Finasteride-Induced Inhibition of 5α-Reductase Type 2 Could Lead to Kidney Damage—Animal, Experimental Study

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Hormone Assays
2.3. Immunohistochemistry (IHC)
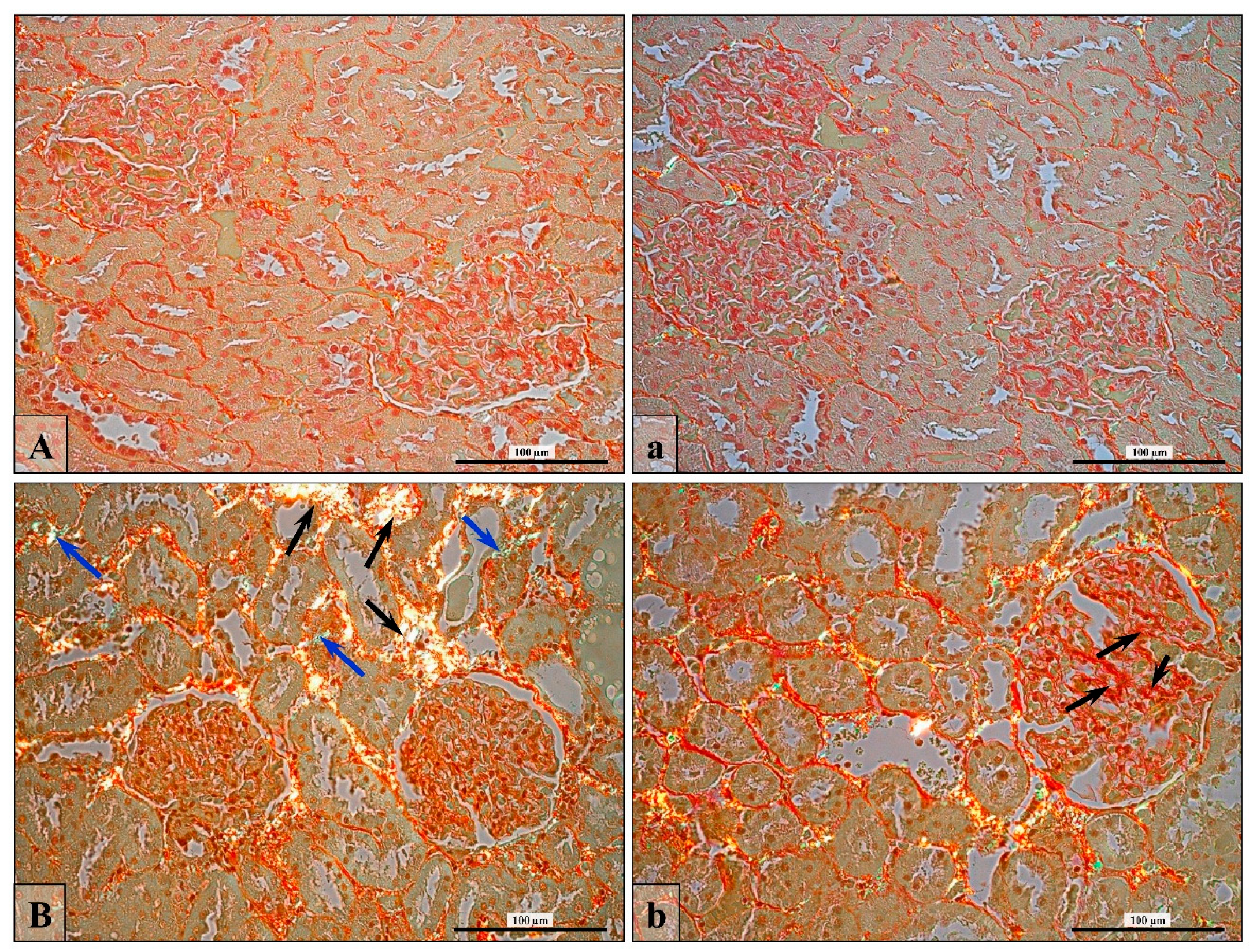
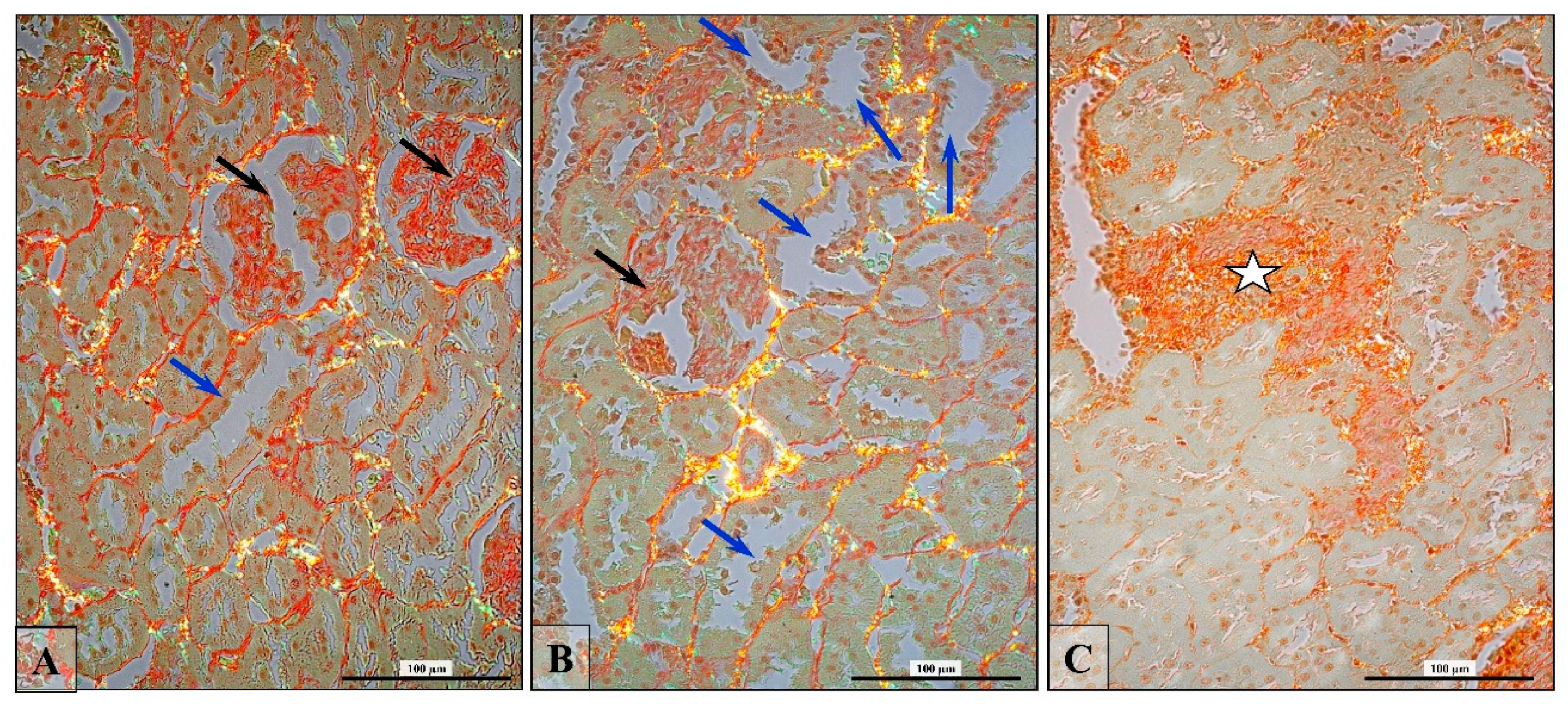
2.4. Collagen Fiber Visualization and Validation
2.5. Apoptosis In Situ Detection
