Aedes Mosquitoes and Aedes-Borne Arboviruses in Africa: Current and Future Threats

 ,
,  , , , and
, , , and
Abstract
:1. Introduction and Overview
2. The Burden of Aedes-Borne Viral Diseases in Africa
3. Range and Distribution of Aedes Vector Species in Africa
4. Vectorial Capacity of Different Aedes Species in Africa
4.1. Aedes aegypti vs. Aedes albopictus
4.2. Sylvatic vs. Domestic Forms of Aedes aegypti
4.3. Other Aedes Species and Potential for Emergent Transmission Cycles
5. Vector Surveillance and Control of Aedes species in Africa
5.1. Integrating Aedes Species Control with Malaria Vector Control: Uniquely African Opportunities?
5.2. Insecticide Resistance and Challenges to Control
6. Conclusions
Major Knowledge Gaps and Recommendations
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaenisch, T.; Junghans, T.; Wills, B.; Brady, O.J.; Eckerle, I.; Farlow, A.; Hay, S.I.; McCall, P.J.; Messina, J.P.; Ofula, V.; et al. Dengue expansion in Africa-not recognized or not happening? Emerg. Infect. Dis. 2014, 20. [Google Scholar] [CrossRef] [PubMed]
- Chan, M. Yellow fever: The resurgence of a forgotten disease. Lancet 2016, 387, 2165–2166. [Google Scholar] [CrossRef]
- Garske, T.; Van Kerkhove, M.D.; Yactayo, S.; Ronveaux, O.; Lewis, R.F.; Staples, J.E.; Perea, W.; Ferguson, N.M. Yellow fever in Africa: Estimating the burden of disease and impact of mass vaccination from outbreak and serological data. PLoS Med. 2014, 11, e1001638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jentes, E.S.; Poumerol, G.; Gershman, M.D.; Hill, D.R.; Lemarchand, J.; Lewis, R.F.; Staples, J.E.; Tomori, O.; Wilder-Smith, A.; Monath, T.P. The revised global yellow fever risk map and recommendations for vaccination, 2010: Consensus of the informal WHO working group on geographic risk for yellow fever. Lancet Infect. Dis. 2011, 11, 622–632. [Google Scholar] [CrossRef]
- Stoler, J.; Al Dashti, R.; Anto, F.; Fobil, J.N.; Awandare, G.A. Deconstructing “malaria”: West Africa as the next front for dengue fever surveillance and control. Acta Trop. 2014, 134, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Kindhauser, M.K.; Allen, T.; Frank, V.; Santhana, R.S.; Dye, C. Zika: The origin and spread of a mosquito-borne virus. Bull. World Health Organ. 2016, 94, 675C–686C. [Google Scholar] [CrossRef] [PubMed]
- Burt, F.J.; Chen, W.; Miner, J.J.; Lenschow, D.J.; Merits, A.; Schnettler, E.; Kohl, A.; Rudd, P.A.; Taylor, A.; Herrero, L.J.; et al. Chikungunya virus: An update on the biology and pathogenesis of this emerging pathogen. Lancet Infect. Dis. 2017, 17, e107–e117. [Google Scholar] [CrossRef]
- Barrett, A.D.; Higgs, S. Yellow fever: A disease that has yet to be conquered. Ann. Rev. Entomol. 2007, 52, 209–229. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C.; Twiddy, S.S. The origin, emergence and evolutionary genetics of dengue virus. Infect. Genet Evol. 2003, 3, 19–28. [Google Scholar] [CrossRef]
- Mattingly, P.F. Genetical aspects of the Aedes aegypti problem: I. Taxonomy and bionomics. Ann. Trop. Med. Parasitol. 1957, 51, 392–408. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.; Sylla, M.; Goss, L.; Burugu, M.W.; Sang, R.; Kamau, L.W.; Kenya, E.U.; Bosio, C.; Munoz Mde, L.; Sharakova, M.; et al. Dual African origins of global Aedes aegypti s.l. populations revealed by mitochondrial DNA. PLoS Negl. Trop. Dis. 2013, 7, e2175. [Google Scholar] [CrossRef] [PubMed]
- Tabachnick, W.J. Evolutionary genetics and arthropod-borne disease: The yellow fever mosquito. Am. Entomol. 1991, 37, 14–26. [Google Scholar] [CrossRef]
- World Health Organization. Yellow fever: Rapid field entomological assessment during yellow fever outbreaks in Africa. In Methodological Field Approaches for Scientists with a Basic Background in Entomology; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Huang, X.Y.; Liu, Z.Y.; Zhang, F.; Zhu, X.L.; Yu, J.Y.; Ji, X.; Xu, Y.P.; Li, G.; Li, C.; et al. A single mutation in the PRM protein of Zika virus contributes to fetal microcephaly. Science 2017, 358, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, T.; Foy, B.D.; Marques, E.T.A.; Ebel, G.D.; Weger-Lucarelli, J. Mosquito-borne and sexual transmission of Zika virus: Recent developments and future directions. Virus Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef] [PubMed]
- Alirol, E.; Getaz, L.; Stoll, B.; Chappuis, F.; Loutan, L. Urbanisation and infectious diseases in a globalised world. Lancet Infect. Dis. 2011, 11, 131–141. [Google Scholar] [CrossRef]
- World Bank, Geneva. Gross Domestic Product 2016. Available online: databank.worldbank.org/data/download/GDP.pdf (accessed on 26 January 2018).
- Weaver, S.C. Emergence of epidemic Zika virus transmission and congenital Zika syndrome: Are recently evolved traits to blame? mBio 2017, 8, e02063-16. [Google Scholar] [CrossRef] [PubMed]
- Wetsman, N. The missing pieces: Lack of Zika data from Africa complicates search for answers. Nat. Med. 2017, 23, 904–906. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Kraemer, M.U.; Brady, O.J.; Pigott, D.M.; Shearer, F.M.; Weiss, D.J.; Golding, N.; Ruktanonchai, C.W.; Gething, P.W.; Cohn, E.; et al. Mapping global environmental suitability for Zika virus. eLife 2016, 5, e15272. [Google Scholar] [CrossRef] [PubMed]
- Nsoesie, E.O.; Kraemer, M.U.; Golding, N.; Pigott, D.M.; Brady, O.J.; Moyes, C.L.; Johansson, M.A.; Gething, P.W.; Velayudhan, R.; Khan, K.; et al. Global distribution and environmental suitability for chikungunya virus, 1952 to 2015. Eurosurveill. Eur. Commun. Dis. Bull. 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Shearer, F.; Longbottom, J.; Browne, A.; Pigott, D.M.; Brady, O.J.; Kraemer, M.U.G.; Marinho, F.; Yactayo, S.; Valdelaine, E.M.; Aglaer, A.; et al. Existing and potential infection risk zones of yellow fever worldwide: A modelling analysis. Lancet Glob. Health 2018. [Google Scholar] [CrossRef]
