | −1223.93 | −75.56 | −47,414.66 | |
| 1E1D | −1148.37 | −1224.03 | −75.66 | −47,477.41 | |
| 7-1 | −1148.37 | −1224.09 | −75.72 | −47,515.06 | |
| 1USH | −1148.37 | −1224.11 | −75.74 | −47,527.61 | |
| 11-5 | −1148.37 | −1224.11 | −75.74 | −47,527.61 | |
| 2DYT | −1148.37 | −1223.98 | −75.61 | −47,446.03 | |
| 13-6 | −1148.37 | −1224.11 | −75.74 | −47,527.61 | |
| NOR | 1YZP | −1110.31 | −1186.01 | −75.70 | −47,502.51 |
| 3-6 | −1110.31 | −1186.02 | −75.71 | −47,508.78 | |
| 4DTE | −1110.31 | −1186.04 | −75.73 | −47,521.33 | |
| 5-2 | −1110.31 | −1185.98 | −75.67 | −47,483.68 | |
| 3NQA | −1110.31 | −1185.96 | −75.65 | −47,471.13 | |
| 6-1 | −1110.31 | −1185.87 | −75.56 | −47,414.66 | |
| 1E1D | −1110.31 | −1185.96 | −75.65 | −47,471.13 | |
| 7-1 | −1110.31 | −1186.03 | −75.72 | −47,515.06 | |
| 1USH | −1110.31 | −1185.85 | −75.53 | −47,395.83 | |
| 11-5 | −1110.31 | −1186.05 | −75.74 | −47,527.61 | |
| 2DYT | −1110.31 | −1186.04 | −75.73 | −47,521.33 | |
| 13-6 | −1110.31 | −1186.04 | −75.73 | −47,521.33 | |
| OFL | 1YZP | −1262.93 | −1338.58 | −75.65 | −47,471.13 |
| 3-6 | −1262.93 | −1338.60 | −75.67 | −47,483.68 | |
| 4DTE | −1262.93 | −1338.67 | −75.74 | −47,527.61 | |
| 5-2 | −1262.93 | −1338.67 | −75.74 | −47,527.61 | |
| 3NQA | −1262.93 | −1224.12 | 38.81 | 24,353.66 | |
| 6-1 | −1262.93 | −1338.66 | −75.73 | −47,521.33 | |
| 1E1D | −1262.93 | −1338.66 | −75.73 | −47,521.33 | |
| 7-1 | −1262.93 | −1338.67 | −75.74 | −47,527.61 | |
| 1USH | −1262.93 | −1338.58 | −75.65 | −47,471.13 | |
| 11-5 | −1262.93 | −1338.67 | −75.74 | −47,527.61 | |
| 2DYT | −1262.93 | −1338.67 | −75.74 | −47,527.61 | |
| 13-6 | −1262.93 | −1338.55 | −75.62 | −47,452.31 |
| Target Fluoroquinolone Molecules | Novel Degradation Enzyme Number | Degree of Reduction (%) |
|---|---|---|
| CIP | 7-1 | 6.60 |
| 13-6 | 6.60 | |
| NOR | 3-6 | 6.82 |
| 7-1 | 6.82 | |
| 11-5 | 6.82 | |
| OFL | 3-6 | 5.99 |
| 7-1 | 6.00 | |
| 11-5 | 6.00 |
| PDB ID | Scoring Function | Novel Degrading Enzyme Number | Scoring Function | Amplitude of Variation |
|---|---|---|---|---|
| 1YZP | 2.09 | 3-6 | 2.07 | −0.96% |
| 4DTE | 2.06 | 5-2 | 2.21 | 7.28% |
| 3NQA | 2.40 | 6-1 | 3.01 | 25.42% |
| 1E1D | 2.38 | 7-1 | 2.67 | 12.1% |
| 1USH | 2.36 | 11-5 | 3.77 | 59.75% |
| 2DYT | 2.57 | 13-6 | 2.13 | −17.12% |
| Target Molecular | Degrading Enzyme | Interaction Type | Amino Acid Residue |
|---|---|---|---|
| CIP | 1YZP | Van der Waals | Glu143, Pro144, Gln145, Lys180, Thr219, Gly220 |
| Electrostatic | Arg42, Ser78, Asn81, Val181 | ||
| Carbon hydrogen bond interaction | Ser78 | ||
| Conventional hydrogen bond interaction | Arg42 | ||
| Alkyl interaction | Pro144 | ||
| π-Alkyl interaction | Val181 | ||
| Amide-π stacked interaction | Lys180, Val181 | ||
