Polymorphisms in PCSK9, LDLR, BCMO1, SLC12A3, and KCNJ1 Are Associated with Serum Lipid Profile in Chinese Han Population


Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Diagnostic Criteria
2.3. Epidemiological Investigation
2.4. Isolation of Genomic DNA
2.5. SNP Selection and Genotyping
2.6. Statistical Analysis
3. Results
3.1. General Characteristics, Serum Lipid Levels, and Lifestyles
3.2. SNPs Associated with TC, HDL-C, LDL-C, and TG
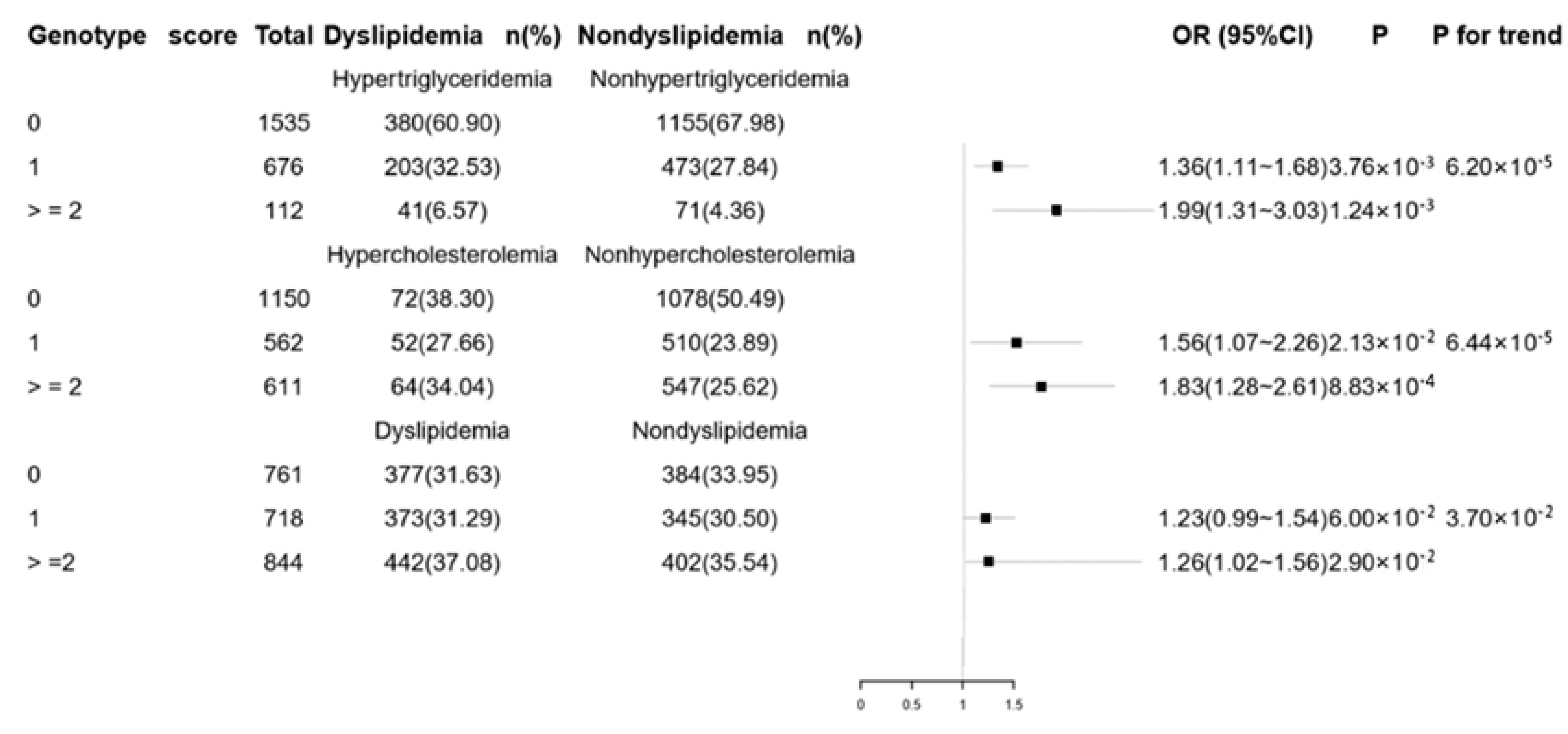
3.3. Combined Effects of Genetic Variants on Lipid Levels and Dyslipidemia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ghaddar, F.; Salameh, P.; Saleh, N.; Farhat, F.; Chahine, R.; Lahoud, N.; Hleyhel, M.; Zeidan, R.K. Noncardiac lebanese hospitalized adult patients’ awareness of their coronary artery disease risk factors. Vasc. Health Risk Manag. 2018, 14, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Helkin, A.; Stein, J.J.; Lin, S.; Siddiqui, S.; Maier, K.G.; Gahtan, V. Dyslipidemia Part 1—Review of Lipid Metabolism and Vascular Cell Physiology. Vasc. Endovasc. Surg. 2016, 50, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Isomaa, B.; Almgren, P.; Tuom, T.; Forsen, B.; Lahti, K.; Nissen, M.; Taskinen, M.R.; Groop, L. Cardiovascular morbidity and mortality associated with the metabolic syndrome. Diabetes Care. 2001, 24, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Stylianou, I.M.; Koseki, M.; Pirruccello, J.P.; Ripatti, S.; Pirruccello, J.P.; Chasman, D.I.; Willer, C.J.; Johansen, C.T. Biological, clinical and population relevance of 95 loci for blood lipids. NIH Public Access. 2010, 466, 707–713. [Google Scholar] [CrossRef]
- Yin, R.X.; Li, Y.Y.; Liu, W.Y.; Zhang, L.; Wu, J.Z. Interactions of the apolipoprotein A5 gene polymorphisms and alcohol consumption on serum lipid levels. PLoS ONE 2011, 6, e17954. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Namboodiri, K.K.; Green, P.P.; Kaplan, E.B.; Morrisom, J.A.; Chase, G.A.; Elston, R.C.; Owen, A.R.G.; Rifkind, B.M.; Tyroler, H.A.; Elston, R.C. The collaborative lipid research clinics program family study. IV. Familial associations of plasma lipids and lipoproteins. Am. J. Epidemiol. 1984, 119, 975–996. [Google Scholar] [CrossRef] [PubMed]