- Messina, J.P.; Brady, O.J.; Pigott, D.M.; Brownstein, J.S.; Hoen, A.G.; Hay, S.I. A global compendium of human dengue virus occurrence. Sci. Data 2014, 1, 140004. [Google Scholar] [CrossRef] [PubMed]
- Patching, H.M.; Hudson, L.M.; Cooke, W.; Garcia, A.J.; Hay, S.I.; Roberts, M.; Moyes, C.L. A supervised learning process to validate online disease reports for use in predictive models. Big Data 2015, 3, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Shearer, F.M.; Moyes, C.L.; Pigott, D.M.; Brady, O.J.; Marinho, F.; Deshpande, A.; Longbottom, J.; Browne, A.J.; Kraemer, M.U.G.; O’Reilly, K.M.; et al. Global yellow fever vaccination coverage from 1970 to 2016: An adjusted retrospective analysis. Lancet Infect. Dis. 2017, 17, 1209–1217. [Google Scholar] [CrossRef]
- Gabor, J.J.; Schwarz, N.G.; Esen, M.; Kremsner, P.G.; Grobusch, M.P. Dengue and chikungunya seroprevalence in Gabonese infants prior to major outbreaks in 2007 and 2010: A sero-epidemiological study. Travel Med. Infect. Dis. 2016, 14, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Ochieng, C.; Ahenda, P.; Vittor, A.Y.; Nyoka, R.; Gikunju, S.; Wachira, C.; Waiboci, L.; Umuro, M.; Kim, A.A.; Nderitu, L. Seroprevalence of infections with dengue, rift valley fever and chikungunya viruses in Kenya, 2007. PLoS ONE 2015, 10, e0132645. [Google Scholar] [CrossRef] [PubMed]
- Gake, B.; Vernet, M.A.; Leparc-Goffart, I.; Drexler, J.; Gould, E.A.; Gallian, P.; Lamballerie, X. Low seroprevalence of Zika virus in Cameroonian blood donors. Braz. J. Infect. Dis. 2017, 21, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.; Mallewa, M. Dengue and other emerging flaviviruses. J. Infect. 2001, 42, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.; Shearer, F.M.; Brady, O.J.; Messina, J.P.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; et al. The global compendium of Aedes aegypti and Ae. albopictus occurrence. Sci. Data 2015, 2. [Google Scholar] [CrossRef] [PubMed]
- Agwu, E.J.; Igbinosa, I.B.; Isaac, C. Entomological assessment of yellow fever-epidemic risk indices in Benue State, Nigeria, 2010–2011. Acta Trop. 2016, 161, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Ayorinde, A.; Oboh, B.; Oduola, A.; Otubanjo, O. The insecticide susceptibility status of Aedes aegypti (Diptera: Culicidae) in farm and nonfarm sites of Lagos State, Nigeria. J. Insect Sci. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Benallal, K.E.; Allal-Ikhlef, A.; Benhamouda, K.; Schaffner, F.; Harrat, Z. First report of Aedes (Stegomyia) albopictus (Diptera: Culicidae) in Oran, West of Algeria. Acta Trop. 2016, 164, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Bennouna, A.; Balenghien, T.; El Rhaffouli, H.; Schaffner, F.; Garros, C.; Gardes, L.; Lhor, Y.; Hammoumi, S.; Chlyeh, G.; Fassi, F.O. First record of Stegomyia albopicta (=Aedes albopictus) in Morocco: A major threat to public health in North Africa? Med. Vet. Entomol. 2017, 31, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Ongoing dengue epidemic—Angola. Morb. Mortal. Wkly. Rep. 2013, 62, 504–507. [Google Scholar]
- Chepkorir, E.; Lutomiah, J.; Mutisya, J.; Mulwa, F.; Limbaso, K.; Orindi, B.; Sang, R. Vector competence of Aedes aegypti populations from Kilifi and Nairobi for dengue 2 virus and the influence of temperature. Parasites Vectors 2014, 7, 435. [Google Scholar] [CrossRef] [PubMed]
- Da Moura, A.J.F.; Santos, M.A.V.d.; Oliveira, C.M.F.; Guedes, D.R.D.; de Carvalho-Leandro, D.; Brito, M.L.d.; Rocha, H.D.R.; Gómez, L.F.; Ayres, C.F.J. Vector competence of the Aedes aegypti population from Santiago Island, Cape Verde, to different serotypes of dengue virus. Parasites Vectors 2015, 8, 114. [Google Scholar] [CrossRef] [PubMed]
- Dia, I.; Diagne, C.T.; Ba, Y.; Diallo, D.; Konate, L.; Diallo, M. Insecticide susceptibility of Aedes aegypti populations from Senegal and Cape Verde Archipelago. Parasites Vectors 2012, 5, 238. [Google Scholar] [CrossRef] [PubMed]
- Donpedro, K.N.; Adegbite, T.O. Nuvan resistance in a field strain of Aedes aegypti (L.) in Lagos, Nigeria. Environ. Pollut. Ser. A Ecol. Biol. 1985, 38, 19–29. [Google Scholar] [CrossRef]
- Gaye, A.; Faye, O.; Diagne, C.T.; Faye, O.; Diallo, D.; Weaver, S.C.; Sall, A.A.; Diallo, M. Oral susceptibility of Aedes aegypti (Diptera: Culicidae) from Senegal for dengue serotypes 1 and 3 viruses. Trop. Med. Int. Health 2014, 19, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Guindo-Coulibaly, N.; Adja, A.M.; Koffi, A.A.; Diakité, N.R.; Ahoua Alou, P.L.; Bassa, K.F.; N’Goran, K.E. Insecticides susceptibility of two distinct morphologies at larval stage of Aedes aegypti (Diptera: Culicidae) from Abidjan (Côte d’Ivoire). Eur. J. Sci. Res. 2014, 126, 434–443. [Google Scholar]
- Hertz, J.T.; Lyaruu, L.J.; Ooi, E.E.; Mosha, F.W.; Crump, J.A. Distribution of Aedes mosquitoes in the Kilimanjaro region of northern Tanzania. Pathog. Glob. Health 2016, 110, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Husham, A.O.; Abdalmagid, M.A.; Brair, M. Status susceptibility of dengue vector; Aedes aegypti to different groups of insecticides in Port Sudan City—Red Sea. Sudan. J. Public Health 2010, 10, 199–202. [Google Scholar]
- Kamgang, B.; Marcombe, S.; Chandre, F.; Nchoutpouen, E.; Nwane, P.; Etang, J.; Corbel, V.; Paupy, C. Insecticide susceptibility of Aedes aegypti and Aedes albopictus in Central Africa. Parasites Vectors 2010, 4, 79. [Google Scholar] [CrossRef] [PubMed]
- Kampango, A.; Abilio, A.P. The Asian tiger hunts in Maputo city-the first confirmed report of Aedes (Stegomyia) albopictus (Skuse, 1895) in Mozambique. Parasites Vectors 2016, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Kawada, H.; Higa, Y.; Futami, K.; Muranami, Y.; Kawashima, E.; Osei, J.H.; Sakyi, K.Y.; Dadzie, S.; de Souza, D.K.; Appawu, M.; et al. Discovery of point mutations in the voltage-gated sodium channel from African Aedes aegypti populations: Potential phylogenetic reasons for gene introgression. PLoS Negl. Trop. Dis. 2016, 10, e0004780. [Google Scholar] [CrossRef] [PubMed]