| 3-6 | Van der Waals | Glu35, His38, Glu39, Ile41, Ala176, Ala178 | |
| Electrostatic | Arg42, Gly82, Ser172, His173, Val175, Arg177, Asp179, Lys180 | ||
| Carbon hydrogen bond interaction | His38, Ser172, Val175, Asp179 | ||
| Conventional hydrogen bond interaction | Asp179, Lys180 | ||
| Halogen (fluorine) interaction | His173 | ||
| Alkyl interaction | Arg177 | ||
| π-Alkyl interaction | His38, Arg42, Ala176 | ||
| π Interaction pair | Arg42 | ||
| π-Sigma interaction | Arg42 | ||
| 4DTE | Van der W#aals | Pro219, Gly360, Ile225, Ser224, His356 | |
| Electrostatic | Ser226, Gln355, Arg255, Glu223, Glu217, Lys357, His252 | ||
| Carbon hydrogen bond interaction | Glu217, Gly360 | ||
| Conventional hydrogen bond interaction | Ser226, Glu223 | ||
| Halogen (fluorine) interaction | Ser226, Gln355 | ||
| Alkyl interaction | Arg255 | ||
| π-Alkyl interaction | Lys357 | ||
| π-Cation interaction | Arg255 | ||
| 5-2 | Van der Waals | Phe15, Leu27, Ala28, Ser30, Pro31, Val83, Leu161, Phe163, Ile225, Val316, Lys317, Leu318, Phe351 | |
| Electrostatic | Leu29, Phe352, Ile354 | ||
| Carbon hydrogen bond interaction | Leu27, Pro31 | ||
| Alkyl interaction | Ala28, Leu161, Leu318 | ||
| π-Donor hydrogen bond interaction | Ile354 | ||
| π-Alkyl interaction | Leu161 | ||
| 3NQA | Van der Waals | Val151, Asn111, Ala113 | |
| Electrostatic | Asn153, Leu149, Leu110, Gly150, Arg107, Val120, Lys152, Glu119, Glu114 | ||
| Carbon hydrogen bond interaction | Val120, Gly150, Leu149 | ||
| Halogen (fluorine) interaction | Leu110 | ||
| π-Alkyl interaction | Leu110 | ||
| π-Cation interaction | Arg107 | ||
| π-Donor hydrogen bond interaction | Arg107 | ||
| 6-1 | Van der Waals | Leu49, Ser50, Gly52, Ala86, Ala90 | |
| Electrostatic | Val48, Leu49, Gly52, Met53, Asp54, Lys89 | ||
| Carbon hydrogen bond interaction | Val48, Leu49 | ||
| Alkyl interaction | Met53, Lys89, Ala90 | ||
| π-Alkyl interaction | Ala90 | ||
| 1E1D | Van der Waals | Asp483, Ile481, Val509, Val486, Met549 | |
| Electrostatic | Asp480, Asn482, Asn535, Lys537, Lys510, Gly511, Arg513, Lys553, Ser487, Tyr488, Arg399 | ||
| Carbon hydrogen bond interaction | Asp480, Asn482 | ||
| Conventional hydrogen bond interaction | Asp480, Lys537, Arg513 | ||
| Halogen (fluorine) interaction | Asn535 | ||
| π-Alkyl interaction | Tyr488 | ||
| 7-1 | Van der Waals | Lys293, Lys411, Ser414, Tyr439, Leu442 | |
| Electrostatic | Ser291, Gln294, Asp407, Gly408, Arg409, Thr417, Tyr437, Arg438 | ||
| Carbon hydrogen bond interaction | Ser291, Asp407, Gly408 | ||
| Halogen (fluorine) interaction | Gly294, Asp407 | ||
| π-Interaction pair | Arg438 | ||
| π-Cation interaction | Arg438 | ||
| 1USH | Van der Waals | Tyr221, Glu225, Asn511, Lys512, Pro513, Tyr515, Val516 | |