Validation of TUNEL-Positive (Apoptotic Cell) and PCNA-Positive (Proliferating Cell)
2.6. Statistical Analysis
3. Results
3.1. Sex Hormone Levels
3.2. Androgen Receptor Expression
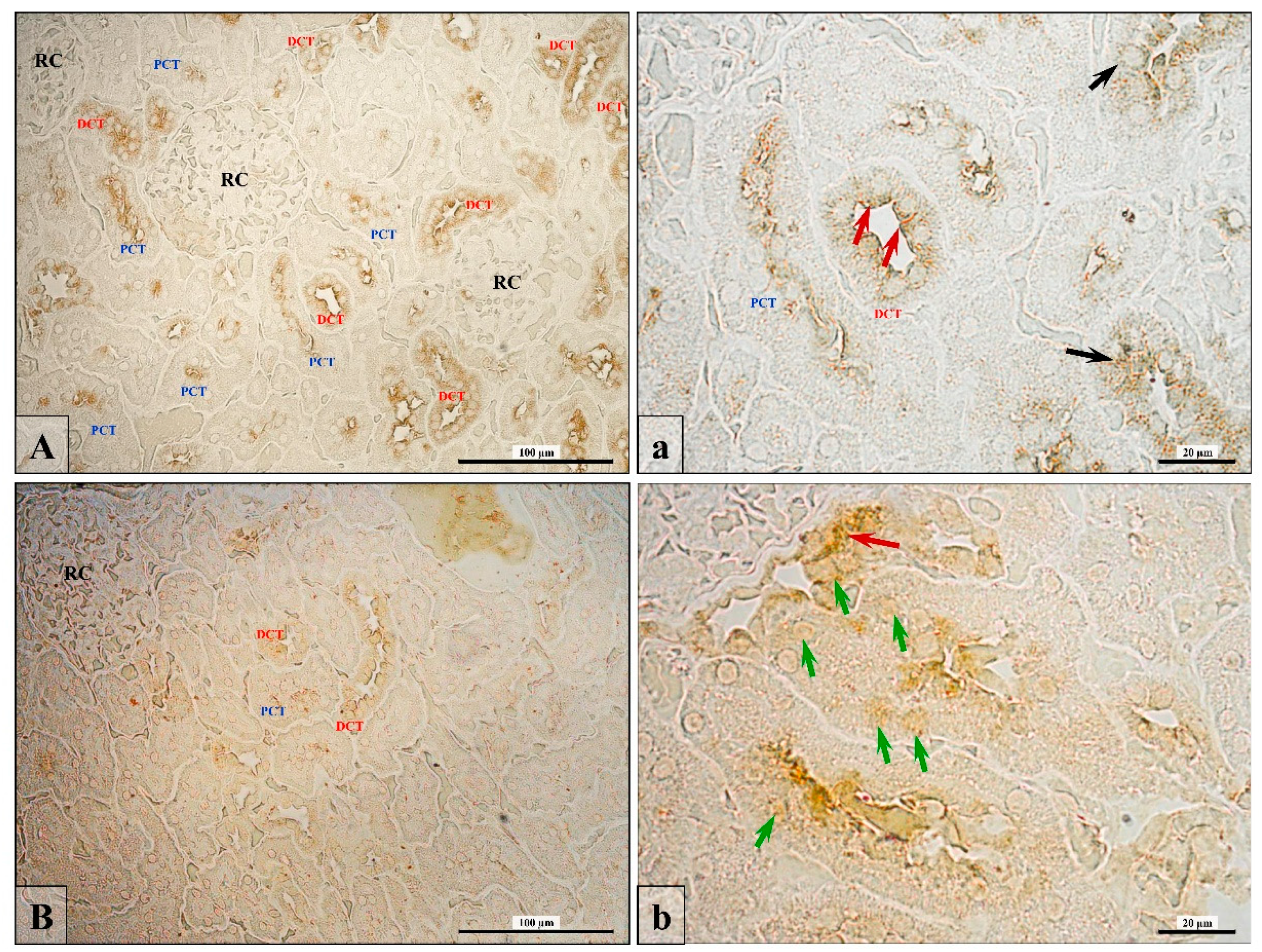
3.3. Junctional Protein Expression
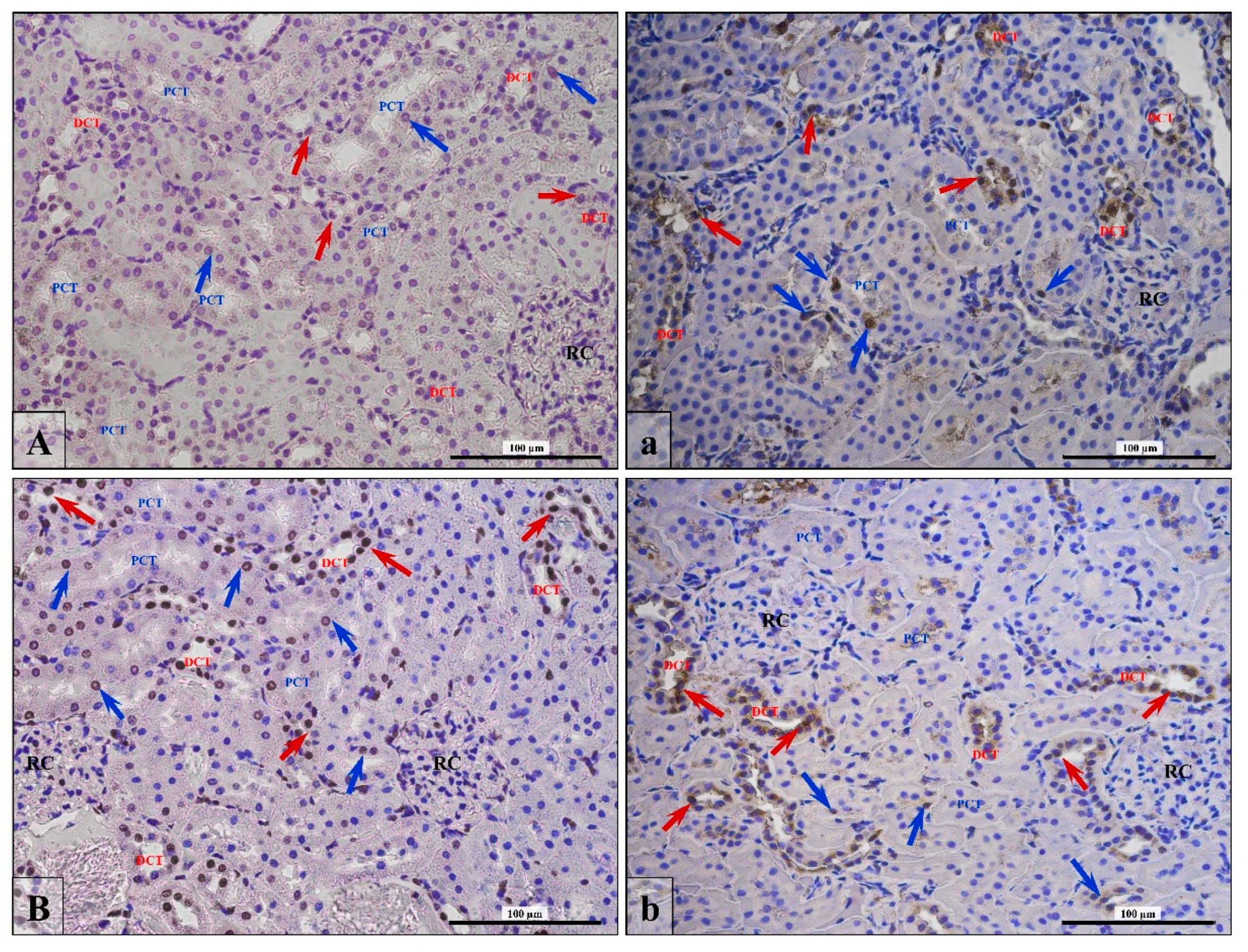
3.4. Apoptosis/Proliferation Ratio