- Konan, L.Y.; Coulibaly, I.Z.; Kone, B.A.; Ziogba, J.T.; Diallo, A.; Ekra, D.K.; Traoré, K.S.; Doannio, M.C.J.; Paul, O. Aedes aegypti susceptibility to insecticide from Abidjan City, Cote d’ivoire. Vector Borne Zoonotic Dis. 2012, 12, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Konan, Y.L.; Coulibaly, Z.I.; Allali, K.B.; Tetchi, S.M.; Kone, A.B.; Coulibaly, D.; Ekra, K.D.; Doannio, J.M.; Oudéhouri-Koudou, P. Management of the yellow fever epidemic in 2010 in Seguela (Cote d’Ivoire): Value of multidisciplinary investigation. Sante Publique 2014, 26, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Lutomiah, J.; Barrera, R.; Makio, A.; Mutisya, J.; Koka, H.; Owaka, S.; Koskei, E.; Nyunja, A.; Eyase, F.; Coldren, R.; et al. Dengue outbreak in Mombasa City, Kenya, 2013–2014: Entomologic investigations. PLoS Negl. Trop. Dis. 2016, 10, e0004981. [Google Scholar] [CrossRef] [PubMed]
- Massangaie, M.; Pinto, G.; Padama, F.; Chambe, G.; da Silva, M.; Mate, I.; Chirindza, C.; Ali, S.; Agostinho, S.; Chilaule, D.; et al. Clinical and epidemiological characterization of the first recognized outbreak of dengue virus-type 2 in Mozambique, 2014. Am. J. Trop. Med. Hyg. 2016, 94, 413–416. [Google Scholar] [CrossRef] [PubMed]
- Mouchet, J. La resistance aux insecticides chez Aedes aegypti et les especes voisines. Bull. World Health Organ. 1967, 36, 569–577. [Google Scholar] [PubMed]
- Müller, G.C.; Tsabari, O.; Traore, M.M.; Traore, S.F.; Kravchenko, V.D.; Junnila, A.; Beier, J.C. First record of Aedes albopictus in inland Africa along the River Niger in Bamako and Mopti, Mali. Acta Trop. 2016, 162, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Ngoagouni, C.; Kamgang, B.; Brengues, C.; Yahouedo, G.; Paupy, C.; Nakouné, E.; Kazanji, M.; Chandre, F. Susceptibility profile and metabolic mechanisms involved in Aedes aegypti and Aedes albopictus resistant to DDT and deltamethrin in the Central African Republic. Parasites Vectors 2016, 9, 599. [Google Scholar] [CrossRef] [PubMed]
- Noden, B.H.; Musuuo, M.; Aku-Akai, L.; van der Colf, B.; Chipare, I.; Wilkinson, R. Risk assessment of flavivirus transmission in Namibia. Acta Trop. 2014, 137, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Oduola, A.O.; Obembe, A.; Adelaja, O.J.; Ande, A.T. Surveillance and insecticide susceptibility status of culicine mosquitoes in selected communities utilizing long-lasting insecticidal nets in Kwara State, Nigeria. Anim. Res. Int. 2016, 13, 2483–2491. [Google Scholar]
- Pocquet, N.; Darriet, F.; Zumbo, B.; Milesi, P.; Thiria, J.; Bernard, V.; Toty, C.; Labbé, P.; Chandre, F. Insecticide resistance in disease vectors from Mayotte: An opportunity for integrated vector management. Parasites Vectors 2014, 7, 299. [Google Scholar] [CrossRef] [PubMed]
- Reis, S.; Cornel, A.J.; Melo, M.; Pereira, H.; Loiseau, C. First record of Aedes albopictus (Skuse 1894) on Sâo Tomé island. Acta Trop. 2017, 171, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro Rocha, H.D.; Santos Paiva, M.H.; Machado Silva, N.; Araújo, A.P.; de Azevedo Camacho, D.d.R.d.; da Moura, A.J.F.; Ferrero Gómez, L.; Junqueira Ayres, C.F.; de Melo Santos, M.A.V. Susceptibility profile of Aedes aegypti from Santiago Island, Cabo Verde, to insecticides. Acta Trop. 2015, 152, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Self, L.S.; Pant, C.P. Insecticide susceptibility and resistance in populations of Anopheles gambiae, Culex fatigans and Aedes aegypti in southern Nigeria. Bull. World Health Organ. 1966, 24, 960–962. [Google Scholar]
- Lindsay, S.W.; Wilson, A.; Golding, N.; Scott, T.W.; Takken, W. Improving the built environment in urban areas to control Aedes aegypti-borne diseases. Bull. World Health Organ. 2017, 95, 607–608. [Google Scholar] [CrossRef] [PubMed]
- Talla, C.; Diallo, D.; Dia, I.; Ba, Y.; Ndione, J.; Morse, A.P.; Diop, A.; Diallo, M. Modelling hotspots of the two dominant Rift Valley fever vectors (Aedes vexans and Culex poicilipes) in Barkedji, Senegal. Parasites Vectors 2016, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites Vectors 2013, 6, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, R.; Lutomiah, J.; Said, M.; Makio, A.; Koka, H.; Koskei, E.; Nyunja, A.; Owaka, S.; Matoke-Muhia, D.; Bukachi, S.; et al. Effects of irrigation and rainfall on the population dynamics of rift valley fever and other arbovirus mosquito vectors in the epidemic-prone Tana River County, Kenya. J. Med. Entomol. 2017, 54, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Kamgang, B.; Happi, J.Y.; Boisier, P.; Njiokou, F.; Hervé, J.P.; Simard, F.; Paupy, C. Geographic and ecological distribution of the dengue and chikungunya virus vectors Aedes aegypti and Aedes albopictus in three major Cameroonian towns. Med. Vet. Entomol. 2010, 24, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Kamgang, B.; Ngoagouni, C.; Manirakiza, A.; Nakouné, E.; Paupy, C.; Kazanji, M. Temporal patterns of abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) and mitochondrial DNA analysis of Ae. albopictus in the Central African Republic. PLoS Negl. Trop. Dis. 2013, 7, e2590. [Google Scholar] [CrossRef] [PubMed]
- Kamgang, B.; Yougang, A.P.; Tchoupo, M.; Riveron, J.M.; Wondji, C. Temporal distribution and insecticide resistance profile of two major arbovirus vectors Aedes aegypti and Aedes albopictus in Yaoundé, the capital city of Cameroon. Parasites Vectors 2017, 10, 469. [Google Scholar] [CrossRef] [PubMed]