| Electrostatic | His226, Gly227, Ser228, Asn229, Asp510, Gly514, Asn517 | ||
| Carbon hydrogen bond interaction | His226, Ser228, Asn229, Lys512, Tyr515 | ||
| Coventional hydrogen bond interaction | His226, Ser228 | ||
| Halogen (fluorine) interaction | Asp510 | ||
| π-Alkyl interaction | Tyr515 | ||
| π-π T-shaped interaction | Tyr515 | ||
| π-Lone pair interaction | Asp510 | ||
| 11-5 | Van der Waals | His220, Tyr221, Ser253, Asp510, Asn517 | |
| Electrostatic | Asp222, Glu225, His226, Gly227, Ser228, Asn229, Asn255, Lys265, Pro513, Gly514, Lys515, Arg516 | ||
| Carbon hydrogen bond interaction | His220, Asp255, Lys515 | ||
| Coventional hydrogen bond interaction | Ser228, Asp255, Gly514, Lys515 | ||
| Halogen (fluorine) interaction | Glu225 | ||
| Alkyl interaction | Lys515 | ||
| π-π Stacked interaction | Tyr221 | ||
| 2DYT | Van der Waals | Ile129, Ile132, Leu135, Leu243, Val289, Phe293, Ser296, Thr306 | |
| Electrostatic | Ile288, Lys292, His303 | ||
| Conventional hydrogen bond interaction | Ile288 | ||
| Halogen (fluorine) interaction | His303 | ||
| Alkyl interaction | Ile132 | ||
| π-Alkyl interaction | Lys292 | ||
| π-Sigma interaction | Thr295 | ||
| Amide-π stacked interaction | Thr295, Ser296 | ||
| 13-6 | Van der Waals | Val86, Glu87, Pro89, Asp90, Val91, Glu152, Phe153, Ala155, Gly158, Leu159 | |
| Electrostatic | Lys78, Gly88, Asn154, Pro299, Ser300, Ile301, Gln302, His303 | ||
| Carbon hydrogen bond interaction | Asn154, Gln302 | ||
| Conventional hydrogen bond interaction | Asn154, His303 | ||
| Alkyl interaction | Val86, Pro89 | ||
| π-π T-shaped interaction | His303 | ||
| NOR | 1YZP | Van der Waals | His46, Ile141, Pro144, Gln145, Asn218, Gly220, Leu239 |
| Electrostatic | Arg42, Ser78, Ala79, Asn80, Asn81, Pro142, Glu143, Lys180, Val181, Gln183, Thr219 | ||
| Carbon hydrogen bond interaction | Ala79, Thr219 | ||
| Conventional hydrogen bond interaction | Arg42, Asn81 | ||
| Halogen (fluorine) interaction | Arg42, Ser78, Ala79, Asn81 | ||
| Alkyl interaction | Ala79, Pro144 | ||
| π-Sigma interaction | Val181 | ||
| π-Interaction pair | Arg42 | ||
| 3-6 | Van der Waals | Glu35, Glu39, Ile41, Gly82, Ala176, Phe190 | |
| Electrostatic | His38, Arg42, Ser172, His173, Val175, Arg177, Ala178, Asp179 | ||
| Carbon hydrogen bond interaction | His38, Val175, Asp179 | ||
| Halogen (fluorine) interaction | His173 | ||
| π-Alkyl interaction | Arg42, Ala176 | ||
| π-Interaction pair | Arg42 | ||
| π-Cation interaction | Arg42 | ||
| 4DTE | Van der Waals | Pro219, Gly360, His356, Ser224, Tyr220, Ile225 | |
| Electrostatic | Glu223, Glu217, Ser226, Arg255, Lys357, His252, Gln355 | ||
| Carbon hydrogen bond interaction | Glu223, Glu217, Gly360 | ||
| Conventional hydrogen bond interaction | Glu223, Ser226 | ||