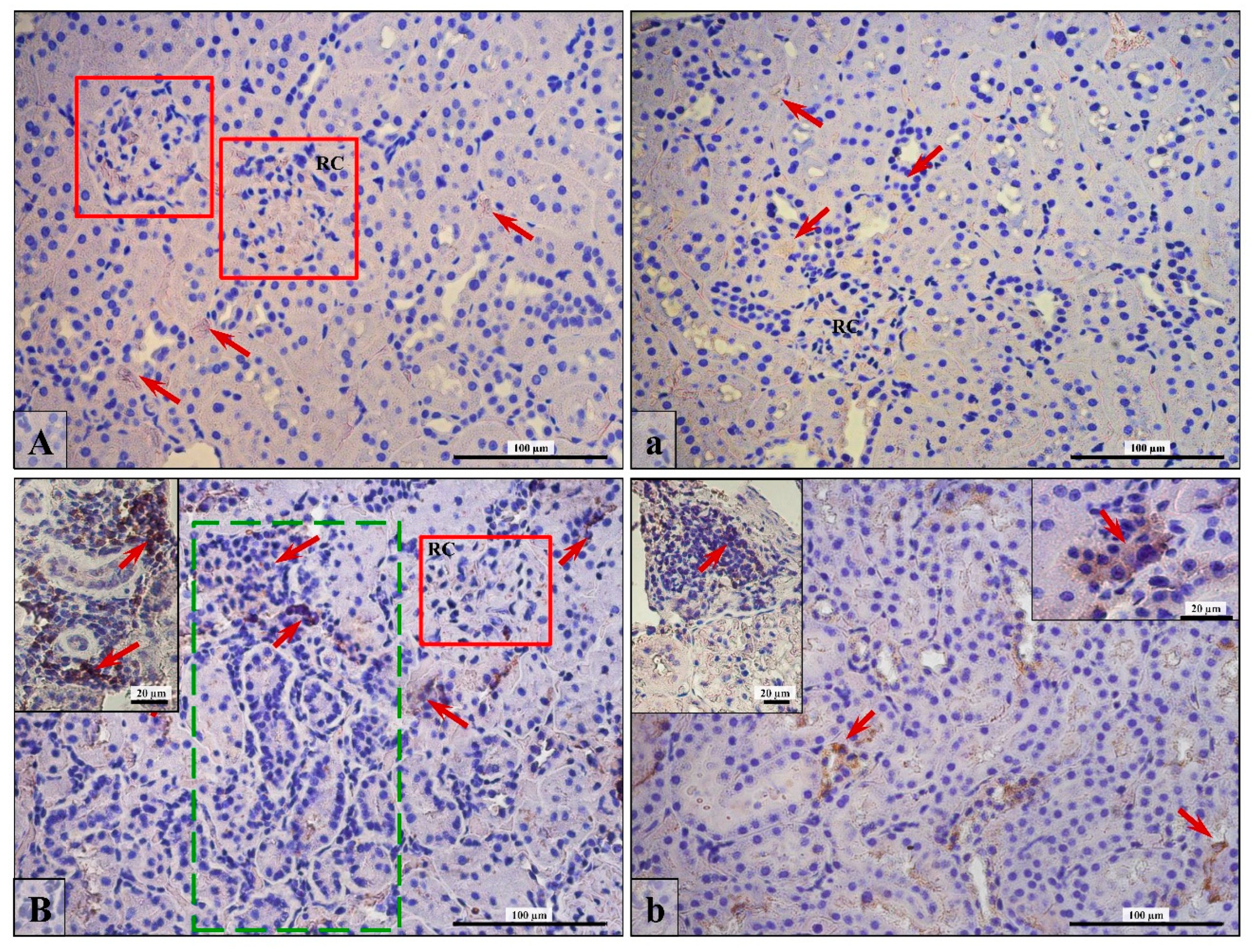
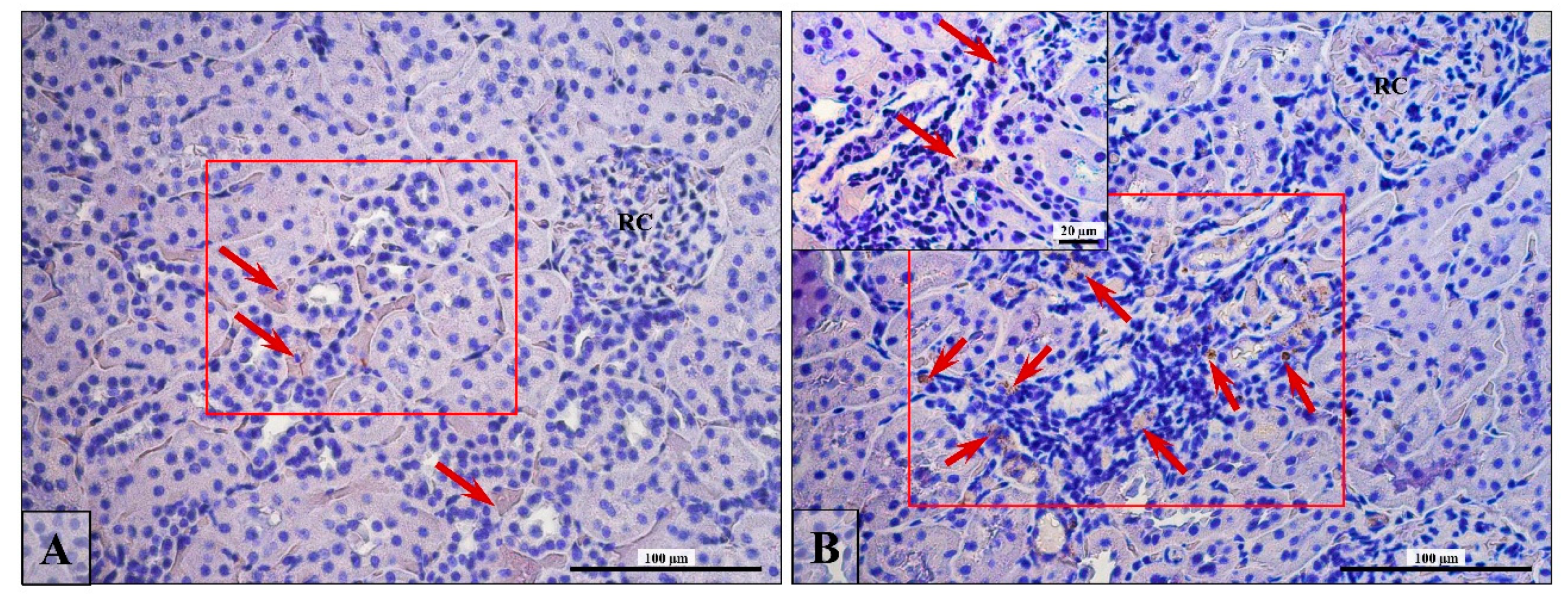
3.5. Lymphocytes T and B Specific Markers and IL-6 Expression
3.6. Renal Fibrosis (Collagen Fiber Thickening)
4. Discussion
5. Conclusions
- The finasteride treatment of adult male rats led to a decrease in androgen receptor expression and its cellular translocation within the kidney cortex.