- Zahouli, J.B.Z.; Koudou, B.G.; Muller, P.; Malone, D.; Tano, Y.; Utzinger, J. Effect of land-use changes on the abundance, distribution, and host-seeking behavior of Aedes arbovirus vectors in oil palm-dominated landscapes, southeastern Cote d’Ivoire. PLoS ONE 2017, 12, e0189082. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Scott, T.W.; Gubler, D.J. Consequences of the expanding global distribution of Aedes albopictus for dengue virus transmission. PLoS Negl. Trop. Dis. 2010, 4, e646. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Forrester, N.L. Chikungunya: Evolutionary history and recent epidemic spread. Antivir. Res. 2015, 120, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Pages, F.; Peyrefitte, C.N.; Mve, M.T.; Jarjaval, F.; Brisse, S.; Iteman, I.; Gravier, P.; Nkoghe, D.; Grandadam, M. Aedes albopictus mosquito: The main vector of the 2007 chikungunya outbreak in Gabon. PLoS ONE 2009, 4, e4691. [Google Scholar] [CrossRef]
- Paupy, C.; Ollomo, B.; Kamgang, B.; Moutailler, S.; Rousset, D.; Demanou, M.; Herve, J.P.; Leroy, E.; Simard, F. Comparative role of Aedes albopictus and Aedes aegypti in the emergence of dengue and chikungunya in Central Africa. Vector Borne Zoonotic Dis. 2010, 10, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Grard, G.; Caron, M.; Mombo, I.M.; Nkoghe, D.; Mboui Ondo, S.; Jiolle, D.; Fontenille, D.; Paupy, C.; Leroy, E.M. Zika virus in Gabon (Central Africa)—2007: A new threat from Aedes albopictus? PLoS Negl. Trop. Dis. 2014, 8, e2681. [Google Scholar] [CrossRef] [PubMed]
- Paupy, C.; Kassa Kassa, F.; Caron, M.; Nkoghe, D.; Leroy, E.M. A chikungunya outbreak associated with the vector Aedes albopictus in remote villages of Gabon. Vector Borne Zoonotic Dis. 2012, 12, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Ngoagouni, C.; Kamgang, B.; Nakoune, E.; Paupy, C.; Kazanji, M. Invasion of Aedes albopictus (Diptera: Culicidae) into Central Africa: What consequences for emerging diseases? Parasites Vectors 2015, 8, 191. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Aedes aegypti—A review. Mem. Inst. Oswaldo Cruz 2013, 108 (Suppl. 1), 11–17. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; McBride, C.S.; Johnson, P.; Ritchie, S.; Paupy, C.; Bossin, H.; Lutomiah, J.; Fernandez-Salas, I.; Ponlawat, A.; Cornel, A.J.; et al. Worldwide patterns of genetic differentiation imply multiple ‘domestications’ of Aedes aegypti, a major vector of human diseases. Proc. Biol. Sci. 2011, 278, 2446–2454. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.E.; Alves, J.M.; Palmer, W.J.; Day, J.P.; Sylla, M.; Ramasamy, R.; Surendran, S.N.; Black, W.C.T.; Pain, A.; Jiggins, F.M. Population genomics reveals that an anthropophilic population of Aedes aegypti mosquitoes in West Africa recently gave rise to American and Asian populations of this major disease vector. BMC Biol. 2017, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Dickson, L.B.; Jiolle, D.; Minard, G.; Moltini-Conclois, I.; Volant, S.; Ghozlane, A.; Bouchier, C.; Ayala, D.; Paupy, C.; Moro, C.V.; et al. Carryover effects of larval exposure to different environmental bacteria drive adult trait variation in a mosquito vector. Sci. Adv. 2017, 3, e1700585. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R. Mosquitoes on the move. Science 2016, 354, 971–972. [Google Scholar] [CrossRef] [PubMed]
- Mattingly, P.F. Taxonomy of Aedes aegypti and related species. Bull. World Health Organ. 1967, 36, 552–554. [Google Scholar] [PubMed]
- McClelland, G.A.H. A worldwide survey of variation in scale pattern of the abdominal tergum of Aedes aegypti (L.) (Diptera: Culicidae). Trans. R. Entomol. Soc. Lond. 1974, 126, 239–259. [Google Scholar] [CrossRef]
- Huber, K.; Ba, Y.; Dia, I.; Mathiot, C.; Sall, A.A.; Diallo, M. Aedes aegypti in Senegal: Genetic diversity and genetic structure of domestic and sylvatic populations. Am. J. Trop. Med. Hyg. 2008, 79, 218–229. [Google Scholar] [PubMed]
- Paupy, C.; Brengues, C.; Ndiath, O.; Toty, C.; Herve, J.P.; Simard, F. Morphological and genetic variability within Aedes aegypti in Niakhar, Senegal. Infect. Genet. Evol. 2010, 10, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Sylla, M.; Bosio, C.; Urdaneta-Marquez, L.; Ndiaye, M.; Black, W.C.T. Gene flow, subspecies composition, and dengue virus-2 susceptibility among Aedes aegypti collections in Senegal. PLoS Negl. Trop. Dis. 2009, 3, e408. [Google Scholar] [CrossRef] [PubMed]
- Gloria-Soria, A.; Ayala, D.; Bheecarry, A.; Calderon-Arguedas, O.; Chadee, D.D.; Chiappero, M.; Coetzee, M.; Elahee, K.B.; Fernandez-Salas, I.; Kamal, H.A.; et al. Global genetic diversity of Aedes aegypti. Mol. Ecol. 2016, 25, 5377–5395. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; Evans, B.R.; Zheng, W.; Obas, V.; Barrera-Martinez, L.; Egizi, A.; Zhao, H.; Caccone, A.; Powell, J.R. Human impacts have shaped historical and recent evolution in Aedes aegypti, the dengue and yellow fever mosquito. Evol. Int. J. Org. Evol. 2014, 68, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Black, W.C.; Bennett, K.E.; Gorrochotegui-Escalante, N.; Barillas-Mury, C.V.; Fernandez-Salas, I.; de Lourdes Munoz, M.; Farfan-Ale, J.A.; Olson, K.E.; Beaty, B.J. Flavivirus susceptibility in Aedes aegypti. Arch. Med. Res. 2002, 33, 379–388. [Google Scholar] [CrossRef]
- Bosio, C.F.; Beaty, B.J.; Black, W.C. Quantitative genetics of vector competence for dengue-2 virus in Aedes aegypti. Am. J. Trop. Med. Hyg. 1998, 59, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Tabachnick, W.J.; Wallis, G.P.; Aitken, T.H.; Miller, B.R.; Amato, G.D.; Lorenz, L.; Powell, J.R.; Beaty, B.J. Oral infection of Aedes aegypti with yellow fever virus: Geographic variation and genetic considerations. Am. J. Trop. Med. Hyg. 1985, 34, 1219–1224. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Ba, Y.; Faye, O.; Soumare, M.L.; Dia, I.; Sall, A.A. Vector competence of Aedes aegypti populations from Senegal for sylvatic and epidemic dengue 2 virus isolated in West Africa. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Sall, A.A.; Moncayo, A.C.; Ba, Y.; Fernandez, Z.; Ortiz, D.; Coffey, L.L.; Mathiot, C.; Tesh, R.B.; Weaver, S.C. Potential role of sylvatic and domestic African mosquito species in dengue emergence. Am. J. Trop. Med. Hyg. 2005, 73, 445–449. [Google Scholar] [PubMed]