| Halogen (fluorine) interaction | Gln355 | ||
| Alkyl interaction | Arg255 | ||
| π-Alkyl interaction | Lys357 | ||
| π-Cation interaction | Arg255 | ||
| π interaction pair | Arg255 | ||
| 5-2 | Van der Waals | Phe15, Ala28, Ser30, Pro31, Val83, Leu161, Phe163, Val316, Lys317, Leu318, Leu353 | |
| Electrostatic | Leu27, Leu29, His162, Phe352, Ile354 | ||
| Carbon hydrogen bond interaction | Leu29, Pro31, Phe352 | ||
| Alkyl interaction | Leu161, Val316, Leu318 | ||
| π-Sigma interaction | Leu161, Ile354 | ||
| π-Donor hydrogen bond interaction | Ile354 | ||
| 3NQA | Van der Waals | Ala113, Gly150, Val151, Leu149 | |
| Electrostatic | Lys152, Glu119, Leu110, Asn111, Val120, Glu114, Arg107, Asn153 | ||
| Carbon hydrogen bond interaction | Glu119, Gly150 | ||
| Conventional hydrogen bond interaction | Glu114 | ||
| Halogen (fluorine) interaction | Leu110 | ||
| π-Cation interaction | Arg107 | ||
| π-Sigma interaction | Leu110 | ||
| π-Anion interaction | Glu114 | ||
| π-Interaction pair | Arg107 | ||
| 6-1 | Van der Waals | Ser50, Ser52 | |
| Electrostatic | Val48, Leu49, Gly52, Met53, Asp54 | ||
| Carbon hydrogen bond interaction | Val48 | ||
| Halogen (fluorine) interaction | Val48, Leu49 | ||
| π-Donor hydrogen bond interaction | Met53 | ||
| π-Anion interaction | Asp54 | ||
| 1E1D | Van der Waals | Asp483, Val509, Val486, Ile512, Met549 | |
| Electrostatic | Asp480, Lys510, Asn535, Lys553, Lys537, Gly511, Arg513, Asn482, Tyr488, Ser487 | ||
| Carbon hydrogen bond interaction | Asp480, Asn482 | ||
| Coventional hydrogen bond interaction | Asp480, Lys537, Arg513 | ||
| π-Alkyl interaction | Tyr488 | ||
| 7-1 | Van der waals | Lys293, Lys411, Ser414, Leu442 | |
| Electrostatic | Ser291, Gln294, Asp407, Gly408, Arg409, Thr417, Tyr437, Arg438 | ||
| Carbon hydrogen bond interaction | Ser291, Asp407, Gly408, Arg409 | ||
| Halogen (fluorine) interaction | Gln294, Asp407 | ||
| π-Cation interaction | Arg438 | ||
| 1USH | Van der Waals | Gly227, Pro513 | |
| Electrostatic | Ser228, Asn229, Ser253, Asp510, Asn511, Lys512, Gly514, Tyr515, Asn517 | ||
| Carbon hydrogen bond interaction | Asp510, Val516 | ||
| Conventional hydrogen bond interaction | Ser228, Asn517 | ||
| Halogen (fluorine) interaction | Ser228, Lys512 | ||
| π-π T-shaped interaction | Tyr515 | ||
| 11-5 | Van der Waals | Gly227, Val434, Thr501, Asn511, Lys512 | |
| Electrostatic | Ser228, Asn229, Asp510, Pro513, Gly514, Lys515, Arg516, Asn517 | ||
| Carbon hydrogen bond interaction | Asp510 | ||
| Coventional hydrogen bond interaction | Ser228, Arg516, Asn517 | ||
| Halogen (fluorine) interaction | Asn517 | ||
| π-Alkyl interaction | Lys515 | ||
| 2DYT | Van der Waals | Ile129, Asp131, Ile132, Leu135, Ile136, Leu243, Val289, Ser291, Phe293, Ser296, Thr306 | |
