Attachment and Survival of Escherichia coli O157:H7 on In-Shell Hazelnuts



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates and Growth Conditions
2.2. Preparation of Inoculum
2.3. Nuts
2.4. Wet Inoculation of In-Shell Hazelnuts and Storage Conditions
2.5. Preparation of Dry Inoculum
2.6. Dry Inoculation of In-Shell Hazelnuts and Storage Conditions
2.7. E. coli O157:H7 Attachment Assays
2.8. Microbiological Analysis
2.9. Real-Time qPCR Assay
2.10. Statistical Analysis and Modeling Microbial Decline
3. Results and Discussion
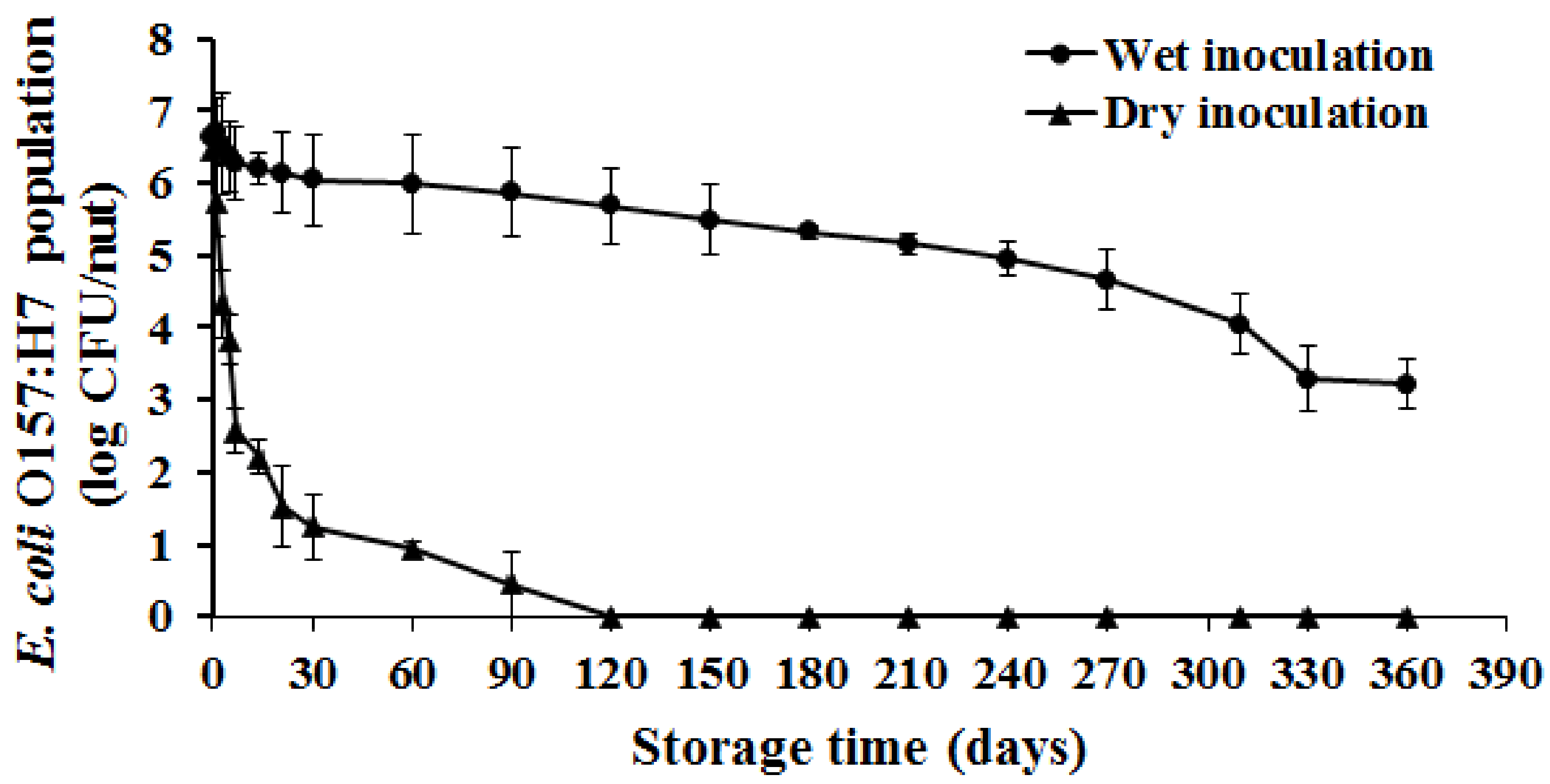
3.1. Survival of E. coli O157:H7 on Hazelnuts
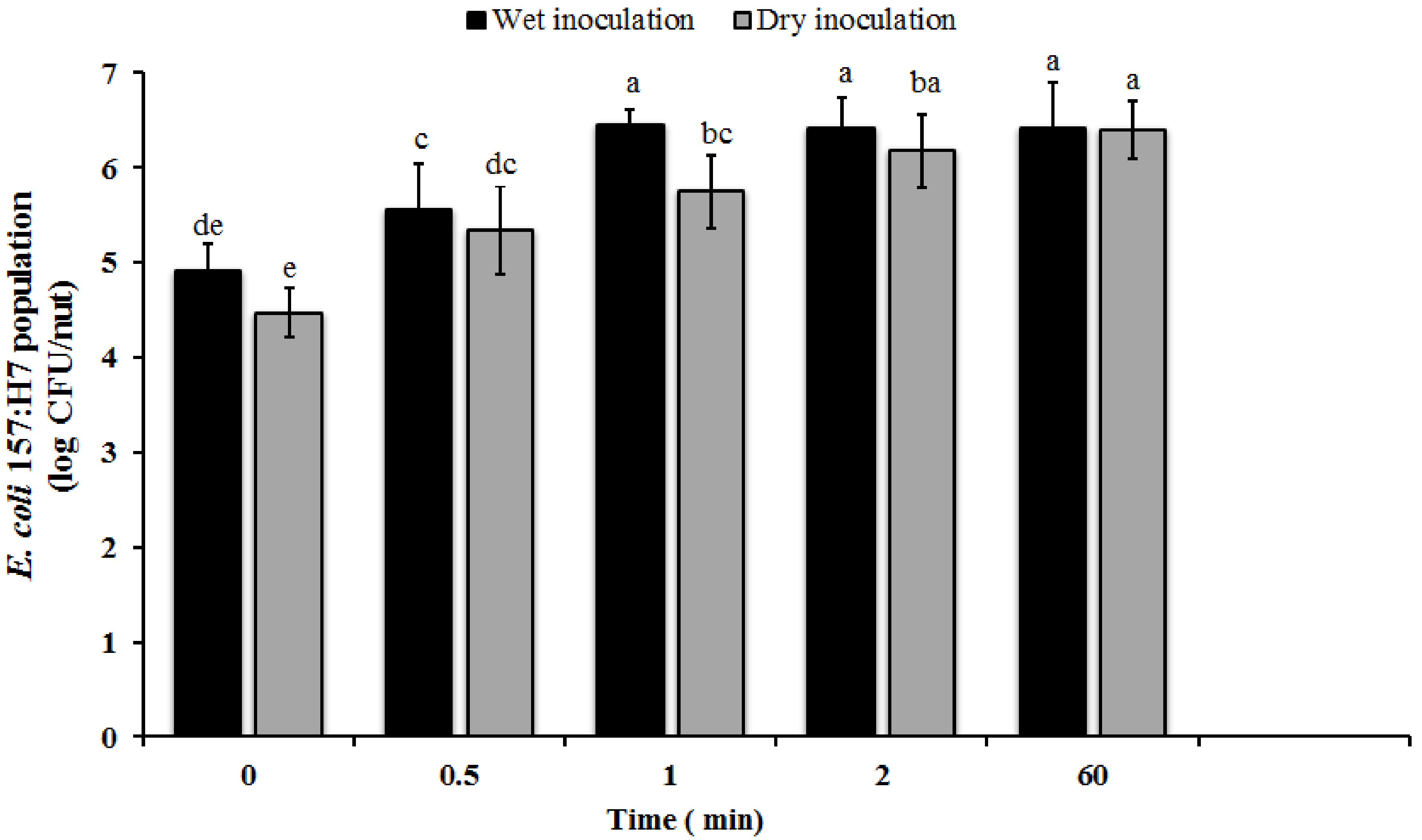
3.2. Attachment of E. coli O157 on In-Shell Hazelnuts
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Brandl, M.T. Fitness of human enteric pathogens on plants and implications for food safety. Annu. Rev. Phytopathol. 2006, 44, 367–392. [Google Scholar] [CrossRef] [PubMed]
- Aruscavage, D.; Lee, K.; Miller, S.; LeJeune, J.T. Interactions affecting the proliferation and control of human pathogens on edible plants. J. Food Sci. 2006, 71, R89–R99. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Outbreak of Salmonella serotype Enteritidis infections associated with raw almonds-United States and Canada, 2003–2004. MMWR Morb. Mortal. Wkly. Rep. 2004, 53, 484–487. [Google Scholar]