- Heller, D.A.; de Faire, U.; Pedersen, N.L.; Mcclearn, G.E. Genetic and environmental influences on serum lipid levels in twins. New England J. Med. 1993, 16, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Aulchenko, Y.S.; Ripatti, S.; Lindqvist, I.; Boomsma, D.; Iris, M. Loci influencing lipid levels and coronary heart disease risk in 16 European population cohorts. Nat. Genet. 2009, 41, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Kathiresan, S.; Melander, O.; Guiducci, C.; Surti, A.; Burtt, N.P.; Rieder, M.J.; Cooper, G.M.; Roos, C.; Voight, B.F.; Havulinna, A.S.; et al. Six new loci associated with blood low-density lipoprotein cholesterol, high-density lipoprotein cholesterol or triglycerides in humans. Nat. Genet. 2008, 40, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Kathiresan, S.; Willer, C.J.; Peloso, G.M.; Demissie, S.; Voight, B.F.; Bonnycastle, L.L.; Jackson, A.U.; Surti, A.; Guiducci, C.; Burtt, N.P.; et al. Common variants at 30 loci contribute to polygenic dyslipidemia. Nat. Genet. 2009, 41, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Sabatti, C.; Service, S.K.; Hartikainen, A.L.; Pouta, A.; Ripatti, S.; Brodsky, J.; Jones, C.G.; Zaitlen, N.A.; Varilo, T.; Kaakinen, M. Genome-wide association analysis of metabolic traits in a birth cohort from a founder population. Nat. Genet. 2009, 41, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, M.S.; Waterworth, D.M.; Debenham, S.L.; Wheeler, E.; Papadakis, K.; Zhao, J.H.; Songm, K.; Yuan, X.; Johnson, T.; Ashford, S.; et al. LDL-cholesterol concentrations: a genome-wide association study. Lancet. 2008, 371, 450–452. [Google Scholar] [CrossRef]
- Saxena, R.; Voight, B.F.; Lyssenko, V.; Burtt, N.P.; Bakker, P.I.W.; Chen, H.; Roix, J.J.; Kathiresan, S.; Hirschhorn, J.N.; Daly, M.J.; et al. Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science 2017, 316, 1331–1336. [Google Scholar] [CrossRef]
- Willer, C.J.; Sanna, S.; Jackson, A.U.; Scuteri, A.; Lori, L.; Clarke, R.; Heath, S.C.; Timpson, N.J.; Najjar, S.S.; Stringham, H.M.; et al. Newly identified loci that influence lipid concentrations and risk of coronary artery disease. Nat. Genet. 2008, 40, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Lye, S.H.; Chahil, J.K.; Bagali, P.; Alex, L.; Vadivelu, J.; Ahmad, W.A.W.; Chan, S.P.; Thong, M.K.; Zain, S.M.; Mohamed, R. Genetic polymorphisms in LDLR, APOB, PCSK9 and other lipid related genes associated with familial hypercholesterolemia in Malaysia. PLoS ONE 2013, 8, e60729. [Google Scholar] [CrossRef]