| Electrostatic | Ile288, Lys292, Thr295, His303 | ||
| Carbon hydrogen bond interaction | Ile288 | ||
| Halogen (fluorine) interaction | His303 | ||
| Alkyl interaction | Ile132, Leu243, Lys292 | ||
| π-Alkyl interaction | Lys292 | ||
| Amide-π stacked interaction | Thr295, Ser296 | ||
| 13-6 | Van der Waals | Val86, Glu87, Gly88, Pro89, Asp90, Ala155, Lys156, Gly158 | |
| Electrostatic | Asp77, Lys78, Asn154, Gly157, Leu159, Ser300, Gln302, His303 | ||
| Carbon hydrogen bond interaction | Asn154 | ||
| Alkyl interaction | Val86, Pro89 | ||
| π-Alkyl interaction | Val86 | ||
| π-Anion interaction | Asp77 | ||
| OFL | 1YZP | Van der Waals | His46, Ile141, Pro144, Val181, Thr219 |
| Electrostatic | Arg42, Ala79, Asn81, Pro142, Glu143, Glu145, Lys180, Gln183 | ||
| Carbon hydrogen bond interaction | Ala79, Pro142, Glu143, Gln145 | ||
| Coventional hydrogen bond interaction | Asn81 | ||
| Halogen (fluorine) interaction | Asn81 | ||
| Alkyl interaction | Ala79, Pro144 | ||
| 3-6 | Van der Waals | Glu35, Glu39, Ile41, Asn81, Gly82, Pro144, Ala178, Phe190, Leu239 | |
| Electrostatic | His38, Arg42, Ser172, His173, Val175, Arg177, Asp179, Lys180, Val181 | ||
| Carbon hydrogen bond interaction | His38, Arg177 | ||
| Conventional hydrogen bond interaction | Val175 | ||
| Alkyl interaction | Ala178, Lys180, Val181 | ||
| π-Alkyl interaction | His38, Arg42, Ala176 | ||
| π-Interaction pair | Arg42 | ||
| π-Cation interaction | Arg42 | ||
| π-Sigma interaction | Ala176 | ||
| π-π T-shaped interaction | Phe190 | ||
| 4DTE | Van der Waals | Asn201, Tyr220, Pro219, His356 | |
| Electrostatic | Arg255, Glu223, Glu217, Lys357, Ser226, Glu355, Ser24, Ile225 | ||
| Carbon hydrogen bond interaction | Glu223 | ||
| Conventional hydrogen bond interaction | Lys357 | ||
| Alkyl interaction | Pro219 | ||
| π-Anion interaction | Glu217 | ||
| 5-2 | Van der Waals | Phe15, Ala28, Ser30, Pro31, Leu77, Glu80, Val83, Leu161, Phe163, Ile225, Val316, Leu318, His356 | |
| Electrostatic | Leu27, Leu29, Tyr32, Phe352, Ile354 | ||
| Carbon hydrogen bond interaction | Ser30 | ||
| Conventional hydrogen bond interaction | Phe352 | ||
| Halogen (fluorine) interaction | Ile354 | ||
| Alkyl interaction | Ala28, Leu161, Val316, Leu318, Ile354 | ||
| π-Alkyl interaction | Ala28, Pro31, Ile354 | ||
| 3NQA | Van der Waals | Arg118, Glu119, Val120, Glu114, Arg107, Leu149 | |
| Electrostatic | Glu119, Arg118, Ala113 | ||
| Carbon hydrogen bond interaction | Leu110 | ||
| Alkyl interaction | Leu110 | ||
| π-Alkyl interaction | Arg118, Glu119, Val120, Glu114, Arg107, Leu149 | ||
| 6-1 | Van der Waals | Val48, Leu49, Ser50, Lys89, Ala90 | |
| Electrostatic | Val48, Leu49, Ser50, Gly52, Met53, Asp54 | ||