- Dickson, L.B.; Sanchez-Vargas, I.; Sylla, M.; Fleming, K.; Black, W.C. Vector competence in West African Aedes aegypti is flavivirus species and genotype dependent. PLoS Negl. Trop. Dis. 2014, 8, e3153. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Yebakima, A.; Lourenco-de-Oliveira, R.; Andriamahefazafy, B.; Correira, A.; Rodrigues, J.M.; Veiga, A.; Moreira, A.; Leparc-Goffart, I.; Grandadam, M.; et al. Oral receptivity of Aedes aegypti from Cape Verde for yellow fever, dengue, and chikungunya viruses. Vector Borne Zoonotic Dis. 2013, 13, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L. Dissecting the genetic architecture of host-pathogen specificity. PLoS Pathog. 2010, 6, e1001019. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.J.; Smithburn, K.C.; Dick, G.W.A.; Kitchen, S.F.; Lumsden, W.H.R. Implication of the mosquito Aedes (Stegomyia) africanus Theobald in the forest cycle of yellow fever in Uganda. Ann. Trop. Med. Parasitol. 1948, 42, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Hanley, K.A.; Monath, T.P.; Weaver, S.C.; Rossi, S.L.; Richman, R.L.; Vasilakis, N. Fever versus fever: The role of host and vector susceptibility and interspecific competition in shaping the current and future distributions of the sylvatic cycles of dengue virus and yellow fever virus. Infect. Genet. Evol. 2013, 19, 292–311. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.J. Mosquito-borne viruses—The need for a balanced team. In Medical Entomology Centenary; Royal Society of Tropical Medicine and Hygiene: London, UK, 1977; pp. 33–38. [Google Scholar]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed]
- Diallo, D.; Chen, R.; Diagne, C.T.; Ba, Y.; Dia, I.; Sall, A.A.; Weaver, S.C.; Diallo, M. Bloodfeeding patterns of sylvatic arbovirus vectors in southeastern Senegal. Trans. R. Soc. Trop. Med. Hyg. 2013, 107, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.J.; Williams, M.C.; Woodall, J.P.; Simpson, D.I.; Goma, L.K. Twelve isolations of Zika virus from Aedes (Stegomyia) africanus (Theobald) taken in and above a Uganda forest. Bull. World Health Organ. 1964, 31, 57–69. [Google Scholar] [PubMed]
- Ngoagouni, C.; Kamgang, B.; Manirakiza, A.; Nangouma, A.; Paupy, C.; Nakoune, E.; Kazanji, M. Entomological profile of yellow fever epidemics in the Central African Republic, 2006–2010. Parasites Vectors 2012, 5, 175. [Google Scholar] [CrossRef] [PubMed]
- Diallo, D.; Sall, A.A.; Buenemann, M.; Chen, R.; Faye, O.; Diagne, C.T.; Faye, O.; Ba, Y.; Dia, I.; Watts, D.; et al. Landscape ecology of sylvatic chikungunya virus and mosquito vectors in southeastern Senegal. PLoS Negl. Trop. Dis. 2012, 6, e1649. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Ba, Y.; Sall, A.A.; Diop, O.M.; Ndione, J.A.; Mondo, M.; Girault, L.; Mathiot, C. Amplification of the sylvatic cycle of dengue virus type 2, Senegal, 1999–2000: Entomologic findings and epidemiologic considerations. Emerg. Infect. Dis. 2003, 9, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Kuno, G.; Mackenzie, J.S.; Junglen, S.; Hubalek, Z.; Plyusnin, A.; Gubler, D.J. Vertebrate reservoirs of arboviruses: Myth, synonym of amplifier, or reality? Viruses 2017, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Diagne, C.T.; Diallo, D.; Faye, O.; Ba, Y.; Faye, O.; Gaye, A.; Dia, I.; Faye, O.; Weaver, S.C.; Sall, A.A.; et al. Potential of selected Senegalese Aedes spp. mosquitoes (Diptera: Culicidae) to transmit Zika virus. BMC Infect. Dis. 2015, 15, 492. [Google Scholar] [CrossRef] [PubMed]
- Diallo, D.; Sall, A.A.; Diagne, C.T.; Faye, O.; Faye, O.; Ba, Y.; Hanley, K.A.; Buenemann, M.; Weaver, S.C.; Diallo, M. Zika virus emergence in mosquitoes in southeastern Senegal, 2011. PLoS ONE 2014, 9, e109442. [Google Scholar] [CrossRef] [PubMed]
- Epelboin, Y.; Talaga, S.; Epelboin, L.; Dusfour, I. Zika virus: An updated review of competent or naturally infected mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0005933. [Google Scholar] [CrossRef] [PubMed]
- Althouse, B.M.; Vasilakis, N.; Sall, A.A.; Diallo, M.; Weaver, S.C.; Hanley, K.A. Potential for Zika virus to establish a sylvatic transmission cycle in the Americas. PLoS Negl. Trop. Dis. 2016, 10, e0005055. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Chevillon, C.; Albright, R.G.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Scott, T.W. Genetic specificity and potential for local adaptation between dengue viruses and mosquito vectors. BMC Evol. Biol. 2009, 9, 160. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, G.; Sang, R.C.; Guerbois, M.; Taracha, E.L.N.; Weaver, S.C. Enzootic circulation of chikungunya virus in East Africa: Serological evidence in non-human Kenyan primates. Am. J. Trop. Med. Hyg. 2017, 97, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Achee, N.L.; Gould, F.; Perkins, T.A.; Reiner, R.C., Jr.; Morrison, A.C.; Ritchie, S.A.; Gubler, D.J.; Teyssou, R.; Scott, T.W. A critical assessment of vector control for dengue prevention. PLoS Negl. Trop. Dis. 2015, 9, e0003655. [Google Scholar] [CrossRef] [PubMed]
- Bowman, L.R.; Donegan, S.; McCall, P.J. Is dengue vector control deficient in effectiveness or evidence? Systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2016, 10, e0004551. [Google Scholar] [CrossRef] [PubMed]
- Mahaffy, A.F. The yellow fever situation in Africa. Bull. World Health Organ. 1954, 11, 319–324. [Google Scholar] [PubMed]