- Mangravite, L.M.; Medina, M.W.; Cui, J.; Pressman, S.; Smith, J.D.; Rieder, M.J.; Guo, X.; Nickerson, D.A.; Rotter, J.I.; Krauss, R.M. Combined influence of LDLR and HMGCR sequence variation on lipid-lowering response to simvastatin. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1485–1492. [Google Scholar] [CrossRef]
- Abifadel, M.; Rabes, J.P.; Jambart, S.; Halabyt, G.; Gannage-Yared, M.H.; Sarkis, A.; Beaino, G.; Varret, M.; Salem, N.; Corbani, S.; et al. The molecular basis of familial hypercholesterolemia in lebanon: Spectrum of LDLR mutations and role of PCSK9 as a modifier gene. Hum. Mutat. 2009, 7, E682–E691. [Google Scholar] [CrossRef]
- Guella, I.; Asselta, R.; Ardissino, D.; Merlini, P.A.; Peyvandi, F.; Kathiresan, S.; Mannucci, P.M.; Tubaro, M.; Duga, S. Effects of PCSK9 genetic variants on plasma LDL cholesterol levels and risk of premature myocardial infarction in the Italian population. J. Lipid Res. 2010, 51, 3342–3349. [Google Scholar] [CrossRef]
- Clifford, A.J.; Rincon, G.; Owens, J.E.; Medrano, J.F.; Moshfegh, A.J.; Baer, D.J.; Novotny, J.A. Single nucleotide polymorphisms in CETP, SLC46A1, SLC19A1, CD36, BCMO1, APOA5, and ABCA1 are significant predictors of plasma HDL in healthy adults. Lipids Health Dis. 2013, 1, 12–66. [Google Scholar] [CrossRef]
- Marchesini, G.; Forlani, G.; Cerrelli, F.; Manini, R.; Natale, S.; Baraldi, L.; Ermini, G.; Savorani, G.; Cerrelli, F.; Zocchi, D.; et al. Who and ATPIII proposals for the definition of the metabolic syndrome in patients with type 2 diabetes. Diabet Med. 2004, 21, 383–387. [Google Scholar] [CrossRef]
- Barany, F. Genetic disease detection and DNA amplification using cloned thermostable ligase. Proc. Natl. Acad. Sci. USA 1991, 88, 189–193. [Google Scholar] [CrossRef] [PubMed]
- McNamara, D.T.; Thomson, J.M.; Kasehagen, L.J.; Zimmerman, P.A. Development of a multiplex PCR-ligase detection reaction assay for diagnosis of infection by the four parasite species causing malaria in humans. J. Clin. Microbiol. 2004, 42, 2403–2410. [Google Scholar] [CrossRef]