- Eglezos, S.; Huang, B.; Stuttard, E. A survey of the bacteriological quality of preroasted peanut, almond, cashew, hazelnut, and brazil nut kernels received into three Australian nut-processing facilities over a period of 3 years. J. Food Prot. 2008, 71, 402–404. [Google Scholar] [CrossRef] [PubMed]
- Little, C.L.; Jemmott, W.; Surman-Lee, S.; Hucklesby, L.; de Pinnal, E. Assessment of the microbiological safety of edible roasted nut kernels on retail sale in England, with a focus on Salmonella. J. Food Prot. 2009, 72, 853–855. [Google Scholar] [CrossRef] [PubMed]
- Little, C.L.; Rawal, N.; de Pinna, E.; McLauchlin, J. Survey of Salmonella contamination of edible nut kernels on retail sale in the UK. Food Microbiol. 2010, 27, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Riyaz-Ul-Hassan, S.; Verma, V.; Malik, A.; Qazi, G.N. Microbiological quality of walnut kernels and apple juice concentrate. World J. Microbiol. Biotechnol. 2003, 19, 845–850. [Google Scholar] [CrossRef]
- Miller, B.D.; Rigdon, C.E.; Ball, J.; Rounds, J.M.; Klos, R.F.; Brennan, B.M.; Arends, K.D.; Kennelly, P.; Hedberg, C.; Smith, K.E. Use of traceback methods to confirm the source of a multistate Escherichia coli O157:H7 outbreak due to in-shell hazelnuts. J. Food Prot. 2012, 75, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Surveillance for foodborne disease outbreaks-United States, 2009–2010. MMWR Morb. Mortal. Wkly. Rep. 2013, 62, 41–59. [Google Scholar]
- Dey, M.; Mayo, J.A.; Saville, D.; Wolyniak, C.; Klontz, K.C. Recalls of foods due to microbiological contamination classified by the U.S. Food and Drug Administration, fiscal years 2003 through 2011. J. Food Prot. 2013, 76, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Stein, R.A.; Katz, D.E. Escherichia coli, cattle and the propagation of disease. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Ingham, S.C.; Losinski, J.A.; Andrews, M.P.; Breuer, J.E.; Breuer, J.R.; Wood, T.M.; Wright, T.H. Escherichia coli contamination of vegetables grown in soils fertilized with noncomposted bovine manure: Garden-scale studies. Appl. Environ. Microbiol. 2004, 70, 6420–6427. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.; Odumeru, J. Irrigation water as source of foodborne pathogens on fruit and vegetables. J. Food Prot. 2004, 67, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J. Nut Growers Handbook. 2018. Available online: http://oregonhazelnuts.org/growers-corner/resources/nut-growers-handbook/ (accessed on 4 January 2018).