- The pathomorphological changes (glomerulosclerosis, tubulosclerosis, dysplastic glomeruli, and tubules with lumen dilatation) in rats’ kidneys with disturbed steroid hormone imbalance were associated with the diminished expression of intracellular junctional proteins.
- The changed apoptotic/proliferating ratio of nephron cells and the increase in the numberof lymphocytes in the area of pathologically altered convoluted tubules were accompaniedby impaired androgen/estrogen homeostasis.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and ist disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef]
- Metcalf, B.W.; Levy, M.A.; Holt, D.A. Inhibitor of steroid 5-reductase in benign prostatic hyperplasia, male pattern baldness and acne. Trends Pharmacol. Sci. 1989, 10, 491–495. [Google Scholar] [CrossRef]
- Peters, D.H.; Sorkin, E.M. Finasteride: A review of its potential in the treatment of benign prostatic hyperplasia. Drugs 1993, 46, 77–208. [Google Scholar] [CrossRef] [PubMed]
- McClellan, K.J.; Markham, A. Finasteride: A review of its use in male pattern hair loss. Drugs 1999, 51, 111–126. [Google Scholar] [CrossRef] [PubMed]
- The Finasteride Male Pattern Hair Loss Group. Long-term (5-year) multinational experience with finasteride 1 mg in the treatment of men with androgenic alopecia. Eur. J. Dermatol. 2002, 12, 38–49. [Google Scholar]
- Lapi, F.; Azoulay, L.; Niazi, M.T.; Yin, H.; Benayoun, S.; Suissa, S. Androgen deprivation therapy and risk of acute kidney injury in patients with prostate cancer. JAMA 2013, 310, 289–296. [Google Scholar] [CrossRef]
- Molinari, C.; Battaglia, A.; Grossini, E.; Mary, D.A.; Vassanelli, C.; Vacca, G. The effect of testosterone on regional blood flow in prepubertal anaesthetized pigs. J. Physiol. 2002, 543, 365–372. [Google Scholar] [CrossRef]
- Hutchens, M.P.; Fujiyoshi, T.; Komers, R.; Herson, P.S.; Anderson, S. Estrogen protects renal endothelial barrier function from ischemia-reperfusion in vitro and in vivo. Am. J. Physiol. Ren. Physiol. 2012, 303, F377–F385. [Google Scholar] [CrossRef]
- Sadler, T.W. Urogenital System. In Langman’s Medical Embryology; Sadler, T.W., Ed.; Lippincott Williams & Wilkins/Wolters Kluwer Company: Philadelphia, PA, USA, 2004; pp. 321–362. [Google Scholar]
- Reyes, J.L.; Meléndez, E.; Alegría, A.; Jaramillo-Juárez, F. Influence of sex differences on the renal secretion of organic anions. Endocrinology 1998, 139, 1581–1587. [Google Scholar] [CrossRef]
- Schmidt, A.; Luger, A.; Hörl, W.H. Sexual hormone abnormalities in male patients with renal failure. Nephrol. Dial. Transpl. 2002, 17, 368–371. [Google Scholar] [CrossRef]
- Sharma, K.; Ziyadeh, F.N. The emerging role of the transforming growth factor-β in the kidney disease. Am. J. Physiol. 1994, 266, F829–F842. [Google Scholar] [CrossRef]
- Liang, L.; Li, L.; Tian, J.; Lee, S.O.; Dang, Q.; Huang, C.K.; Yeh, S.; Erturk, E.; Bushinsky, D.; Chang, L.S.; He, D.; Chang, C. Androgen receptor enhances kidney stone-CaOx crystal formation via modulation of oxalate biosynthesis & oxidative stress. Mol. Endocrinol. 2014, 28, 1291–1303. [Google Scholar]
- Khalil, R.; Kim, N.R.; Jardi, F.; Vanderschueren, D.; Claessens, F.; Decallonne, B. Sex steroids and the kidney: Role in renal calcium and phosphate handling. Mol. Cell. Endocrionl. 2018, 465, 61–72. [Google Scholar] [CrossRef]
- Antus, B.; Yao, Y.; Liu, S.; Song, E.; Lutz, J.; Heemann, U. Contribution of androgens to chronic allograft nephropathy is mediated by dihydrotestosterone. Kidney Int. 2001, 60, 1955–1963. [Google Scholar] [CrossRef] [PubMed]
- Antus, B.; Yao, Y.; Song, E.; Liu, S.; Lutz, J.; Heemann, U. Opposite effects of testosterone and estrogens on chronic allograft nephropathy. Transpl. Int. 2002, 15, 494–501. [Google Scholar] [CrossRef]
- Doublier, S.; Lupia, E.; Catanuto, P.; Periera-Simon, S.; Xia, X.; Korach, K.; Berho, M.; Elliot, S.J.; Karl, M. Testosterone and 17β-estradiol have opposite effects on podocyte apoptosis that precedes glomerulosclerosis in female estrogen receptor knockout mice. Kidney Int. 2011, 79, 404–413. [Google Scholar] [CrossRef]
- Sakemi, T.; Baba, N. Castration attenuates proteinuria and glomerular injury in unilaterally nephrectomized male Sprague-Dawley rats. Lab. Investig. 1993, 69, 51–57. [Google Scholar]
- Sakemi, T.; Toyoshima, H.; Morito, F. Testosterone eliminates attenuating effect of castration on the progressive glomerular injury in hypercholesterolemic male Imai rats. Nephron 1994, 67, 469–476. [Google Scholar] [CrossRef]
- Baylis, C. Age-dependent glomerular damage in rat: Dissociation between glomerular injury and both glomerular hypertension and hypertrophy: Male gender as a primary risk factor. J. Clin. Investig. 1994, 94, 1823–1829. [Google Scholar] [CrossRef]
- Reckelhoff, J.F.; Granger, J.P. Role of androgens in mediating hypertension and renal injury. Clin. Exp. Pharmacol. Physiol. 1999, 26, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Neugarten, J.; Acharya, A.; Silbiger, S.R. Effect of gender on the progression of nondiabetic renal disease: A meta-analysis. J. Am. Soc. Nephrol. 2000, 11, 319–329. [Google Scholar]
- Iliescu, R.; Reckelhoff, J.F. Sex and the kidney. Hypertension 2008, 51, 1000–1001. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, K. Mechanisms underlying sex differences in progressive renal disease. Gend. Med. 2008, 5, 10–23. [Google Scholar] [CrossRef]
- Goldberg, I.; Krause, I. The role of gender in chronic kidney diseases. EMJ 2016, 1, 58–64. [Google Scholar]