- Gubler, D.J. Dengue, urbanization and globalization: The unholy trinity of the 21st century. Trop. Med. Health 2011, 39, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.D. Yellow fever in Angola and beyond--the problem of vaccine supply and demand. N. Engl. J. Med. 2016, 375, 301–303. [Google Scholar] [CrossRef] [PubMed]
- Godoi, I.P.; Lemos, L.L.; de Araujo, V.E.; Bonoto, B.C.; Godman, B.; Guerra Junior, A.A. CYD-TDV dengue vaccine: Systematic review and meta-analysis of efficacy, immunogenicity and safety. J. Comp. Eff. Res. 2017, 6, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Runge-Ranzinger, S.; McCall, P.J.; Kroeger, A.; Horstick, O. Dengue disease surveillance: An updated systematic literature review. Trop. Med. Int. Health 2014, 19, 1116–1160. [Google Scholar] [CrossRef] [PubMed]
- Demanou, M.; Pouillot, R.; Grandadam, M.; Boisier, P.; Kamgang, B.; Herve, J.P.; Rogier, C.; Rousset, D.; Paupy, C. Evidence of dengue virus transmission and factors associated with the presence of anti-dengue virus antibodies in humans in three major towns in Cameroon. PLoS Negl. Trop. Dis. 2014, 8, e2950. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.K.; Musoke, S.; Ocheng, D.; Gichogo, A.; Rees, P.H. Dengue-2 virus in Kenya. Lancet 1982, 2, 208–209. [Google Scholar] [CrossRef]
- Vu, D.M.; Mutai, N.; Heath, C.J.; Vulule, J.M.; Mutuku, F.M.; Ndenga, B.A.; LaBeaud, A.D. Unrecognized dengue virus infections in children, western Kenya, 2014-2015. Emerg. Infect. Dis. 2017, 23, 1915–1917. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.; Samuel, M.; Maoz, D.; Runge-Ranzinger, S.; Boyce, R.; Toledo, J.; Velayudhan, R.; Horstick, O. Dengue data and surveillance in Tanzania: A systematic literature review. Trop. Med. Int. Health 2017, 22, 960–970. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Strategy for Dengue Prevention and Control, 2012–2020; World Health Organization: Geneva, Switzerland, 2012; p. 43. [Google Scholar]
- Special Programme for Research and Training in Tropical Diseases (TDR), World Health Organization. Technical Handbook for Dengue Surveillance, Dengue Outbreak Prediction/Detection and Outbreak Response (Model Contingency Plan); World Health Organization: Geneva, Switzerland, 2016; p. 92. [Google Scholar]
- Brady, O.J.; Smith, D.L.; Scott, T.W.; Hay, S.I. Dengue disease outbreak definitions are implicitly variable. Epidemics 2015, 11, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Bowman, L.R.; Runge-Ranzinger, S.; McCall, P.J. Assessing the relationship between vector indices and dengue transmission: A systematic review of the evidence. PLoS Negl. Trop. Dis. 2014, 8, e2848. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. World Malaria Report 2017; World Health Organization: Geneva, Switzerland, 2017; p. 196. [Google Scholar]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Ponlawat, A.; Harrington, L.C. Blood feeding patterns of Aedes aegypti and Aedes albopictus in Thailand. J. Med. Entomol. 2005, 42, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.W.; Takken, W. Feeding strategies of anthropophilic mosquitoes result in increased risk of pathogen transmission. Trends Parasitol. 2012, 28, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A.A. The invasive mosquito species Aedes albopictus: Current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Chadee, D.D. Resting behaviour of Aedes aegypti in Trinidad: With evidence for the re-introduction of indoor residual spraying (IRS) for dengue control. Parasites Vectors 2013, 6, 255. [Google Scholar] [CrossRef] [PubMed]
- Dzul-Manzanilla, F.; Ibarra-Lopez, J.; Bibiano Marin, W.; Martini-Jaimes, A.; Leyva, J.T.; Correa-Morales, F.; Huerta, H.; Manrique-Saide, P.; Vazquez-Prokopec, G.M. Indoor resting behavior of Aedes aegypti (Diptera: Culicidae) in Acapulco, Mexico. J. Med. Entomol. 2017, 54, 501–504. [Google Scholar] [PubMed]
- Perich, M.J.; Davila, G.; Turner, A.; Garcia, A.; Nelson, M. Behavior of resting Aedes aegypti (Culicidae: Diptera) and its relation to ultra-low volume adulticide efficacy in Panama City, Panama. J. Med. Entomol. 2000, 37, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Esquivel, C.; Lenhart, A.; del Rio, R.; Leza, M.M.; Estrugo, M.; Chalco, E.; Casanova, W.; Miranda, M.A. The impact of indoor residual spraying of deltamethrin on dengue vector populations in the Peruvian Amazon. Acta Trop. 2016, 154, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Prokopec, G.M.; Medina-Barreiro, A.; Che-Mendoza, A.; Dzul-Manzanilla, F.; Correa-Morales, F.; Guillermo-May, G.; Bibiano-Marin, W.; Uc-Puc, V.; Geded-Moreno, E.; Vadillo-Sanchez, J.; et al. Deltamethrin resistance in Aedes aegypti results in treatment failure in Merida, Mexico. PLoS Negl. Trop. Dis. 2017, 11, e0005656. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Prokopec, G.M.; Montgomery, B.L.; Horne, P.; Clennon, J.A.; Ritchie, S.A. Combining contact tracing with targeted indoor residual spraying significantly reduces dengue transmission. Sci. Adv. 2017, 3, e1602024. [Google Scholar] [CrossRef] [PubMed]
- Kigozi, R.; Baxi, S.M.; Gasasira, A.; Sserwanga, A.; Kakeeto, S.; Nasr, S.; Rubahika, D.; Dissanayake, G.; Kamya, M.R.; Filler, S.; et al. Indoor residual spraying of insecticide and malaria morbidity in a high transmission intensity area of Uganda. PLoS ONE 2012, 7, e42857. [Google Scholar] [CrossRef] [PubMed]
- Kirby, M.J.; Ameh, D.; Bottomley, C.; Green, C.; Jawara, M.; Milligan, P.J.; Snell, P.C.; Conway, D.J.; Lindsay, S.W. Effect of two different house screening interventions on exposure to malaria vectors and on anaemia in children in The Gambia: A randomised controlled trial. Lancet 2009, 374, 998–1009. [Google Scholar] [CrossRef]
