Neonatal Bisphenol A Exposure Affects the IgM Humoral Immune Response to 4T1 Breast Carcinoma Cells in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Neonatal BPA Exposure
2.3. Mammary Tumour Induction
2.4. Cell Culture and Flow Cytometry
2.5. Two-Dimensional Immunoblot and Image Processing
2.6. Statistical Analysis
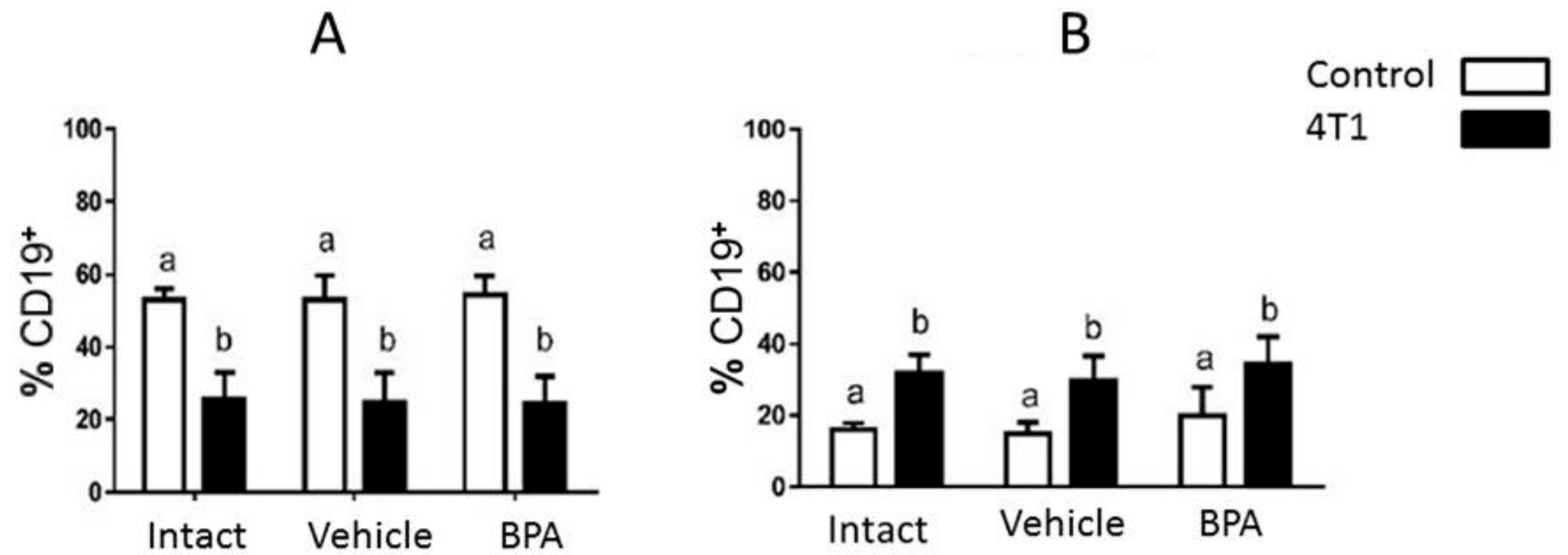
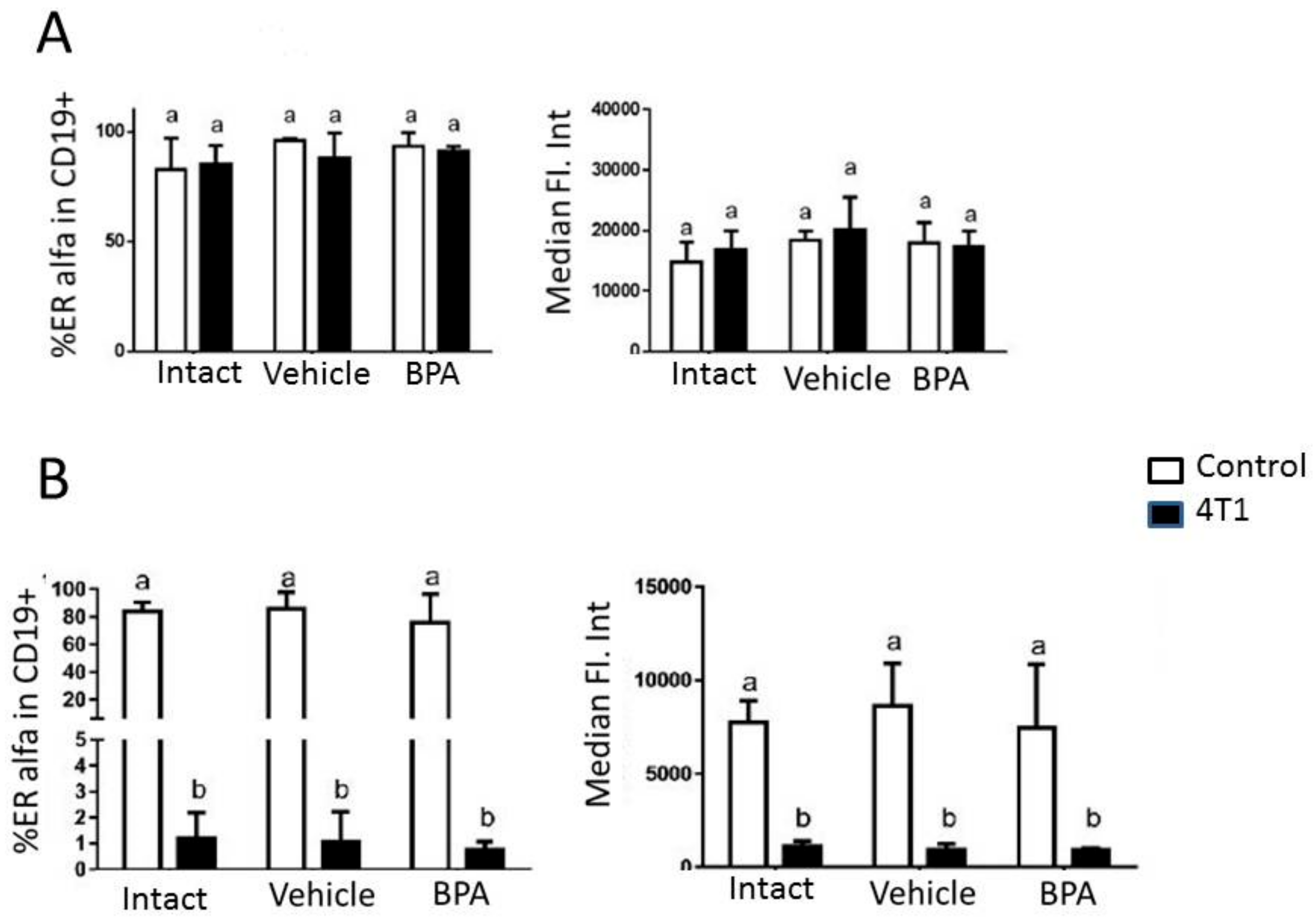
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stefanidou, M.; Maravelias, C.; Spiliopoulou, C. Human Exposure to Endocrine Disruptors and Breast Milk. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 269–276. [Google Scholar] [CrossRef]
- Welshons, W.V.; Nagel, S.C.; vom Saal, F.S. Large effects from small exposures. III. Endocrine mechanisms mediating effects of bisphenol A at levels of human exposure. Endocrinology 2006, 147, S56–S69. [Google Scholar] [CrossRef]
- Amaral Mendes, J.J. The endocrine disrupters: A major medical challenge. Food Chem. Toxicol. 2002, 40, 781–788. [Google Scholar] [CrossRef]
- Soto, A.M.; Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C. Does breast cancer start in the womb? Basic Clin. Pharmacol. Toxicol. 2008, 102, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Maffini, M.V.; Rubin, B.S.; Sonnenschein, C.; Soto, A.M. Endocrine disruptors and reproductive health: The case of bisphenol A. Mol. Cell Endocrinol. 2006, 254–255, 179–186. [Google Scholar] [CrossRef]
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. Mol. Biol. 2011, 127, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Zalko, D.; Jacques, C.; Duplan, H.; Bruel, S.; Perdu, E. Viable skin efficiently absorbs and metabolizes bisphenol A. Chemosphere 2011, 82, 424–430. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Palacios-Arreola, M.I.; Nava-Castro, K.E.; Río-Araiza, V.H.D.; Pérez-Sánchez, N.Y.; Morales-Montor, J. A single neonatal administration of Bisphenol A induces higher tumour weight associated to changes in tumor microenvironment in the adulthood. Sci. Rep. 2017, 7, 10573. [Google Scholar] [CrossRef] [PubMed]