- Blessington, T.; Theofel, C.G.; Harris, L.J. A dry-inoculation method for nut kernels. Food Microbiol. 2013, 33, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Du, W.X.; Danyluk, M.D.; Harris, L.J. Efficacy of aqueous and alcohol-based quaternary ammonium sanitizers for reducing Salmonella in dusts generated in almond hulling and shelling facilities. J. Food Sci. 2010, 75, M7–M13. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Bingol, G.; Brandl, M.T.; McHugh, T.H. Review of current technologies for reduction of Salmonella populations on almonds. Food Bioprocess Technol. 2012, 5, 2046–2057. [Google Scholar] [CrossRef]
- Fischbach, J.K. Harvest and Post-Harvest Processing. 2017. Available online: https://www.midwesthazelnuts.org/processing-101.html (accessed on 4 February 2018).
- Ghirardello, D.; Contessa, C.; Valentini, N.; Zeppa, G.; Rolle, L.; Gerbi, V.; Botta, R. Effect of storage conditions on chemical and physical characteristics of hazelnut (Corylus avellana L.). Postharvest Biol. Technol. 2013, 81, 37–43. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture (USDA). Fibert/Hazelnut Kernels and Filberts in the Shell—Inspection Instructions. 2008. Available online: http://ucfoodsafety.ucdavis.edu/files/175649.pdf (accessed on 5 January 2018).
- Jung, J.; Wang, W.; McGorrin, R.J.; Zhao, Y. Moisture adsorption isotherm and storability of heazelnut inshells and kernels produced in Iregon, USA. J. Food Sci. 2018, 88, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.S.; Moore, S.C.; Tabor, R.F.; Fegan, N.; Rahman, S.; Dykes, G.A. Attachment of Salmonella strains to a plant cell wall model is modulated by surface characteristics and not by specific carbohydrate interactions. BMC Microbiol. 2016, 16, 212. [Google Scholar] [CrossRef] [PubMed]
- Fornefeld, E.; Schierstaedt, J.; Jechalke, S.; Grosch, R.; Smalla, K.; Schikora, A. Interaction between Salmonella and Plants: Potential Hosts and Vectors for Human Infection. In Current Topics in Salmonella and Salmonellosis; InTech: London, UK, 2017. [Google Scholar]
- Garrett, T.R.; Bhakoo, M.; Zhang, Z. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Dunne, W.M., Jr. Bacterial adhesion: Seen any good biofilms lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Collignon, S.; Korsten, L. Attachment and colonization by Escherichia coli O157:H7, Listeria monocytogenes, Salmonella enterica subsp. enterica serovar Typhimurium, and Staphylococcus aureus on stone fruit surfaces and Survival through a simulated commercial export chain. J. Food Prot. 2010, 73, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Sherman, D.M.; Linton, R.H.; Nielsen, S.S.; Nelson, P.E. The effects of washing and chlorine dioxide gas on survival and attachment of Escherichia coli O157:H7 to green pepper surfaces. Food Microbiol. 2000, 17, 521–533. [Google Scholar] [CrossRef]
- Barak, J.D.; Whitehand, L.C.; Charkowski, A.O. Differences in attachment of Salmonella enterica serovars and Escherichia coli O157:H7 to alfalfa sprouts. Appl. Environ. Microbiol. 2002, 68, 4758–4763. [Google Scholar] [CrossRef] [PubMed]
- Macarisin, D.; Patel, J.; Bauchan, G.; Giron, J.A.; Sharma, V.K. Role of curli and cellulose expression in adherence of Escherichia coli O157:H7 to spinach leaves. Foodborne Pathog. Dis. 2012, 9, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Fink, R.C.; Black, E.P.; Hou, Z.; Sugawara, M.; Sadowsky, M.J.; Diez-Gonzalez, F. Transcriptional responses of Escherichia coli K-12 and O157:H7 associated with lettuce leaves. Appl. Environ. Microbiol. 2012, 78, 1752–1764. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Elcott, E.; Chen, J. Differentual attachment of Salmonella enterica and Enteroheorrhagic Escherichia coli to alfalfa, fenugreek, lettuce and tomato seeds. Appl. Environ. Microbiol. 2017, 88, e03170-16. [Google Scholar]