- Sullivan, J.C.; Gillis, E.E. Sex and gender differences in hypertensive kidney injury. Am. J. Physiol. Ren. Physiol. 2017, 313, F1009–F1017. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Roeher, O.; Hickstein, H.; Korht, S. Blood pressure guided profiling of ultrafiltration during hemodialysis. Saudi J. Kidney Dis. Transpl. 2001, 12, 337–344. [Google Scholar]
- Neugarten, J.; Golestaneh, L. Gender and the prevalence and progression of renal disease. Adv. Chronic Kidney Dis. 2013, 20, 390–395. [Google Scholar] [CrossRef]
- Silbiger, S.R.; Neugarten, J. The impact of gender on the progression of chronic renal disease. Am. J. Kidney Dis. 1995, 25, 515–533. [Google Scholar] [CrossRef]
- Schneeberger, E.E.; Lynch, R.D. The tight junction: A multifunctional complex. Am. J. Physiol. Cell Physiol 2004, 286, C1213–C1228. [Google Scholar] [CrossRef] [PubMed]
- Capaldo, C.T.; Macara, I.G. Depletion of E-cadherin disrupts establishment but not maintenance of cell junctions in Madin-Darby canine kidney epithelial cells. Mol. Biol. Cell 2007, 18, 189–200. [Google Scholar] [CrossRef]
- Theard, D.; Steiner, M.; Kalicharan, D.; Hoekstra, D.; van Ijzendoorn, S.C. Cell polarity development and protein trafficking in hepatocytes lacking E-cadherin/beta-catenin-based adherens junctions. Mol. Biol. Cell 2007, 18, 2313–2321. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Firestone, G.L.; Kapadia, B.J. Minireview: Regulation of gap junction dynamics by nuclear hormone receptors and their ligands. Mol. Endocrinol. 2012, 26, 1798–1807. [Google Scholar] [CrossRef] [PubMed]
- Gary, L.; Firestone, G.L.; Kapadia, B.J. Minireview: Steroid/nuclear receptor-regulated dynamics of occluding and anchoring junctions. Mol. Endocrinol. 2014, 28, 1769–1784. [Google Scholar]
- Gonzalez-Mariscal, L.; Namorado, M.C.; Martin, D.; Luna, J.; Alarcon, L.; Islas, S.; Valencia, L.; Muriel, P.; Ponce, L.; Reyes, J.L. Tight junction proteins ZO-1, ZO-2, and occludin along isolated renal tubules. Kidney Int. 2000, 57, 2386–2402. [Google Scholar] [CrossRef]
- Balkovetz, D.F. Claudins at the gate: Determinants of renal epithelial tight junction paracellular permeability. Am. J. Physiol. Ren. Physiol. 2006, 290, F572–F579. [Google Scholar] [CrossRef]
- Balkovetz, D.F. Tight junction claudins and the kidney in sickness and in health. Biochim. Biophys. Acta 2009, 1788, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Prozialeck, W.C.; Lamar, P.C.; Appelt, D.M. Differential expression of E-cadherin, N-cadherin and beta-catenin in proximal and distal segments of the rat nephron. BMC Physiol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Tian, X.; Liu, Z.; Niu, B.; Zhang, J.; Tan, T.K.; Lee, S.R.; Zhao, Y.; Harris, D.C.; Zheng, G. E-cadherin/β-catenin complex and the epithelial barrier. J. Biomed. Biotechnol. 2011, 2011, 567305. [Google Scholar] [CrossRef] [PubMed]
- Hanner, F.; Sorensen, C.M.; Holstein-Rathlou, N.H.; Peti-Peterdi, J. Connexins and the kidney. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1143–R1155. [Google Scholar] [CrossRef]
- Mese, G.; Richard, G.; White, T.W. Gap Junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2516–2524. [Google Scholar] [CrossRef] [PubMed]
- Huynh, H.T.; Alpert, L.; Laird, D.W.; Batias, G.; Chalifour, L.; Alaoui-Jamali, M.A. Regulation of the gap junction connexin 43 gene by androgens in the prostate. J. Mol. Endocrinol. 2001, 26, 1–10. [Google Scholar] [CrossRef]
- Kolasa, A.; Marchlewicz, M.; Wenda-Różewicka, L.; Wiszniewska, B. DHT deficiency perturbs the integrity of the rat seminiferous epithelium by disrupting tight and adherens junctions. Folia Histochem. Cytobiol. 2011, 49, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Kolasa-Wołosiuk, A.; Misiakiewicz-Has, K.; Baranowska-Bosiacka, I.; Gutowska, I.; Wiszniewska, B. Androgen levels and apoptosis in the testis during postnatal development of finasteride-treated male rat offspring. Folia Histiochem. Cytobiol. 2015, 53, 236–248. [Google Scholar] [CrossRef]
- Kolasa-Wołosiuk, A.; Misiakiewicz-Has, K.; Baranowska-Bosiacka, I.; Gutowska, I.; Tarnowski, M.; Tkacz, M.; Wiszniewska, B. Connexin 43 expression in the testes during postnatal development of finasteride-treated male rat offspring. Arch. Med. Sci. 2018, 14, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Ashby, J.; Tinwell, H.; Odum, J.; Lefevre, P. Natural variability and the influence of concurrent control values on the detection and interpretation of low-dose or weak endocrine toxicities. Environ. Health Perspect. 2004, 112, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Kearbey, J.D.; Nair, V.A.; Chung, K.; Parlow, A.F.; Miller, D.D.; Dalton, J.T. Comparison of the pharmacological effects of a novel selective androgen receptor modulator, the 5alpha-reductase inhibitor finasteride, and the anti-androgen hydroxyl flutamide in intact rats: New approach for benign prostate hyperplasia. Endocrinology 2004, 145, 5420–5428. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Kędzierska, K.; Sporniak-Tutak, K.; Kolasa, A.; Domański, L.; Domański, M.; Sindrewicz, K.; Smektała, T.; Bober, J.; Safranow, K.; Osekowska, B.; et al. The effect of immunosuppressive therapy on renal cell apoptosis in native rat kidneys. Histol. Histopathol. 2015, 30, 105–116. [Google Scholar] [CrossRef]
- PROPECIA—Finasteride Tablet, Film Coated. Available online: http://archive.is/xdDiK#selection-131.0-131.621 (accessed on 24 March 2019).