- Chalubinski, M.; Kowalski, M.L. Endocrine disrupters—Potential modulators of the immune system and allergic response. Allergy 2006, 61, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Zaragoza, M.; Hernández, R.; Ostoa-Saloma, P. 2D immunoblots show differential response of mouse IgG and IgM antibodies to antigens of mammary carcinoma 4 T1 cells. Cancer Cell Int. 2014, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- NORMA Oficial Mexicana NOM-062-ZOO-1999. Available online: https://www.biomedicas.unam.mx/wp-content/pdf/unidad-de-modelos-biologicos/nom-062-zoo-1999.pdf?x21431 (accessed on 11 April 2019).
- Pulaski, B.A.; Ostrand-Rosenberg, S. Mouse 4T1 breast tumor model. In Current Protocols in Immunology; Coligan, J., John, E., Eds.; Wiley & Sons Inc.: New York, NY, USA, 2001. [Google Scholar]
- Garín, M.I.; Chu, C.-C.; Dela, G.; Cernuda-Morollón, E.; Wait, R.; Lechler, R. Galectin-1: A key effector of regulation mediated by CD4 CD25 T cells. Blood 2007, 109, 2058–2065. [Google Scholar] [CrossRef]
- Dalotto-Moreno, T.; Croci, D.O.; Cerliani, J.P.; Martinez-Allo, V.C.; Dergan-Dylon, S.; Mendez-Huergo, S.P.; Stupirski, J.C.; Mazal, D.; Osinaga, E.; Toscano, M.A.; et al. Targeting Galectin-1 Overcomes Breast Cancer-Associated Immunosuppression and Prevents Metastatic Disease. Cancer Res. 2013, 73, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, I.C. Germinal centers. Annu. Rev. Immunol. 1994, 12, 117–139. [Google Scholar] [CrossRef] [PubMed]
- McHeyzer-Williams, M.; Okitsu, S.; Wang, N.; McHeyzer-Williams, L. Molecular programming of B cell memory. Nat. Rev. Immunol. 2011, 12, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Weill, J.C.; Le Gallou, S.; Hao, Y.; Reynaud, C.A. Multiple players in mouse B cell memory. Curr. Opin. Immunol. 2013, 25, 334–338. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Coutinho, A.; Kazatchkine, M.D.; Avrameas, S. Natural autoantibodies. Curr. Opin. Immunol. 1995, 7, 812–818. [Google Scholar] [CrossRef]