- Tusting, L.S.; Ippolito, M.M.; Willey, B.A.; Kleinschmidt, I.; Dorsey, G.; Gosling, R.D.; Lindsay, S.W. The evidence for improving housing to reduce malaria: A systematic review and meta-analysis. Malar. J. 2015, 14, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanzirah, H.; Tusting, L.S.; Arinaitwe, E.; Katureebe, A.; Maxwell, K.; Rek, J.; Bottomley, C.; Staedke, S.G.; Kamya, M.; Dorsey, G.; et al. Mind the gap: House structure and the risk of malaria in Uganda. PLoS ONE 2015, 10, e0117396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tusting, L.S.; Thwing, J.; Sinclair, D.; Fillinger, U.; Gimnig, J.; Bonner, K.E.; Bottomley, C.; Lindsay, S.W. Mosquito larval source management for controlling malaria. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad-Franch, F.; Zamora-Perea, E.; Luz, S.L. Mosquito-disseminated insecticide for citywide vector control and its potential to block arbovirus epidemics: Entomological observations and modeling results from Amazonian Brazil. PLoS Med. 2017, 14, e1002213. [Google Scholar] [CrossRef] [PubMed]
- Boyce, R.; Lenhart, A.; Kroeger, A.; Velayudhan, R.; Roberts, B.; Horstick, O. Bacillus thuringiensis israelensis (Bti) for the control of dengue vectors: Systematic literature review. Trop. Med. Int. Health 2013, 18, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Geissbuhler, Y.; Kannady, K.; Chaki, P.P.; Emidi, B.; Govella, N.J.; Mayagaya, V.; Kiama, M.; Mtasiwa, D.; Mshinda, H.; Lindsay, S.W.; et al. Microbial larvicide application by a large-scale, community-based program reduces malaria infection prevalence in urban Dar es Salaam, Tanzania. PLoS ONE 2009, 4, e5107. [Google Scholar] [CrossRef] [PubMed]
- Mbare, O.; Lindsay, S.W.; Fillinger, U. Dose-response tests and semi-field evaluation of lethal and sub-lethal effects of slow release pyriproxyfen granules (Sumilarv® 0.5G) for the control of the malaria vectors Anopheles gambiae sensu lato. Malar. J. 2013, 12, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangoro, O.; Turner, E.; Simfukwe, E.; Miller, J.E.; Moore, S.J. A cluster-randomized controlled trial to assess the effectiveness of using 15% DEET topical repellent with long-lasting insecticidal nets (LLINs) compared to a placebo lotion on malaria transmission. Malar. J. 2014, 13, 324. [Google Scholar] [CrossRef] [PubMed]
- Sluydts, V.; Durnez, L.; Heng, S.; Gryseels, C.; Canier, L.; Kim, S.; Van Roey, K.; Kerkhof, K.; Khim, N.; Mao, S.; et al. Efficacy of topical mosquito repellent (picaridin) plus long-lasting insecticidal nets versus long-lasting insecticidal nets alone for control of malaria: A cluster randomised controlled trial. Lancet Infect. Dis. 2016, 16, 1169–1177. [Google Scholar] [CrossRef]
- Wilson, A.L.; Chen-Hussey, V.; Logan, J.G.; Lindsay, S.W. Are topical insect repellents effective against malaria in endemic populations? A systematic review and meta-analysis. Malar. J. 2014, 13, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horstmann, S.; Sonneck, R. Contact bioassays with phenoxybenzyl and tetrafluorobenzyl pyrethroids against target-site and metabolic resistant mosquitoes. PLoS ONE 2016, 11, e0149738. [Google Scholar] [CrossRef] [PubMed]
- Ogoma, S.B.; Mmando, A.S.; Swai, J.K.; Horstmann, S.; Malone, D.; Killeen, G.F. A low technology emanator treated with the volatile pyrethroid transfluthrin confers long term protection against outdoor biting vectors of lymphatic filariasis, arboviruses and malaria. PLoS Negl. Trop. Dis. 2017, 11, e0005455. [Google Scholar] [CrossRef] [PubMed]
- Fiorenzano, J.M.; Koehler, P.G.; Xue, R.D. Attractive toxic sugar bait (ATSB) for control of mosquitoes and its impact on non-target organisms: A review. Int. J. Environ. Res. Public Health 2017, 14, 398. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Vector Control Response 2017–2030; World Health Organization: Geneva, Switzerland, 2017; p. 53. [Google Scholar]
- Golding, N.; Wilson, A.L.; Moyes, C.L.; Cano, J.; Pigott, D.M.; Velayudhan, R.; Brooker, S.J.; Smith, D.L.; Hay, S.I.; Lindsay, S.W. Integrating vector control across diseases. BMC Med. 2015, 13, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouchet, J.; Pichon, G.; Gayral, P.; Hamon, J. Sensitivity of and resistance to insecticides of Aedes aegypti in West Africa and methods of control of the vector. Bull. World Health Organ. 1971, 45, 394–404. [Google Scholar] [PubMed]
- World Health Organization. Test Procedures for Insecticide Resistance Monitoring in Malaria Vectors, Bio-Efficacy and Persistence of Insecticides on Treated Surfaces; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Mazzarri, M.B.; Georghiou, G.P. Characterization of resistance to organophosphate, carbamate, and pyrethroid insecticides in field populations of Aedes aegypti from Venezuela. J. Am. Mosq. Control Assoc. 1995, 11, 315–322. [Google Scholar] [PubMed]
- Seixas, G.; Grigoraki, L.; Weetman, D.; Vicente, J.L.; Silva, A.C.; Pinto, J.; Vontas, J.; Sousa, C.A. Insecticide resistance is mediated by multiple mechanisms in recently introduced Aedes aegypti from Madeira Island (Portugal). PLoS Negl. Trop. Dis. 2017, 11, e0005799. [Google Scholar] [CrossRef] [PubMed]
- Rocha, H.D.; Paiva, M.H.; Silva, N.M.; de Araujo, A.P.; Camacho Ddos, R.; Moura, A.J.; Gomez, L.F.; Ayres, C.F.; Santos, M.A. Susceptibility profile of Aedes aegypti from Santiago Island, Cabo Verde, to insecticides. Acta Trop. 2015, 152, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Osei, J.H.; Sasaki, A.; Adimazoya, M.; Appawu, M.; Boakye, D.; Ohta, N.; Dadzie, S. Risk of transmission of viral haemorrhagic fevers and the insecticide susceptibility status of Aedes aegypti (Linnaeus) in some sites in Accra, Ghana. Ghana Med. J. 2016, 50, 136–141. [Google Scholar] [PubMed]
- Mathias, L.; Baraka, V.; Philbert, A.; Innocent, E.; Francis, F.; Nkwengulila, G.; Kweka, E.J. Habitat productivity and pyrethroid susceptibility status of Aedes aegypti mosquitoes in Dar es Salaam, Tanzania. Infect. Dis. Poverty 2017, 6, 102. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Nomura, Y.; Satar, G.; Hu, Z.; Nauen, R.; He, S.Y.; Zhorov, B.S.; Dong, K. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc. Natl. Acad. Sci. USA 2013, 110, 11785–11790. [Google Scholar] [CrossRef] [PubMed]