- Amory, J.K.; Watts, N.B.; Easley, K.A.; Sutton, P.R.; Anawalt, B.D.; Matsumoto, A.M.; Bremner, W.J.; Tenover, J.L. Exogenous testosterone or testosterone with finasteride increases bone mineral density in older men with low serum testosterone. J. Clin. Endocrinol. Metab. 2004, 89, 503–510. [Google Scholar] [CrossRef]
- Vaughan, C.; Goldstein, F.C.; Tenover, J.L. Exogenous testosterone alone or with finasteride does not improve measurements of cognition in healthy older men with low serum testosterone. J. Androl. 2007, 28, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Takeda, H.; Chodak, G.; Mutchnik, S.; Nakamoto, T.; Chang, C. Immunohistochemical localization of androgen receptors with mono- and polyclonal antibodies to androgen receptor. J. Endocrinol. 1999, 126, 17–25. [Google Scholar] [CrossRef]
- Kimura, N.; Mizokami, A.; Oonuma, T.; Sasano, H.; Nagura, H. Immunocytochemical localization of androgen receptor with polyclonal antibody in paraffin-embedded human tissues. J. Histochem. Cytochem. 1993, 41, 671–678. [Google Scholar] [CrossRef]
- Langner, C.; Ratschek, M.; Rehak, P.; Schips, L.; Zigeuner, R. Steroid hormone receptor expression in renal cell carcinoma: An immunohistochemical analysis of 182 tumors. J. Urol. 2004, 171, 611–614. [Google Scholar] [CrossRef]
- Li, J.Y.; Zhou, T.; Gao, X.; Xu, C.; Sun, Y.; Peng, Y.; Chang, Z.; Zhang, Y.; Jiang, J.; Wang, L.; et al. Testosterone and androgen receptor in human nephrolithiasis. J. Urol. 2010, 184, 2360–2363. [Google Scholar] [CrossRef] [PubMed]
- Yakirevich, E.; Matoso, A.; Morris, D.J.; Resnick, M.B. Steroid Receptors in Renal Cell Carcinoma. In Emerging Research and Treatments in Renal Cell Carcinoma; Robert, A., Ed.; In Tech: Ramsey, MN, USA, 2012; pp. 99–126. [Google Scholar]
- Turner, K.J.; Morley, M.; MacPherson, S.; Millar, M.R.; Wilson, J.A.; Sharpe, R.M.; Saunders, P.T. Modulation of gene expression by androgen and oestrogens in the testis and prostate of the adult rat following androgen withdrawal. Mol. Cell. Endocrinol. 2001, 178, 73–87. [Google Scholar] [CrossRef]
- Liu, T.; Wu, Z.; Fulton, M.D.; Johnson, J.M.; Berkman, E.C. Prolonged androgen deprivation leads to downregulation of androgen receptor and prostate-specific membrane antigen in prostate cancer cells. Int. J. Oncol. 2012, 41, 2087–2092. [Google Scholar] [CrossRef] [PubMed]
- Deslypere, J.-P.; Young, M.; Wilson, J.D.; McPhaul, M.J. Testosterone and 5α-dihydrotestosterone interact differently with the androgen receptor to enhance transcription of the MMTV-CAT reporter gene. Mol. Cell. Endocrinol. 1992, 88, 15–22. [Google Scholar] [CrossRef]
- Chang, C.; Lee, S.O.; Yeh, S.; Chang, T.M. Androgen receptor (AR) differential roles in hormone-related tumors including prostate, bladder, kidney, lung, breast and liver. Oncogene 2014, 33, 3225–3234. [Google Scholar] [CrossRef] [PubMed]
- Bass, R.; Perry, B.; Langenstroer, P.; Thrasher, J.B.; Dennis, K.L.; Tawfik, O.; Holzbeierlein, J. Effects of short-term finasteride on apoptotic factors and androgen receptors in prostate cancer cells. J. Urol. 2009, 181, 615–619. [Google Scholar] [CrossRef]
- Iguchi, M.; Takamura, C.; Umekawa, T.; Kurita, T.; Kohri, K. Inhibitory effects of female sex hormones on urinary stone formation in rats. Kidney Int. 1999, 56, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Panet-Raymond, V.; Gottlieb, B.; Umekawa, T.; Kurita, T.; Kohri, K. Interactions between androgen and estrogen receptors and the effects on their transactional properties. Mol. Cell. Endocrinol. 2000, 167, 139–150. [Google Scholar] [CrossRef]
- Lee, S.H. Coexistence of cytoplasmic and nuclear estrogen receptors. A histochemical study on human mammary cancer and rabbit uterus. Cancer 1989, 64, 1461–1466. [Google Scholar] [CrossRef]
- Benten, W.P.M.; Stephan, C.; Lieberherr, M.; Wunderlich, F. Estradiol signaling via sequestrable surface receptors. Endocrinology 2001, 142, 1669–1677. [Google Scholar] [CrossRef]
- Gruber, C.J.; Tschugguel, W.; Schneeberger, C.; Huber, J.C. Production and actions of estrogens. N. Engl. J. Med. 2002, 346, 340–352. [Google Scholar] [CrossRef]
- Migliaccio, A.; Castoria, D.; Di Domenico, M.; de Falco, A.; Bilancio, A.; Lombardi, M.; Bottero, D.; Varricchio, L.; Nanayakkara, M.; Rotondi, A.; et al. Sex steroid hormones act as growth factor. J. Steroid Biochem. Mol. Biol. 2002, 83, 31–35. [Google Scholar] [CrossRef]
- Shihan, M.; Bulldan, A.; Scheiner-Bobis, G. Non-classical testosterone signaling is mediated by a G-protein-coupled receptor interacting with Gnα11. Biochim. Biophys. Acta 2014, 1843, 1172–1181. [Google Scholar] [CrossRef]
- Thomas, P.; Converse, A.; Berg, H.A. ZIP9, a novel membrane androgen receptor and zinc transporter protein. Gen. Comp. Endocriol. 2018, 257, 130–136. [Google Scholar] [CrossRef]
- Zhu, G.; Liang, L.; Li, L.; Dang, Q.; Song, W.; Yeh, S.; He, D.; Chang, C. The expression and evaluation of androgen receptor in human renal cell carcinoma. Urology 2014, 83, e19–e24. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y. Nuclear receptors link gender dimorphism of renal disease progression. Kidney Int. 2006, 70, 1889–1890. [Google Scholar] [CrossRef]