- Tan, X.H.; Zhang, J.Y.; Di, C.H.; Hu, A.R.; Yang, L.; Qu, S.; Zhao, R.L.; Yang, P.R.; Guo, S.X. Distribution of CCR5-Δ32, CCR5m303A, CCR2-64I and SDF1-3′A in HIV-1 infected and uninfected high-risk Uighurs in Xinjiang, China. Infect. Genet. Evol. 2010, 10, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, D.Z.; Zhang, Z.; Song, Y.Q.; Zhang, D.; Zhao, T.; Chen, Z.; Sun, Y.; Zhang, D.; Yang, Y.; et al. Effects of genetic variants on lipid parameters and dyslipidemia in a Chinese population. J. Lipid Res. 2011, 52, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.A. When to use the Bonferroni correction. Ophthalmic Physiol. Opt. 2014, 34, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, D.M.; Ricketts, S.L.; Song, K.; Chen, L.; Zhao, J.H.; Ripatti, S.; Aulchenko, Y.S.; Zhang, W.; Yuan, X.; Lim, N.; et al. Genetic variants influencing circulating lipid levels and risk of coronary artery disease. Arterioscler Thromb. Vasc. Biol. 2010, 30, 2264–2276. [Google Scholar] [CrossRef] [PubMed]
- Ríos-González, B.E.; Ibarra-Cortés, B.; Ramírez-López, G.; Sánchez-Corona, J.; Magaña-Torres, M.T. Association of polymorphisms of genes involved in lipid metabolism with blood pressure and lipid values in Mexican hypertensive individuals. Dis. Markers 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Slimani, A.; Harira, Y.; Trabelsi, I.; Jomaa, W.; Maatouk, F.; Hamda, K.B.; Slimane, M.N. Effect of E670G polymorphism in PCSK9 gene on the risk and severity of coronary heart disease and ischemic stroke in a Tunisian cohort. J. Mol. Neurosci. 2014, 53, 150–157. [Google Scholar] [CrossRef]
- Webb, J.C.; Patel, D.D.; Shoulders, C.C.; Knight, B.L.; Soutar, A.K. Genetic variation at a splicing branch point in intron 9 of the low density lipoprotein (LDL)-receptor gene: A rare mutation that disrupts mRNA splicing in a patient with familial hypercholesterolaemia and a common polymorphism. Hum. Mol. Genet. 1996, 5, 1325–1331. [Google Scholar] [CrossRef]
- Andreotti, G.; Chen, J.B.; Gao, Y.T.; Rashid, A.; Chen, B.E.; Rosenberg, P.; Sakoda, L.C.; Deng, J.; Shen, M.C.; Wang, B.S.; et al. Polymorphisms of genes in the lipid metabolism pathway and risk of biliary tract cancers and stones: a population-based case-control study in Shanghai, China. Cancer Epidemiol. Prev. Biomark. 2008, 17, 525–534. [Google Scholar] [CrossRef]
- Tajamolian, M.; Kolahdouz, P.; Nikpour, P.; Forouzannia, S.K.; Sheikhha, M.H.; Yazd, E.F. A rare missense mutation and a polymorphism with high frequency in LDLR gene among iranian patients with familial hypercholesterolemia. Adv. Biomed. Res. 2018, 2, 7–37. [Google Scholar] [CrossRef]