- Baumgarth, N.; Herman, O.C.; Jager, G.C.; Herzenberg, L.A.; Herzenberg, L.A. Innate and acquired humoral immunities to influenza virus are mediated by distinct arms of the immune system. Proc. Natl. Acad. Sci. USA 1999, 96, 2250–2255. [Google Scholar] [CrossRef]
- MacLennan, C.; Fieschi, C.; Lammas, D.A.; Picard, C.; Dorman, S.E.; Sanal, O.; MacLennan, J.M.; Holland, S.M.; Ottenhoff, T.H.; Casanova, J.L.; et al. Interleukin IL-12 and IL-23 are key cytokines for immunity against Salmonella in humans. J. Infect. Dis. 2004, 190, 1755–1757. [Google Scholar] [CrossRef]
- Jouanguy, E.; Döffinger, R.; Dupuis, S.; Pallier, A.; Altare, F.; Casanova, J.L. IL-12 and IFN-gamma in host defense against mycobacteria and salmonella in mice and men. Curr. Opin. Immunol. 1999, 11, 346–351. [Google Scholar] [CrossRef]
- Gostner, J.M.; Raggl, E.; Becker, K.; Überalla, F.; Schennach, H.; Pease, J.E.; Fuchs, D. Bisphenol A suppresses Th1-type immune response in human peripheral blood mononuclear cells in vitro. Immunol. Lett. 2015, 168, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Sawai, C.; Anderson, K.; Walser-Kuntz, D. Effect of Bisphenol A on Murine Immune Function: Modulation of Interferon-gamma, IgG2a and Disease Symptoms in NZB × NZW F1 Mice. Environ. Health Perspect. 2003, 111, 1883–1887. [Google Scholar] [CrossRef]
- Waite, K.J.; Floyd, Z.E.; Arbour-Reily, P.; Stephens, J.M. Interferon-gamma-induced regulation of peroxisome proliferator-activated receptor gamma and STATs in adipocytes. J. Biol. Chem. 2001, 276, 7062–7068. [Google Scholar] [CrossRef] [PubMed]