- Müller, V.; Szabo, A.; Viklicky, O. Sex hormones and gender related differences: Their influence on chronic renal allograft rejection. Kidney Int. 1999, 55, 2011–2020. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.; Ali, S.; McDonnell, B.J.; Burt, A.D.; Kirby, J.A. Chronic renal allograft dysfunction: The role of T cell-mediated tubular epithelial to mesenchymal cell transition. J. Am. Sci. Nephrol. 2004, 15, 390–397. [Google Scholar] [CrossRef]
- Araneo, B.A.; Dowell, T.; Diegel, M.; Daynes, R.A. Dihydrotestosterone exerts a depressive influence on the production of interleukin-4 (IL-4), IL-5, and γ-interferon, but not IL-2 by activated murine T cells. Blood 1991, 78, 688–699. [Google Scholar]
- Benten, W.P.M.; Lieberherr, M.; Giese, G.; Wrehlke, C.; Stamm, O.; Sekeris, C.E.; Mossmann, H.; Wunderlich, F. Functional testosterone receptors in plasma membranes of T cells. FASEB J. 1999, 13, 123–133. [Google Scholar] [CrossRef]
- Grossman, C. Possible underlying mechanisms of sexual dimorphism in the immune response, fact and hypothesis. J. Steroid Biochem. 1989, 34, 241–251. [Google Scholar] [CrossRef]
- Huynh, H. Induction of apoptosis in rat ventral prostate by finasteride is associated with alteration in MAP kinase pathways and Bcl-2 related family of proteins. Int. J. Oncol. 2002, 20, 1297–1303. [Google Scholar] [CrossRef]
- Huynh, H.; Seyam, R.M.; Brock, G.B. Reduction of central prostate weight by finasteride is associated with suppression of insulin-like growth factor I (IGF-I), IGF-I receptor and an increase in IGF binding protein 3. Cancer Res. 1998, 58, 215–218. [Google Scholar] [PubMed]
- Huynh, H.; Alpert, L.; Alaoui-Jamali, M.A.; Ng, C.Y. Co-administration of finasteride and pure anti-estrogen ICI 182,780 act synergistically in modulating the IGF system in rat prostate. J. Endocrinol. 2001, 171, 109–118. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Metcalfe, P.D.; Leslie, J.A.; Campbell, M.T.; Meldrum, D.R.; Hile, K.L.; Meldrum, KK. Testosterone exacerbates obstructive renal injury by stimulating TNF-alpha production and increasing proapoptotic and profibrotic signaling. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E435–443. [Google Scholar] [CrossRef] [PubMed]
- Verzola, D.; Gandolfo, M.T.; Salvatore, F.; Villaggio, B.; Gianiorio, F.; Traverso, P.; Deferrari, G.; Garibotto, G. Testosterone promotes apoptotic damage in human renal tubular cells. Kidney Int. 2004, 65, 1252–1261. [Google Scholar] [CrossRef]
- Verzola, D.; Villaggio, B.; Procopio, V.; Gandolfo, M.T.; Gianiorio, F.; Famà, A.; Tosetti, F.; Traverso, P.; Deferrari, G.; Garibotto, G. Androgen-mediated apoptosis of kidney tubule cells: Role of c-Jun amino terminal kinase. Biochem. Biophys. Res. Commun. 2009, 387, 531–536. [Google Scholar] [CrossRef]
- Rittmaster, R.S.; Norman, R.W.; Thomas, L.N.; Rowden, G. Evidence for atrophy and apoptosis in the prostates of men given finasteride. J. Clin. Endocrinol Metab. 1996, 81, 814–819. [Google Scholar] [PubMed]
- Sirinarumitr, K.; Sirnarumitr, T.; Johnston, S.D.; Sarkar, D.K.; Kustritz, M.V. Finasteride-induced prostatic involution by apoptosis in dogs with benign prostatic hypertrophy. Am. J. Vet. Res. 2002, 63, 495–498. [Google Scholar] [CrossRef]
- Gilleron, J.; Carette, D.; Carpentier, F.; Segretain, D.; Pointis, G. Three dimensional analysis of connexin 43 gap junction in the ex vivo rat seminiferous tubules: Short-term effects of hormonal effectors. Microsc. Res. Tech. 2009, 72, 845–855. [Google Scholar] [CrossRef]
- Hejmej, A.; Bilińska, B. A role of junction-mediated interactions in cells of the male reproductive tract: Impact of prenatal, neonatal, and prepubertal exposure to anti-androgens on adult reproduction. Histol. Histopathol. 2014, 29, 815–830. [Google Scholar]
- Yao, J.; Oite, T.; Kitamura, M. Gap junctional intercellular communication in the juxtaglomerular apparatus. Am. J. Physiol. Ren. Physiol. 2009, 296, F939–F946. [Google Scholar] [CrossRef]
- Pechere-Bertschi, A.; Burnier, M. Gonadal steroids, salt-sensitivity and renal function. Curr. Opin. Nephrol. Hypertens. 2007, 16, 16–21. [Google Scholar] [CrossRef]
- Rastaldi, M.P.; Ferrario, F.; Giardino, L.; Dell’Antonio, G.; Grillo, C.; Grillo, P.; Strutz, F.; Müller, G.A.; Colasanti, G.; D’Amico, G. Epithelial-mesenchymal transition of tubular epithelial cells in human renal biopsies. Kidney Int. 2002, 62, 137–146. [Google Scholar] [CrossRef]
- Zheng, G.; Lyons, J.G.; Tan, T.K.; Wang, Y.; Hsu, T.T.; Min, D.; Succar, L.; Rangan, G.K.; Hu, M.; Henderson, B.R.; Alexander, S.I.; Harris, D.C. Disruption of E-cadherin by matrix metalloproteinase directly mediates epithelial-mesenchymal transition downstream of transforming growth factor-β1 in renal tubular epithelial cells. Am. J. Pathol. 2009, 175, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.J.; Oh, S.; Lee, G.T.; Chung, J.; Min, K.; Yoon, J.; Kim, W.; Ryu, D.S.; Kim, I.Y.; Kang, D.I. Clinical Significance of Wnt/β-Catenin Signaling and Androgen Receptor Expression in Prostate Cancer. World J. Men’s Health 2013, 31, 36–46. [Google Scholar] [CrossRef]