- Yaron, S.; Romling, U. Biofilm formation by enteric pathogens and its role in plant colonization and persistence. Microb. Biotechnol. 2014, 7, 496–516. [Google Scholar] [CrossRef] [PubMed]
- Beuchat, L.R.; Heaton, E.K. Salmonella survival on pecans as influenced by processing and storage conditions. Appl. Microbiol. 1975, 29, 795–801. [Google Scholar] [PubMed]
- Frelka, J.C.; Davidson, G.R.; Harris, L.J. Changes in aerobic plate and Escherichia coli-coliform Counts and in populations of inoculated foodborne pathogens on inshell walnuts during storage. J. Food Prot. 2016, 79, 1143–1153. [Google Scholar] [CrossRef] [PubMed]
- Kimber, M.A.; Kaur, H.; Wang, L.; Danyluk, M.D.; Harris, L.J. Survival of Salmonella, Escherichia coli O157:H7, and Listeria monocytogenes on inoculated almonds and pistachios stored at −19, 4, and 24 degrees C. J. Food Prot. 2012, 75, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Hokunan, H.; Koyama, K.; Hasegawa, M.; Kawamura, S.; Koseki, S. Survival Kinetics of Salmonella enterica and Enterohemorrhagic Escherichia coli on a Plastic Surface at Low Relative Humidity and on Low-Water Activity Foods. J. Food Prot. 2016, 79, 1680–1692. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, A.R.; Harris, L.J. Growth of Salmonella Enteritidis phage type 30 in almond hull and shell slurries and survival in drying almond hulls. J. Food Prot. 2006, 69, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Brar, P.K.; Proano, L.G.; Friedrich, L.M.; Harris, L.J.; Danyluk, M.D. Survival of Salmonella, Escherichia coli O157:H7, and Listeria monocytogenes on raw peanut and pecan kernels stored at −24, 4, and 22 degrees C. J. Food Prot. 2015, 78, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Blessington, T.; Theofel, C.G.; Mitcham, E.J.; Harris, L.J. Survival of foodborne pathogens on inshell walnuts. Int. J. Food Microbiol. 2013, 166, 341–348. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Bacteriological Analytical Manual (BAM) Online. 1998. Available online: https://www.fda.gov/Food/FoodScienceResearch/LaboratoryMethods/ucm070080.htm (accessed on 5 February 2018).
- Barak, J.D.; Gorski, L.; Naraghi-Arani, P.; Charkowski, A.O. Salmonella enterica virulence genes are required for bacterial attachment to plant tissue. Appl. Environ. Microbiol. 2005, 71, 5685–5691. [Google Scholar] [CrossRef] [PubMed]
- Carey, C.M.; Kostrzynska, M.; Thompson, S. Escherichia coli O157:H7 stress and virulence gene expression on Romaine lettuce using comparative real-time PCR. J. Microbiol. Methods 2009, 77, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ali, G.S.; Ouellette, L.M.; Henderson, S.T.; Whittam, T.S.; Manning, S.D. Differences in adherence and virulence gene expression between two outbreak strains of enterohaemorrhagic Escherichia coli O157:H7. Microbiology 2010, 156, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Hu, L.; Melka, D.; Wang, H.; Laasri, A.; Brown, E.W.; Strain, E.; Allard, M.; Bunning, V.K.; Musser, S.M.; et al. Prevalence of Salmonella in Cashews, Hazelnuts, Macadamia Nuts, Pecans, Pine Nuts, and Walnuts in the United States. J. Food Prot. 2017, 80, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Marcus, K.A.; Amling, H.J. Escherichia coli field contamination of pecan nuts. Appl. Microbiol. 1973, 26, 279–281. [Google Scholar] [PubMed]