- Ochoa, E.; Iriondo, M.; Manzano, C.; Fullaondo, A.; Villar, I.; Ruiz-Irastorza, G.; Zubiaga, A.M.; Estonba, A. LDLR and PCSK9 are associated with the presence of antiphospholipid antibodies and the development of thrombosis in aPLA carriers. PLoS ONE 2016, 11, e0146990. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Coordinators, N.R. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2016, 45, 12–17. [Google Scholar] [CrossRef]
- Bogdański, P.; Suliburska, J.; Szulińska, M.; Sikora, M.; Walkowiak, J.; Jakubowski, H. L-Arginine and vitamin C attenuate pro-atherogenic effects of high-fat diet on biomarkers of endothelial dysfunction in rats. Biomed. Pharmacother 2015, 76, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Habte-Tsion, H.M.; Ge, X.P.; Ren, M.; Xie, J.; Miao, L.; Lin, J.; Pan, W. Dietary arginine affects the insulin signaling pathway, glucose metabolism and lipogenesis in juvenile blunt snout bream Megalobrama amblycephala. Sci. Rep. 2017, 7, 7864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhuang, L.; Li, M.; Zhao, W.; Ge, X.; Chen, Y.; Wang, F.; Wang, N.; Bao, Y.; Liu, L.; et al. Arg913Gln of SLC12A3 gene promotes development and progression of end-stage renal disease in Chinese type 2 diabetes mellitus. Mol. Cell. Biochem. 2018, 437, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.; Perry, J.R.B.; Matteini, A.; Perola, M.; Tanaka, T.; Silander, K.; Rice, N.; Melzer, D.; Murray, A.; Cluett, C.; et al. Common variation in the β-carotene 15,15′-monooxygenase 1 gene affects circulating levels of carotenoids: a genome-wide association study. Am. J. Hum. Genet. 2009, 84, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yin, Q.; Ma, G.; Qian, Q. KCNQ1 gene polymorphisms are associated with lipid parameters in a Chinese Han population. Cardiovasc. Diabetol. 2010, 9, 35. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Charecteristics | Male | Female | Total |
|---|---|---|---|
| 1069 | 1254 | n = 2323 | |
| Age (years) | 60 (50~69) | 58 (49~65) | 59 (49–67) |
| BMI (kg/m2) | 22.85 (21.07~24.92) | 23.17 (21.17~25.23) | 23.00 (21.12~25.11) |
| Waist circumference (cm) | 82 (77~88) | 80 (75~86) | 81 (76–87) |
| TC (mmol/l) | 4.78 ± 0.90 | 4.98 ± 0.93 | 4.89 ± 0.92 |
| TG (mmol/l) | 1.20 (0.92~1.63) | 1.33 (0.99~1.86) | 1.28 (0.96~1.76) |
| HDL-C (mmol/l) | 1.30 ± 0.34 | 1.29 ± 0.28 | 1.29 ± 0.31 |
| LDL-C (mmol/l) | 2.98 ± 0.78 | 3.13 ± 0.84 | 3.06 ± 0.81 |
| SBP | 134.64 ± 19.20 | 133.92 ± 19.13 | 134.25 ± 19.16 |