- Wang, K.; Li, N.; Yeung, C.H.; Li, J.Y.; Wang, H.Y.; Cooper, TG. Oncogenic Wnt/β-catenin signaling pathways in the cancer-resistant epididymis have implications for cancer research. Mol. Hum. Reprod. 2013, 19, 57–71. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kawakami, T.; Ren, S.; Duffield, J.S. Wnt signaling in kidney diseases: Dual roles in renal injury and repair. J. Pathol. 2013, 229, 221–231. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DHT (ng/mL) | T (ng/mL) | E2 (pg/mL) | |||
|---|---|---|---|---|---|
| Control | Fin | Control | Fin | Control | Fin |
| 0.67 ± 0.03 | 0.25 ± 0.09 ** vs. Control | 1.53 ± 0.19 | 0.57 ± 0.25 ** vs. Control | 56.28 ± 9.47 | 31.33 ± 3.63 |
| Region | AR | |
|---|---|---|
| Control | Fin | |
| RC | 0.14 ± 0.12 | 0.06 ± 0.03 |
| PCT | 0.69 ± 0.32 | 0.16 ± 0.17 * |
| DCT | 0.91 ± 0.12 | 0.5 ± 0.26 * |
| Occludin | Control | Fin |
|---|---|---|
| RC mean ± SD | 100% (1) 1 ± 0 | 100% (1) 1 ± 0 |
| PCT mean ± SD | 18.5% (1); 81.5% (0) 0.185 ± 0.39 | 33.3% (1); 66.6% (0) 0.33 ± 0.48 |
| DCT mean ± SD | 58% (2); 42% (1) 1.58 ± 0.5 | 40% (2); 60 % (1) 1.4 ± 0.54 |
| Connexin 43 | ||
| RC mean ± SD | 100% (1) 1 ± 0 | 66.6% (2); 33.3% (1) 1.67 ± 0.58 *** |
| PCT mean ± SD | 24% (3); 24% (2); 52% (1) 1.71 ± 0.84 | 54.5% (3); 31.8% (2); 9.2% (1); 4.5% (0) 2.36 ± 0.84 * |
| DCT mean ± SD | 33.3% (4); 50% (3); 16.6% (2) 3.62 ± 0.71 | 80% (4); 20% (3) 3.8 ± 0.44 |
| E-cadherin | ||
| RC mean ± SD | 100% (2) 2 ± 0 | 100% (1) 1 ± 0 *** |
| PCT mean ± SD | 48,2% (2); 51.8% (1) 1.48 ± 0.51 | 7.2% (2); 17.8% (1); 75% (0) 0.32 ± 0.61 *** |
| DCT mean ± SD | 42.8% (4); 57.2% (3) 3.43 ± 0.51 | 31.6% (2); 68.4% (1) 1.3 ± 0.47 *** |
| N-cadherin | ||
| RC mean ± SD | 100% (3) 3 ± 0 | 100% (1) 1 ± 0 *** |
| PCT mean ± SD | 28,6 (1); 71.4% (0) 1.83 ± 0.41 | 21% (2); 52.6% (1); 26.4% (0) 0.95 ± 0.7 ** |
| DCT mean ± SD | 53.3% (3); 46.6% (2) 2.57 ± 0.5 | 83.3% (2); 16,6% (1) 0.28 ± 0.46 *** |
| β-catenin | ||
| RC mean ± SD | 100% (1) 1 ± 0 | 100% (1) 1 ± 0 |
| PCT mean ± SD | 45% (2); 54% (1) 1.46 ± 0.5 | 27.8% (1); 72.2% (0) 0.28 ± 0.45 |
| DCT mean ± SD | 62% (2); 28,6 (2); 9.4% (3) 3.33 ± 0.9 | 16.7% (4); 55.5% (3); 27.8% (2) 2.89 ± 0.68 |
| Apoptosis | Control | Fin |
|---|---|---|
| TUNEL+DCT | 0.77 ± 0.006 | 0.90 ± 0.0 6 * vs. Control |
| TUNEL+PCT | 0.73 ± 0.01 | 0.81 ± 0.14 |
| TUNEL+ cells per DCT | 0.18 ± 0.007 | 0.30 ± 0.07 |
| TUNEL+ cells per PCT | 0.15 ± 0.002 | 0.21 ± 0.043 |
| Proliferation | ||
| PCNA+DCT | 0.57 ± 0.14 | 0.86 ± 0.02 * vs. Control |
| PCNA+PCT | 0.35 ± 0.17 | 0.46 ± 0.16 |
| PCNA+ cells per DCT | 0.08 ± 0.002 | 0.17 ± 0.004 *** vs. Control |
| PCNA+ cells per PCT | 0.04 ± 0.01 | 0.07 ± 0.03 |
| Thickness of Collagen Fibers | Collagen Type I Fibers (µm) | Collagen Type III Fibers (µm) | ||
|---|---|---|---|---|
| Control | Fin | Control | Fin | |
| in the Interstitial Cortical Region | ||||
| Mean ± SD | 1.111 ± 0.469 | 3.298 ± 1.760 vs. Control *** | 1.561 ± 0.755 | 2.121 ± 1.154 |
| within the Renal Corpuscle | ||||
| Mean ± SD | 2.542 ± 0.974 | 2.287 ± 0.964 | 0.925 ± 0.532 | 1.426 ± 0.569 vs. Control * |
| Percentage of Area Occupied by the Collagen in Correlation to the Entire Area of the Section | ||||
| Control | Fin | |||
| Mean ± SD | 3.63 ± 1.55 | 8.56 ± 0.89 ** | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baig, M.S.; Kolasa-Wołosiuk, A.; Pilutin, A.; Safranow, K.; Baranowska-Bosiacka, I.; Kabat-Koperska, J.; Wiszniewska, B. Finasteride-Induced Inhibition of 5α-Reductase Type 2 Could Lead to Kidney Damage—Animal, Experimental Study. Int. J. Environ. Res. Public Health 2019, 16, 1726. https://doi.org/10.3390/ijerph16101726
Baig MS, Kolasa-Wołosiuk A, Pilutin A, Safranow K, Baranowska-Bosiacka I, Kabat-Koperska J, Wiszniewska B. Finasteride-Induced Inhibition of 5α-Reductase Type 2 Could Lead to Kidney Damage—Animal, Experimental Study. International Journal of Environmental Research and Public Health. 2019; 16(10):1726. https://doi.org/10.3390/ijerph16101726
Chicago/Turabian StyleBaig, Mirza Saim, Agnieszka Kolasa-Wołosiuk, Anna Pilutin, Krzysztof Safranow, Irena Baranowska-Bosiacka, Joanna Kabat-Koperska, and Barbara Wiszniewska. 2019. "Finasteride-Induced Inhibition of 5α-Reductase Type 2 Could Lead to Kidney Damage—Animal, Experimental Study" International Journal of Environmental Research and Public Health 16, no. 10: 1726. https://doi.org/10.3390/ijerph16101726
APA StyleBaig, M. S., Kolasa-Wołosiuk, A., Pilutin, A., Safranow, K., Baranowska-Bosiacka, I., Kabat-Koperska, J., & Wiszniewska, B. (2019). Finasteride-Induced Inhibition of 5α-Reductase Type 2 Could Lead to Kidney Damage—Animal, Experimental Study. International Journal of Environmental Research and Public Health, 16(10), 1726. https://doi.org/10.3390/ijerph16101726