- Miksch, R.R.; Leek, J.; Myoda, S.; Nguyen, T.; Tenney, K.; Svidenko, V.; Greeson, K.; Samadpour, M. Prevalence and counts of Salmonella and enterohemorrhagic Escherichia coli in raw, shelled runner peanuts. J. Food Prot. 2013, 76, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, S.; Aramini, J.; Ciebin, B.; Farrar, J.A.; Ahmed, R.; Middleton, D.; Chandran, A.U.; Harris, L.J.; Howes, M.; Chan, E.; et al. Salmonella Enteritidis PT30 outbreak investigation working group an international outbreak of salmonellosis associated with raw almonds contaminated with a rare phage type of Salmonella enteritidis. J. Food Prot. 2005, 68, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, A.R.; Danyluk, M.D.; Mandrell, R.E.; Harris, L.J. Isolation of Salmonella Enteritidis phage type 30 from a single almond orchard over a 5-year period. J. Food Prot. 2007, 70, 1784–1789. [Google Scholar] [CrossRef] [PubMed]
- Keller, S. E. Tree fruits and nuts: Outbreaks, contamination, sources, prevention, and remediation. In The Produce Contamination Problem: Causes and Solutions; Sapers, G.M., Solomon, E.B., Matthews, K.R., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 249–269. [Google Scholar]
- Blessington, T.; Mitcham, E.J.; Harris, L.J. The survival of Salmonella Enteritidis on walnuts during and after walnut hulling. In Proceedings of the International Association for Food Protection Annual Meeting, Columbus, OH, USA, 3–6 August 2008. Abstract P1-32. [Google Scholar]
- Shaw, R.K.; Berger, C.N.; Feys, B.; Knutton, S.; Pallen, M.J.; Frankel, G. Enterohemorrhagic Escherichia coli exploits EspA filaments for attachment to salad leaves. Appl. Environ. Microbiol. 2008, 74, 2908–2914. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Jeter, C.; Langley, W.; Matthysse, A.G. Differential binding of Escherichia coli O157:H7 to alfalfa, human epithelial cells, and plastic is mediated by a variety of surface structures. Appl. Environ. Microbiol. 2005, 71, 8008–8015. [Google Scholar] [CrossRef] [PubMed]
- Matthysse, A.G.; Deora, R.; Mishra, M.; Torres, A.G. Polysaccharides cellulose, poly-beta-1,6-N-acetyl-d-glucosamine, and colanic acid are required for optimal binding of Escherichia coli O157:H7 strains to alfalfa sprouts and K-12 strains to plastic but not for binding to epithelial cells. Appl. Environ. Microbiol. 2008, 74, 2384–2390. [Google Scholar] [CrossRef] [PubMed]
- Jeter, C.; Matthysse, A.G. Characterization of the binding of diarrheagenic strains of E. coli to plant surfaces and the role of curli in the interaction of the bacteria with alfalfa sprouts. Mol. Plant Microbe Interact. 2005, 18, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Saldana, Z.; Sanchez, E.; Xicohtencatl-Cortes, J.; Puente, J.L.; Giron, J.A. Surface structures involved in plant stomata and leaf colonization by shiga-toxigenic Escherichia coli O157:H7. Front. Microbiol. 2011, 2, 119. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, L.; Muyyarikkandy, M.S.; Brown, S.R.B.; Amalaradjou, M.A. Attachment and Survival of Escherichia coli O157:H7 on In-Shell Hazelnuts. Int. J. Environ. Res. Public Health 2018, 15, 1122. https://doi.org/10.3390/ijerph15061122
Feng L, Muyyarikkandy MS, Brown SRB, Amalaradjou MA. Attachment and Survival of Escherichia coli O157:H7 on In-Shell Hazelnuts. International Journal of Environmental Research and Public Health. 2018; 15(6):1122. https://doi.org/10.3390/ijerph15061122
Chicago/Turabian StyleFeng, Lingyu, Muhammed S. Muyyarikkandy, Stephanie R. B. Brown, and Mary Anne Amalaradjou. 2018. "Attachment and Survival of Escherichia coli O157:H7 on In-Shell Hazelnuts" International Journal of Environmental Research and Public Health 15, no. 6: 1122. https://doi.org/10.3390/ijerph15061122
APA StyleFeng, L., Muyyarikkandy, M. S., Brown, S. R. B., & Amalaradjou, M. A. (2018). Attachment and Survival of Escherichia coli O157:H7 on In-Shell Hazelnuts. International Journal of Environmental Research and Public Health, 15(6), 1122. https://doi.org/10.3390/ijerph15061122