| Carbon hydrogen bond interaction | Val48, Leu49, Ser50 | ||
| Alkyl interaction | Val48, Leu49, Met53 | ||
| π-Donor hydrogen bond interaction | Met53 | ||
| 1E1D | Van der Waals | Asp480, Asp483, Val509, Val486, Met549 | |
| Electrostatic | Lys510, Asn482, Ser487, Arg399, Asn535, Lys537, Gly511, Arg513, Lys553, Tyr488 | ||
| Carbon hydrogen bond interaction | Asp480, Asn482, Lys510 | ||
| Conventional hydrogen bond interaction | Lys537, Arg513 | ||
| Halogen (fluorine) interaction | Asn535 | ||
| Alkyl interaction | Lys510 | ||
| π-Alkyl interaction | Tyr488 | ||
| 7-1 | Van der Waals | Phe61, Ala82 | |
| Electrostatic | Gln64, Ala68, Asn74, Arg79, Met83, Glu86, Val331 | ||
| Carbon hydrogen bond interaction | Glu86 | ||
| Conventional hydrogen bond interaction | Asn74, Arg79 | ||
| Alkyl interaction | Ala82, Ala83 | ||
| π-Alkyl interaction | Phe61, Arg79, Ala82, Met83 | ||
| Amide-π stacked interaction | Ala82, Met83 | ||
| π-Sulfur interaction | Met83 | ||
| 1USH | Van der Waals | Ala436, Met438, Lys441, Ala449, Pro507, Leu509, Lys512, Pro513, Gly514, Tyr515 | |
| Electrostatic | Asp437, Glu442, Asp445, Tyr446 | ||
| Carbon hydrogen bond interaction | Asp437, Glu442, Pro513 | ||
| Alkyl interaction | Ala436, Met438, Leu509, Lys512 | ||
| 11-5 | Van der Waals | Tyr221, His226, Gly227, Ser228, Ser253, Asn511, Pro513, Gly514, Arg516 | |
| Electrostatic | Asn229, His252, Gln254, Arg432, Asp510, Lys512, Lys515, Asn517, Gly519 | ||
| Carbon hydrogen bond interaction | Asn229, Asp510, Lys512, Lys515, Asn517 | ||
| Conventional hydrogen bond interaction | Arg432 | ||
| Halogen (fluorine) interaction | Asp510 | ||
| Alkyl interaction | Lys515 | ||
| π-Alkyl interaction | His252 | ||
| 2DYT | Van der Waals | Ile129, Asp131, Ile132, Leu135, Leu243, Leu291, Phe293, Ser296, Thr295, His303, Thr306 | |
| Electrostatic | Tyr209, Lys292 | ||
| Alkyl interaction | Ile132, Lys292, Leu243, | ||
| π-Alkyl interaction | Lys292 | ||
| π-Sigma interaction | Ser296 | ||
| 13-6 | Van der Waals | Glu87, Gly157, Leu159, Pro299, Ile301, Gln302, Asp304 | |
| Electrostatic | Asp77, Lys78, Val86, Gly88, Glu151, Glu152, Phe153, Ser300 | ||
| Carbon hydrogen bond interaction | Phe153, Pro299, Ser300, Ile301 | ||
| conventional hydrogen bond interaction | Glu152, His303 | ||
| π-interaction pair | His303 | ||
| π-Alkyl interaction | Phe153 | ||
| π-π Stacked interaction | His303 |
| Degrading Enzyme | Amino Acid Residue | Distance from the Target Fluoroquinolone Molecule | |||||
|---|---|---|---|---|---|---|---|
| Distance from CIP (Å) | Mean Distance (Å) | Distance from NOR (Å) | Mean Distance (Å) | Distance from OFL (Å) | Mean Distance (Å) | ||
| 1YZP | Ile91 | 7.50 | 8.10 | 7.80 | 8.57 | 8.10 | 7.03 |
| Phe93 | 9.10 | 9.70 | 7.40 | ||||