| DBP | 82.66 ± 12.02 | 81.05 ± 11.86 | 81.79 ± 11.96 |
| Current smoking n (%) | 588 (55.00) | 23 (1.83) | 611 (26.30) |
| Current drinking n (%) | 592 (55.38) | 139 (11.08) | 731 (31.47) |
| Exercise n (%) | |||
| No | 729 (68.19) | 764 (60.92) | 1493 (64.27) |
| Regular | 340 (31.81) | 490 (39.07) | 830 (35.73) |
| SNP | Gene | Chromosome | Major Allele | Minor Allele | MAF a (%) | HWE-p Value |
|---|---|---|---|---|---|---|
| rs11646692 | BCMO1 | 16 | C | G | 34.22 | 0.037 |
| rs12934922 | BCMO1 | 16 | A | T | 13.53 | 0.875 |
| rs2479409 | PCSK9 | 1 | A | G | 31.62 | 0.217 |
| rs662145 | PCSK9 | 1 | C | T | 10.70 | 0.472 |
| rs2738466 | LDLR | 19 | A | G | 38.92 | 0.976 |
| rs1003723 | LDLR | 19 | C | T | 14.82 | 0.345 |
| rs6413504 | LDLR | 19 | A | G | 27.26 | 0.454 |
| rs11643718 | SLC12A3 | 16 | G | A | 8.79 | 0.251 |
| rs675759 | KCNJ1 | 11 | C | G | 12.52 | 0.921 |
| rs675388 | KCNJ1 | 11 | T | A | 11.70 | 0.804 |
| rs2846679 | KCNJ1 | 11 | G | A | 10.96 | 0.392 |
| Polymorphisms | TC mmol/L | HDL-C mmol/L | LDL-C mmol/L | TG mmol/L | |
|---|---|---|---|---|---|
| BCMO1 | AA (n = 1742) | 4.87 ± 0.90 | 1.30 ± 0.31 | 3.05 ± 0.80 | 1.26 (0.95~1.75) |
| rs12934922 | AT (n = 537) | 4.94 ± 0.99 | 1.29 ± 0.29 | 3.09 ± 0.84 | 1.31 (0.97~1.85) |
| TT (n = 44) | 4.88 ± 1.03 | 1.34 ± 0.29 | 3.08 ± 0.92 | 1.23 (0.94~1.69) | |
| effect a | 0.06 (−0.02~0.13) | −0.01 (−0.03~0.02) | 0.04 (−0.02~0.11) | 0.06 (0.01~0.12) | |
| p | 1.42 × 10−1 | 5.84 × 10−1 | 1.99 × 10−1 | 3.20 × 10−2 | |
| PCSK9 | AA (n = 1074) | 4.85 ± 0.89 | 1.29 ± 0.30 | 3.04 ± 0.79 | 1.27 (0.96~1.73) |
| rs2479409 | AG (n = 1026) | 4.93 ± 0.95 | 1.30 ± 0.33 | 3.09 ± 0.83 | 1.28 (0.95~1.77) |
| GG (n = 223) | 4.89 ± 0.95 | 1.27 ± 0.29 | 3.02 ± 0.84 | 1.29 (0.95~1.84) | |
| effect a | 0.04 (−0.02~0.10) | 0.00 (−0.02~0.02) | 0.01 (−0.04~0.06) | 0.01 (−0.03~0.06) | |
| p | 1.67 × 10−1 | 9.42 × 10−1 | 7.01 × 10−1 | 4.93×10−1 | |
| PCSK9 | CC (n = 1851) | 4.88 ± 0.92 | 1.30 ± 0.31 | 3.06 ± 0.83 | 1.26 (0.95~1.72) |
| rs662145 | CT (n = 441) | 4.94 ± 0.92 | 1.28 ± 0.33 | 3.07 ± 0.75 | 1.36 (1.00~1.92) |
| TT (n = 31) | 4.92 ± 0.87 | 1.24 ± 0.22 | 3.27 ± 0.90 | 1.23 (0.86~2.13) | |
| effect a | 0.06 (−0.03~0.14) | −0.02 (−0.05~0.01) | 0.03 (−0.04~0.11) | 0.09 (0.03~0.15) | |
| p | 1.89 × 10−1 | 1.91 × 10−1 | 3.57 × 10−1 | 7.64 × 10−4 | |