- Riu, A.; Grimaldi, M.; le Maire, A.; Bey, G.; Phillips, K.; Boulahtouf, A.; Perdu, E.; Zalko, D.; Bourguet, W.; Balaguer, P. Peroxisome proliferator-activated receptor gamma is a target for halogenated analogs of bisphenol A. Environ. Health Perspect. 2011, 119, 1227–1232. [Google Scholar] [CrossRef]
- Ramon, S.; Bancos, S.; Thatcher, T.H.; Murant, T.I.; Moshkani, S.; Sahler, J.M.; Bottaro, A.; Sime, P.J.; Phipps, R.P. Peroxisome proliferator-activated receptor gamma B cell-specific deficient mice have an impaired antibody response. J. Immunol. 2012, 189, 4740–4747. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández Avila, R.; Palacios-Arreola, M.I.; Nava-Castro, K.E.; Morales-Montor, J.; Ostoa-Saloma, P. Neonatal Bisphenol A Exposure Affects the IgM Humoral Immune Response to 4T1 Breast Carcinoma Cells in Mice. Int. J. Environ. Res. Public Health 2019, 16, 1784. https://doi.org/10.3390/ijerph16101784
Hernández Avila R, Palacios-Arreola MI, Nava-Castro KE, Morales-Montor J, Ostoa-Saloma P. Neonatal Bisphenol A Exposure Affects the IgM Humoral Immune Response to 4T1 Breast Carcinoma Cells in Mice. International Journal of Environmental Research and Public Health. 2019; 16(10):1784. https://doi.org/10.3390/ijerph16101784
Chicago/Turabian StyleHernández Avila, Ricardo, Margarita I. Palacios-Arreola, Karen E. Nava-Castro, Jorge Morales-Montor, and Pedro Ostoa-Saloma. 2019. "Neonatal Bisphenol A Exposure Affects the IgM Humoral Immune Response to 4T1 Breast Carcinoma Cells in Mice" International Journal of Environmental Research and Public Health 16, no. 10: 1784. https://doi.org/10.3390/ijerph16101784
APA StyleHernández Avila, R., Palacios-Arreola, M. I., Nava-Castro, K. E., Morales-Montor, J., & Ostoa-Saloma, P. (2019). Neonatal Bisphenol A Exposure Affects the IgM Humoral Immune Response to 4T1 Breast Carcinoma Cells in Mice. International Journal of Environmental Research and Public Health, 16(10), 1784. https://doi.org/10.3390/ijerph16101784