- Hirata, K.; Komagata, O.; Itokawa, K.; Yamamoto, A.; Tomita, T.; Kasai, S. A single crossing-over event in voltage-sensitive Na+ channel genes may cause critical failure of dengue mosquito control by insecticides. PLoS Negl. Trop. Dis. 2014, 8, e3085. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, B.J.; Pignatelli, P.; Nikou, D.; Paine, M.J. Pinpointing p450s associated with pyrethroid metabolism in the dengue vector, Aedes aegypti: Developing new tools to combat insecticide resistance. PLoS Negl. Trop. Dis. 2012, 6, e1595. [Google Scholar] [CrossRef] [PubMed]
- Kasai, S.; Komagata, O.; Itokawa, K.; Shono, T.; Ng, L.C.; Kobayashi, M.; Tomita, T. Mechanisms of pyrethroid resistance in the dengue mosquito vector, Aedes aegypti: Target site insensitivity, penetration, and metabolism. PLoS Negl. Trop. Dis. 2014, 8, e2948. [Google Scholar] [CrossRef] [PubMed]
- Grigoraki, L.; Pipini, D.; Labbe, P.; Chaskopoulou, A.; Weill, M.; Vontas, J. Carboxylesterase gene amplifications associated with insecticide resistance in Aedes albopictus: Geographical distribution and evolutionary origin. PLoS Negl. Trop. Dis. 2017, 11, e0005533. [Google Scholar] [CrossRef] [PubMed]
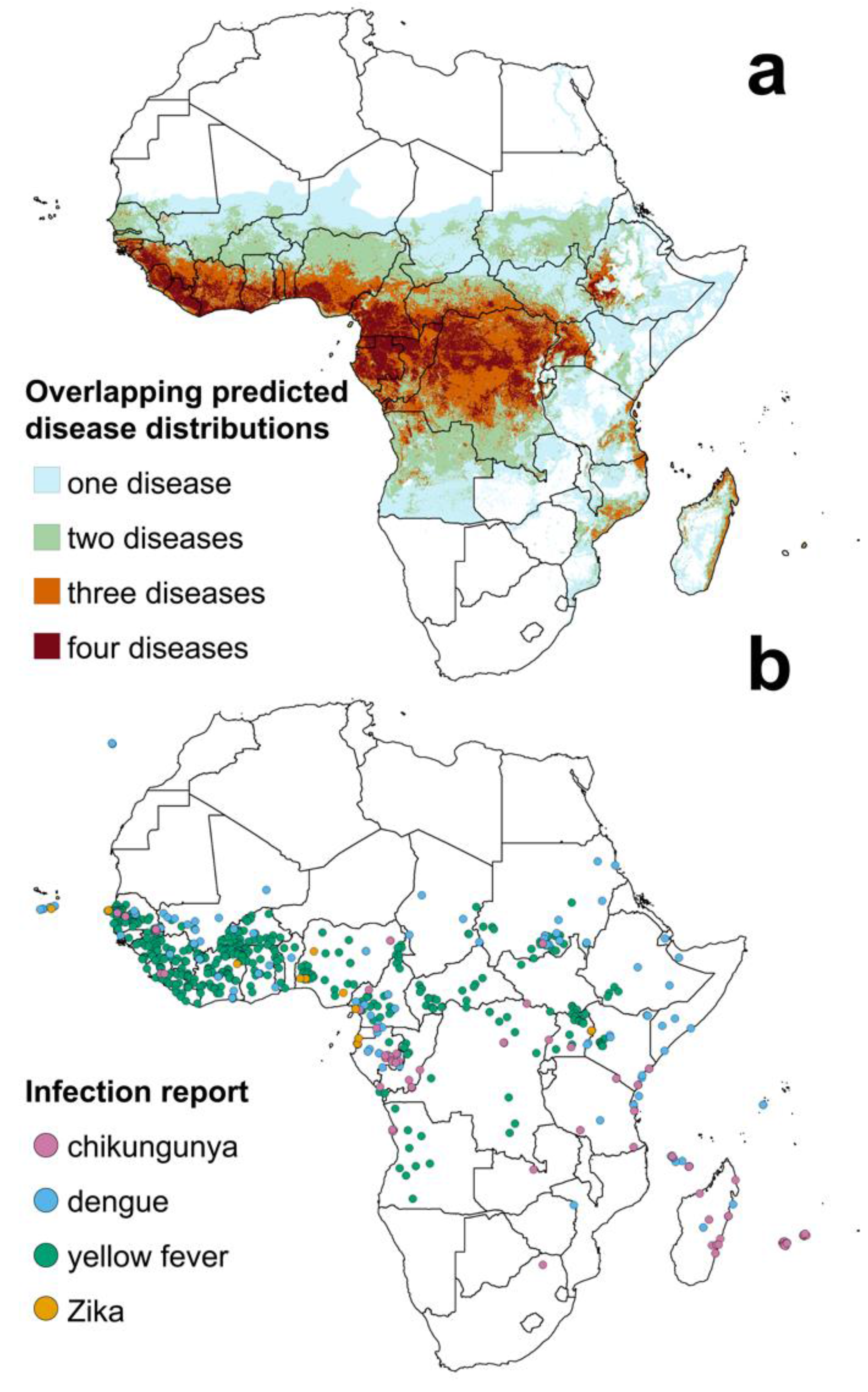

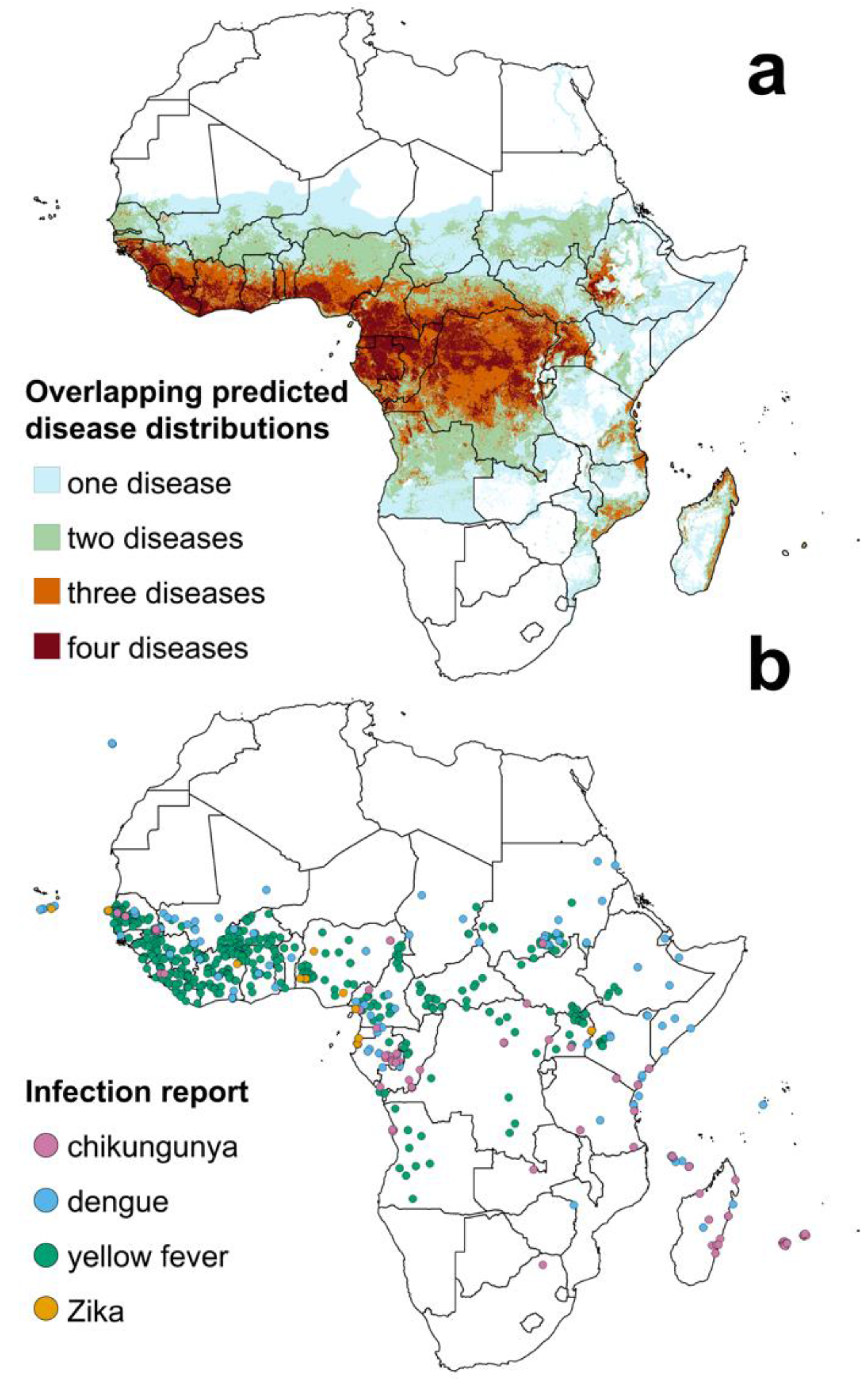{kind=link}
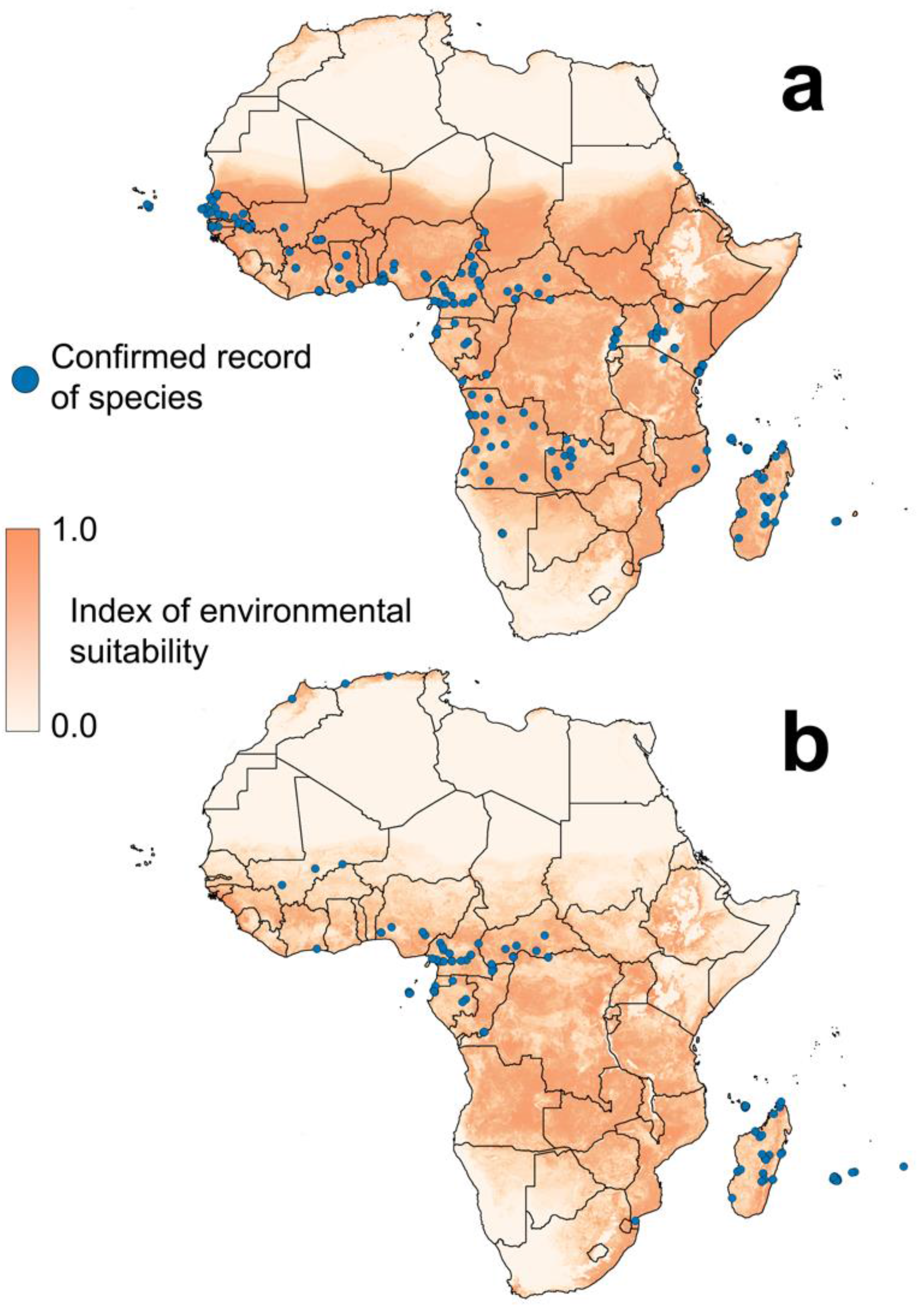
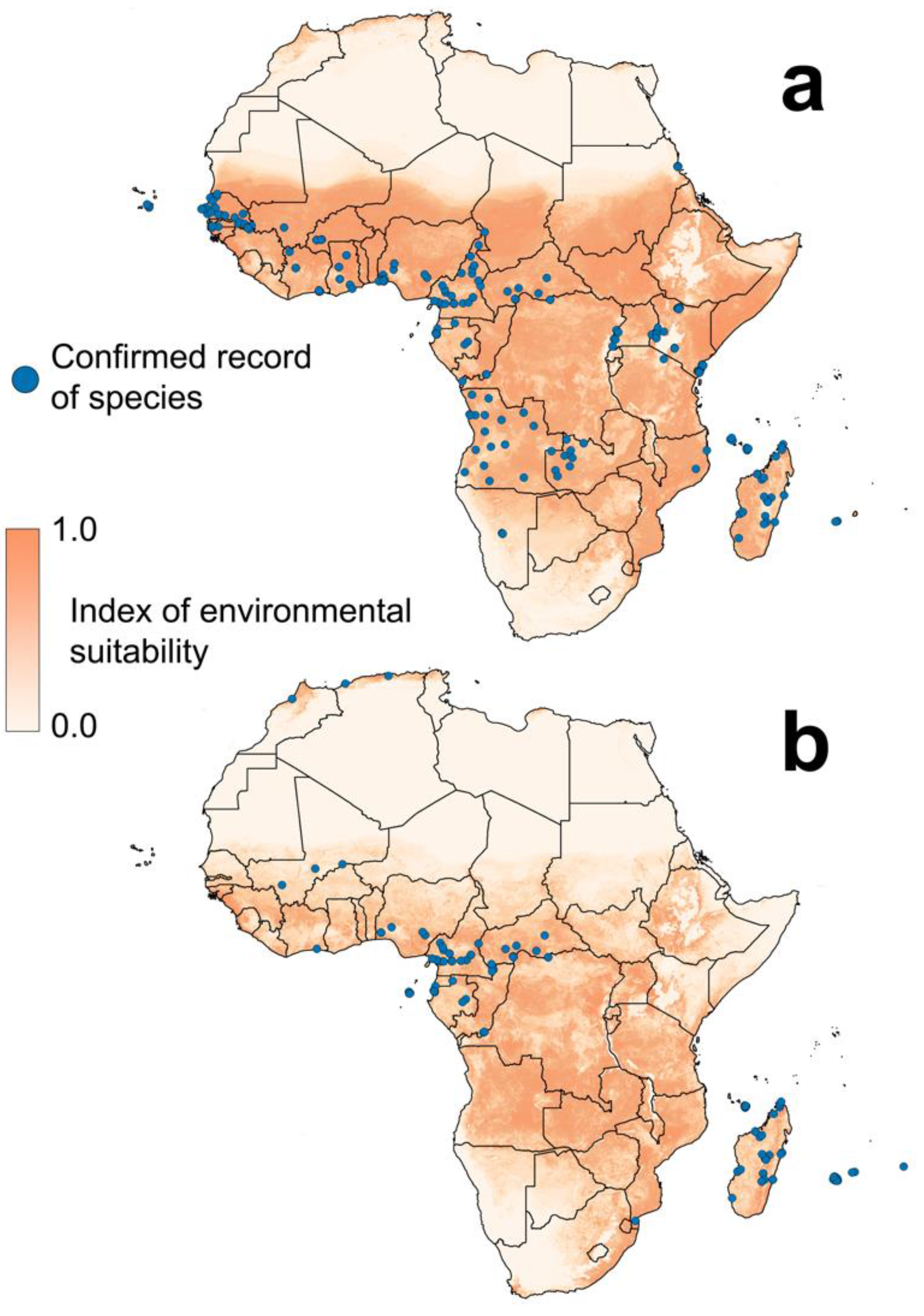{kind=link}
| Infection | Estimated Population at Risk | Percentage of African Population |
|---|---|---|
| Chikungunya | 271 million | 23% |
| Dengue | 750 million | 63% |
| Yellow fever * | 21 million | 2% |
| Zika | 406 million | 34% |
| At least one of the above | 831 million | 70% |
| Study | Year | Country | Area | Species | DDT (a) | Pyr I (a) | Pyr II (a) | Carb (a) | OP (a) | Temephos (l) | Bti (l) | Other (l) | kdr Mutations | Metabolic Resistance |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [162] | 2013 | Madeira | Funchal, Paul do Mar | Ae. aegypti | R | R | R | R, S | F1534C; V1016I | MFO, esterases (biochemistry); PBO, DEM significant (synergist; metabolizing genes overexpressed) | ||||
| [43] | 2009 | Cape Verde | Santiago | Ae. aegypti | R | S | S | R | S | |||||
| [163] | 2012 | Cape Verde | Santiago, Praia | Ae. aegypti | R | S | S | Not detected | MFO, esterases, GSTs (biochemistry) | |||||
| [63] | 2009 | Senegal | Dakar | Ae. aegypti | R | S | R, RS | R | S | |||||
| [52] | 2010 | Côte d’Ivoire | Abidjan | Ae. aegypti | S | RS, S | R, RS | |||||||
| [46] | 2014 | Côte d’Ivoire | Abidjan | Ae. aegypti | R | S | RS | RS | S (DDT, Pyr) | |||||
| [164] | 2012–2013 | Ghana | Accra | Ae. aegypti | R | R | R | |||||||
| [51] | 2013–2014 | Ghana | Widespread | Ae. aegypti | R | R, RS, S | F1534C; V1016I | |||||||
| [51] | 2013–2014 | Ghana | Widespread | Aedes aedes formosus | R | R, RS, S | F1534C | |||||||
| [37] | 2011–2012 | Nigeria | Lagos | Ae. aegypti | R | S | RS, S | |||||||
| [60] | 2013 | Nigeria | Kwara State | Ae. aegypti | S | S | R | |||||||
| [49] | 2007 | Cameroon | Widespread | Ae. aegypti | RS | S | S | S | S | S | ||||
| [71] | 2015–2016 | Cameroon | Yaoundé | Ae. aegypti | R | S | R | R, S | S | Not detected | Limited effect of synergist PBO | |||
| [58] | 2013 | Central African Rep. | Bangui | Ae. aegypti | R, RS | RS, S | S | S | S | S | Not detected | MFO, esterases, GSTs (biochemistry) | ||
| [49] | 2007 | Gabon | Libreville | Ae. aegypti | R | S | S | S | S | S | ||||
| [48] | 2009, 2010 | Sudan | Port Sudan | Ae. aegypti | R | RS, S | S | R, RS | ||||||
| [165] | 2015 | Tanzania | Dar es Salaam | Ae. aegypti | R, RS | R, RS | ||||||||
| [166] | 2010–2011 | Mayotte | Petit Terre | Ae. aegypti | S | S | S | S (multiple) | ||||||
| [49] | 2007 | Cameroon | Widespread | Ae. albopictus | R | RS, S | S | S | S | S | ||||
| [71] | 2015–2016 | Cameroon | Yaounde | Ae. albopictus | R | R, S | R, RS | R, RS | S | Not detected | Variable effect of synergist PBO among locales | |||
| [58] | 2014 | CAR | Bangui | Ae. albopictus | R, RS, S | R, RS, S | RS, S | RS, S | S | S | Not detected | MFO, esterases, GSTs (biochemistry) | ||
| [49] | 2007 | Gabon | Libreville | Ae. albopictus | S | |||||||||
| [61] | 2010–2011 | Mayotte | Kaweni | Ae. albopictus | S | S | S | S (multiple) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weetman, D.; Kamgang, B.; Badolo, A.; Moyes, C.L.; Shearer, F.M.; Coulibaly, M.; Pinto, J.; Lambrechts, L.; McCall, P.J. Aedes Mosquitoes and Aedes-Borne Arboviruses in Africa: Current and Future Threats. Int. J. Environ. Res. Public Health 2018, 15, 220. https://doi.org/10.3390/ijerph15020220
Weetman D, Kamgang B, Badolo A, Moyes CL, Shearer FM, Coulibaly M, Pinto J, Lambrechts L, McCall PJ. Aedes Mosquitoes and Aedes-Borne Arboviruses in Africa: Current and Future Threats. International Journal of Environmental Research and Public Health. 2018; 15(2):220. https://doi.org/10.3390/ijerph15020220
Chicago/Turabian StyleWeetman, David, Basile Kamgang, Athanase Badolo, Catherine L. Moyes, Freya M. Shearer, Mamadou Coulibaly, João Pinto, Louis Lambrechts, and Philip J. McCall. 2018. "Aedes Mosquitoes and Aedes-Borne Arboviruses in Africa: Current and Future Threats" International Journal of Environmental Research and Public Health 15, no. 2: 220. https://doi.org/10.3390/ijerph15020220
APA StyleWeetman, D., Kamgang, B., Badolo, A., Moyes, C. L., Shearer, F. M., Coulibaly, M., Pinto, J., Lambrechts, L., & McCall, P. J. (2018). Aedes Mosquitoes and Aedes-Borne Arboviruses in Africa: Current and Future Threats. International Journal of Environmental Research and Public Health, 15(2), 220. https://doi.org/10.3390/ijerph15020220