| LDLR | AA (n = 871) | 4.87 ± 0.90 | 1.30 ± 0.31 | 3.05 ± 0.80 | 1.26 (0.96~1.72) |
| rs2738466 | AG (n = 1102) | 4.92 ± 0.92 | 1.30 ± 0.32 | 3.08 ± 0.82 | 1.28 (0.95~1.75) |
| GG (n = 350) | 4.86 ± 0.96 | 1.25 ± 0.28 | 3.05 ± 0.82 | 1.30 (0.97~1.87) | |
| effect a | 0.01 (−0.06~0.05) | −0.02 (−0.04~0.00) | 0.00 (−0.05~0.05) | 0.01 (−0.03~0.05) | |
| p | 9.04 × 10−1 | 4.80 × 10−2 | 9.79 × 10−1 | 5.08×10−1 | |
| LDLR | CC (n = 1694) | 4.85 ± 0.90 | 1.30 ± 0.31 | 3.03 ± 0.79 | 1.28 (0.96~1.74) |
| rs1003723 | CT (n = 574) | 5.00 ± 0.98 | 1.31 ± 0.33 | 3.16 ± 0.87 | 1.28 (0.94~1.86) |
| TT (n = 55) | 4.99 ± 0.98 | 1.35 ± 0.26 | 3.18 ± 0.86 | 1.24 (0.91~1.79) | |
| effect a | 0.15 (0.08~0.22) | 0.02(−0.01~0.04) | 0.13 (0.06~0.19) | 0.02 (−0.03~0.08) | |
| p | 6.80 × 10−5 | 1.91 × 10−1 | 8.70 × 10−5 | 4.09×10−1 | |
| LDLR | AA (n = 1234) | 4.83 ± 0.90 | 1.29 ± 0.31 | 3.00 ± 0.77 | 1.28 (0.96~1.74) |
| rs6413504 | AG (n = 913) | 4.95 ± 0.95 | 1.29 ± 0.31 | 3.11 ± 0.86 | 1.28 (0.95~1.79) |
| GG (n = 176) | 4.59 ± 0.85 | 1.33 ± 0.32 | 3.17 ± 0.87 | 1.29 (0.92~1.85) | |
| effect a | 0.11 (0.05~0.17) | 0.01 (−0.01~0.03) | 0.10 (0.05~0.15) | 0.01 (−0.03~0.06) | |
| p | 1.45 × 10−4 | 5.14 × 10−1 | 1.55 × 10−4 | 5.45×10−1 | |
| SLC12A3 | GG (n = 1930) | 4.89 ± 0.91 | 1.30 ± 0.31 | 3.08 ± 0.81 | 1.26 (0.95~1.72) |
| rs11643718 | GA (n = 379) | 4.88 ± 0.93 | 1.28 ± 0.31 | 3.00 ± 0.83 | 1.35 (0.99~1.93) |
| AA (n = 14) | 4.90 ± 0.98 | 1.31 ± 0.25 | 2.59 ± 0.61 | 1.52 (0.76~2.34) | |
| effect a | −0.04(−0.07~0.08) | −0.02(−0.05~0.01) | −0.08(−0.16~0.00) | 0.11(0.05~0.18) | |
| p | 4.02 × 10−1 | 2.60 × 10−1 | 5.80 × 10−2 | 1.00 × 10−3 | |
| KCNJ1 | CC (n = 1774) | 4.89 ± 0.92 | 1.30 ± 0.31 | 3.06 ± 0.81 | 1.28 (0.96~1.74) |
| rs675759 | CG (n = 514) | 4.88 ± 0.93 | 1.30 ± 0.33 | 3.07 ± 0.82 | 1.30 (0.93~1.83) |
| GG (n = 35) | 4.90 ± 0.98 | 1.30 ± 0.26 | 2.99 ± 0.86 | 1.29 (1.09~1.83) | |
| effect a | 0.01 (−0.07~0.09) | 0.00 (−0.03~0.03) | 0.01 (−0.07~0.07) | 0.02 (−0.04~0.07) | |
| p | 8.83 × 10−1 | 8.73 × 10−1 | 9.31 × 10−1 | 5.85×10−1 | |
| KCNJ1 | TT (n = 1813) | 4.89 ± 0.92 | 1.29 ± 0.30 | 3.06 ± 0.81 | 1.28 (1.05~1.75) |
| rs675388 | TA (n = 478) | 4.91 ± 0.92 | 1.31 ± 0.33 | 3.08 ± 0.83 | 1.29 (0.92~1.83) |
| AA (n = 32) | 4.91 ± 1.04 | 1.28 ± 0.29 | 2.96 ± 0.90 | 1.29 (1.05~1.76) | |
| effect a | 0.04 (−0.04~0.12) | 0.01 (−0.02~0.04) | 0.01 (−0.06~0.08) | 0.01 (−0.05~0.07) | |