| Ile100 | 7.70 | 8.20 | 5.60 | ||||
| 3-6 | Arg91 | 18.50 | 19.80 | 23.40 | 24.17 | 22.70 | 24.20 |
| Arg93 | 20.20 | 24.70 | 25.00 | ||||
| Arg100 | 20.70 | 24.40 | 24.90 | ||||
| 4DTE | Phe181 | 6.20 | 5.50 | 9.10 | 6.77 | 5.30 | 4.57 |
| Val184 | 4.10 | 2.90 | 3.20 | ||||
| Pro349 | 6.20 | 8.30 | 5.20 | ||||
| 5-2 | Cys181 | 15.70 | 15.33 | 18.40 | 16.83 | 23.30 | 23.80 |
| Ser184 | 17.50 | 18.40 | 26.90 | ||||
| Lys349 | 12.80 | 13.70 | 21.20 | ||||
| 3NQA | Val155 | 8.50 | 5.60 | 5.40 | 5.87 | 7.00 | 4.97 |
| Ile200 | 3.70 | 3.90 | 2.90 | ||||
| Val201 | 4.60 | 8.30 | 5.00 | ||||
| 6-1 | Lys155 | 2.60 | 5.67 | 7.90 | 8.20 | 8.50 | 8.00 |
| Lys200 | 4.70 | 8.70 | 8.30 | ||||
| Lys201 | 9.70 | 8.00 | 7.20 | ||||
| 1E1D | Trp292 | 2.10 | 5.93 | 2.40 | 6.00 | 3.00 | 6.57 |
| Trp293 | 7.50 | 7.80 | 8.50 | ||||
| Tyr493 | 8.20 | 7.80 | 8.20 | ||||
| 7-1 | Lys292 | 16.70 | 20.27 | 15.30 | 17.97 | 19.20 | 23.00 |
| Lys293 | 23.30 | 20.10 | 25.70 | ||||
| Lys493 | 20.80 | 18.50 | 24.10 | ||||
| 1USH | Val432 | 6.90 | 7.17 | 8.30 | 6.90 | 4.20 | 6.77 |
| Tyr515 | 8.10 | 6.60 | 8.20 | ||||
| Val516 | 6.50 | 5.80 | 7.90 | ||||
| 11-5 | Arg432 | 11.00 | 7.60 | 6.20 | 6.40 | 10.10 | 7.63 |
| Lys515 | 5.50 | 6.30 | 6.90 | ||||
| Arg516 | 6.30 | 6.70 | 5.90 | ||||
| 2DYT | Ile136 | 8.20 | 8.50 | 7.80 | 8.13 | 7.80 | 7.90 |
| Met139 | 9.20 | 9.70 | 8.10 | ||||
| Leu258 | 8.10 | 6.90 | 7.80 | ||||
| 13-6 | Lys136 | 17.70 | 19.77 | 15.80 | 17.37 | 11.20 | 13.27 |
| Arg139 | 13.70 | 11.80 | 10.60 | ||||
| Arg258 | 27.90 | 24.50 | 18.00 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.-c.; Sun, S.-j.; Cui, P.; Ding, Y.-f. Molecular Modification of Fluoroquinolone-Biodegrading Enzymes Based on Molecular Docking and Homology Modelling. Int. J. Environ. Res. Public Health 2019, 16, 3407. https://doi.org/10.3390/ijerph16183407
Liu S-c, Sun S-j, Cui P, Ding Y-f. Molecular Modification of Fluoroquinolone-Biodegrading Enzymes Based on Molecular Docking and Homology Modelling. International Journal of Environmental Research and Public Health. 2019; 16(18):3407. https://doi.org/10.3390/ijerph16183407
Chicago/Turabian StyleLiu, Si-cheng, Shi-jun Sun, Peng Cui, and Yi-fan Ding. 2019. "Molecular Modification of Fluoroquinolone-Biodegrading Enzymes Based on Molecular Docking and Homology Modelling" International Journal of Environmental Research and Public Health 16, no. 18: 3407. https://doi.org/10.3390/ijerph16183407
APA StyleLiu, S.-c., Sun, S.-j., Cui, P., & Ding, Y.-f. (2019). Molecular Modification of Fluoroquinolone-Biodegrading Enzymes Based on Molecular Docking and Homology Modelling. International Journal of Environmental Research and Public Health, 16(18), 3407. https://doi.org/10.3390/ijerph16183407