| p | 3.43 × 10−1 | 2.92 × 10−1 | 8.01 × 10−1 | 7.14×10−1 | |
| KCNJ1 | GG (n = 1843) | 4.90 ± 0.92 | 1.29 ± 0.31 | 3.07 ± 0.81 | 1.29 (0.96~1.76) |
| rs2846679 | GA (n = 457) | 4.84 ± 0.91 | 1.31 ± 0.33 | 3.06 ± 0.83 | 1.24 (0.92~1.76) |
| AA (n = 23) | 4.67 ± 0.95 | 1.25 ± 0.30 | 2.72 ± 0.73 | 1.28 (0.99~1.69) | |
| effect a | −0.05 (−0.14~0.03) | 0.01 (−0.02~0.04) | −0.03 (−0.10~0.05) | −0.02 (−0.08~0.04) | |
| p | 2.35 × 10−1 | 3.92 × 10−1 | 4.72 × 10−1 | 5.49×10−1 | |
| Lipids | Genotype Score a | Β (95% CI) | p for Trend | ||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |||
| TC | |||||||
| N | 1150 | 562 | 473 | 111 | 27 | ||
| Mean ± SD | 4.83 ± 0.89 | 4.81 ± 0.89 | 4.89 ± 0.91 | 4.99 ± 0.93 | 5.34 ± 0.85 | 0.085 (0.048~0.122) | 7.00 × 10−6 |
| LDL-C | |||||||
| N | 1150 | 562 | 473 | 111 | 27 | ||
| Mean ± SD | 3.01 ± 0.77 | 3.06 ± 0.83 | 3.15 ± 0.88 | 3.15 ± 0.88 | 3.41 ± 0.79 | 0.075 (0.042~0.108) | 9.00 × 10−6 |
| TG | |||||||
| N | 1535 | 676 | 102 | 10 | - | ||
| Median (IQR) | 1.23 (0.95~1.68) | 1.31 (0.96~1.86) | 1.37 (1.00~1.87) | 1.31 (0.98~2.08) | - | 0.096 (0.051~0.140) | 2.90×10−5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhao, T.; Tan, X.; Lei, S.; Huang, L.; Yang, L. Polymorphisms in PCSK9, LDLR, BCMO1, SLC12A3, and KCNJ1 Are Associated with Serum Lipid Profile in Chinese Han Population. Int. J. Environ. Res. Public Health 2019, 16, 3207. https://doi.org/10.3390/ijerph16173207
Li Z, Zhao T, Tan X, Lei S, Huang L, Yang L. Polymorphisms in PCSK9, LDLR, BCMO1, SLC12A3, and KCNJ1 Are Associated with Serum Lipid Profile in Chinese Han Population. International Journal of Environmental Research and Public Health. 2019; 16(17):3207. https://doi.org/10.3390/ijerph16173207
Chicago/Turabian StyleLi, Zheng, Tianyu Zhao, Xiaohua Tan, Song Lei, Liu Huang, and Lei Yang. 2019. "Polymorphisms in PCSK9, LDLR, BCMO1, SLC12A3, and KCNJ1 Are Associated with Serum Lipid Profile in Chinese Han Population" International Journal of Environmental Research and Public Health 16, no. 17: 3207. https://doi.org/10.3390/ijerph16173207
APA StyleLi, Z., Zhao, T., Tan, X., Lei, S., Huang, L., & Yang, L. (2019). Polymorphisms in PCSK9, LDLR, BCMO1, SLC12A3, and KCNJ1 Are Associated with Serum Lipid Profile in Chinese Han Population. International Journal of Environmental Research and Public Health, 16(17), 3207. https://doi.org/10.3390/ijerph16173207